COPYRIGHTC 1977 American Society forMicrobiology Printed inU.S.A.
Morphogenesis
of Vesicular Stomatitis Virus:
Electron
Microscope Observations with Freeze-Fracture Techniques
DENNIS T. BROWN* AND BIRGIT RIEDELInstitutfarGenetik der Universitdt zuK6ln, 5 Cologne 41, West Germany
Received for publication21July1976
The morphogenesis of vesicular stomatitis virus was examined using freeze-fracture techniques, and the results obtained were compared with those from
previouslypublished experimentscarriedoutwith influenza virusesand
togavi-ruses. The process of conversion of the host cell plasma membrane into the
vesicularstomatitis virusenvelope wasaccompaniedby a lossof the
intramem-branal particles abundantincell membranes.Frequentlyadense accumulation
ofintramembranal particles could beseen atthe baseof the developing virion,
suggesting that thesestructuresmight playsomeroleinthe generation of viral
envelope. In addition to the viral structures that were seen to
develop
in theclassical fashion, with their long axisperpendicular to the cellsurface,
struc-tures were also found that suggested the initiation of a process similar to
budding, with the long axis of the viralcapsid paralleltothe
plasma
membrane.In this situation, as in the "perpendicular" process, intramembranal particles
wereexcluded from theviral structure, andanaccumulation of theseparticles
could beseenadjacentto the developingviral membrane.
Vesicularstomatitis virus (VSV) is an enve- 20). Little more is known regarding the
proc-lopedRNA-containing virus which acquires its esses that organize the protein components of
membranous envelope by the process of "bud- the virion during the budding process. Models
ding" through the plasma membrane of the have beensuggested (basedonelectron
micro-host cell after that membrane has been modi- scope observations of thin sections of
VSV-in-fiedby the insertion of
virus-specified
glycopro- fected cells) that show the viralcomponents totein (5, 8, 26). VSV is a structurally complex beassembledinastepwisefashionasbudding virion which has theshape of either a bullet, progresses. Thematuration process is
morpho-withoneendtapered and the othersquare, or a logically depicted as beginning atthe tapered
bacillus, with both ends tapered, when viewed end of the virion and progressing in such a
in the electron microscope (4, 14, 19, 25). The manner that the virion buds with its long axis
coreof thevirion ishelicalinstructure,has the perpendicular to the surface of the plasma
overall shape ofabullet, and islocated inthe membrane (29). This model allows the core to
center of thecylindrial virion (4, 25).The viral beprogressivelyshapedinto atight spiralasits
membrane is believed to be comprised oftwo protein interactswith themodifiedmembrane.
virus-specified proteins, aninternally situated The Gprotein isexposedorreorganizedintothe
membrane protein (M) and an externally lo- spikeontheexteriorof thedevelopingvirion as
catedglycoprotein (G) (4, 25).Theorganization the process continues (25, 26). No suggestion
ofthese proteinsinthe membraneis notclear. has been offeredas tohowthecompletedvirion
The G proteinisclearlyexposedonthe surface isreleasedfrom the cell surface.
of the virion and serves as the organelle of We have investigated the process of VSV
attachment to susceptible host cells (1, 12, 18, maturation using the
procedure
offreeze-frac-19, 25). TheMprotein isbelievedtobe located turing. This procedure hasanumberof advan-on the inner surface of the viral membrane; tages over classical thin sectioning forstudying however, it is unclear to what extent (ifany) the morphogenesis of membrane-bound
vi-this protein extends into the envelope bilayer ruses. Thefreeze-fracturingprocess eliminates itself (25). It is also not clear whether the G artifacts of dehydration and preserves three-protein extends across the membrane bilayer to dimensional structure. The fracturingprocess interact with the M protein. It has been shown exposes to view the interior of the developing
recentlythat the G protein does have ahydro- viral membrane, makingitpossibletoobserve
phobic fragment of about 5,000 molecular topological alterations occurring within the weight buried in the envelope membrane (13, membrane during envelopment (17, 24). The
601
on November 10, 2019 by guest
http://jvi.asm.org/
verted from host plasma membrane to viral (abbreviated E) refers to the layer that is in contact
envelope. Thischangewasreflected in the loss with the extracellular fluid. Each of these bilayer
of the intramembranal particles from the leaflets has an inwardly facing (toward the
cyto-plasmamembraneasit became virusenvelope. plasm) andan outwardly facing(towardthe
extra-The intramembranal particles have been cellular fluid) surface. Of these four possible
sur-shown to be membrane structural proteins faces, only twoarerevealed by the freeze-fracturing whichextendinto or acrossthenonpolar region process used in this study: the outwardly facing of the membrane bilayer
(22,
24).Thus,
the surface of the protoplasmic part of the membranetopologicalchange
andbilayer
(protoplasmicfractureface, PF) and the in-topologicalchange
ane
the
structure-free ap- wardly facingsurface of theexoplasmic part ofthe pearance of the interior of the envelopes of in- bilayer (exoplasmic fracture face, EF). Both of thesefluenzavirus andthe togavirus Sindbiscanbe interior membrane surfaces possess the 6- to 10-nm
interpretedto support the hypothesis that the intramembranal particles believed to represent
envelopestructuralproteinsof these virionsdo membrane proteinswhich penetrateinto oracross
notextendinto or across thenonpolar regionof the fracture plane, the nonpolar region of the
bi-the envelope bilayer (3, 7, 10, 21). A freeze- layer (17, 22-24). Theseparticles can beusedasan
fracture study of VSV structure and develop- aid in identifying the exposedfractured faces, be-mentwas therefore undertaken to determine if
cause
the protoplasmicfracture face (PF)possesses thismodelrhabdovirus would besimilarto t the structures ingreater abundance than theexo-this
mouel rabdovilrus
would besimilartothe plasmicfractureface (EF) (3, 17, 23, 24).previously studied influenza and togavirus sys- Viral envelopes fracture in a fashion similar to
tems. that described for plasmamembranes. The
fractur-ingprocess reveals an external fracture face (EF) MATERIALS AND METHODS similar to the exoplasmic fracture face of theplasma
membrane (EF) and a fracture face of the envelope Cells, virus, andmedia. VSV(Indianastereotype) bilayer part adjacent to the core. Because the viral was agift of Elmer Pfefferkorn(DartmouthMedical interiorcontains no protoplasm, we feel the symbol
School, Hanover, N.H.). The viruswasgrownafter
Pis
intap ia foprodescribin
thi
eandbol
plaque purification in BHK-21 cells, which were haveadoptedthedesignation"C"rfor
thatpartofathe
received from Peter Faulkner (Queens University, envelope bilayer locatedclosest tothe core or capsid. Kingston, Ont., Canada). Cell and virus growth Thus, "CF"represents the fractureface in theviralwerecarried out at37°CinEagle minimal essential envelopethat isanalogous to the PF surfaceexposed
medium (6) containing 10% tryptose phosphate byfracturingtheplasmamembrane. brothand10%fetal calf serum. Virus titrationwas
carried out witha 1%agaroseoverlay containing the RESULTS mediumdescribed above.
Preparation of specimens for electron micros- Ultrathinsectioning of VSV-infected cells
re-copy.Monolayersof cells were infected atabout 95% vealed the virions developingfrom the plasma
confluency with 20 PFU of VSV percell from the membrane as described previously (25, 29), and
firstpassageafterplaquepurification. The infected a single electron micrograph is provided here as cells were incubated for 10 h at 37°C and subse- a frame of reference for the reader in
examin-quently washed with phosphate-buffered saline
ing
thefollowing images
offreeze-fracturerep-(4WC)
and fixed inmonolayers with 2%glutaralde-lia
the1o
fhydeincoldphosphate-buffered saline for 2 h. The licas
(Fig.
1).fixed cells were scraped from the monolayers and Figure2 shows an area of aVSV-infected cell embedded for ultrathin sectioningasdescribedpre- surface after freeze-fracturing. The fracture viously (11) or prepared for freeze-fracturing. plane has passed through theexoplasmic part
Freeze-fracturing was carried out exactly as de- of the membrane bilayer and has exposed to
scribed by Brown et al. (3), exceptthatbothfixed view the PF of the membrane bilayer before and unfixed infected cells were used. The results breaking through the protoplasmic part
expos-obtainedwerethesameregardless of whether fixa-
ing
the
cell cytoplasm.
On theexposed
surface tion was used.Freeze-fracturing wascarried out in a canbe
seen
large
numbers of the intramem-Balzers BAF-300freeze-etching device,andreplicascan
parge
numbed
of
weasem
werephotographedinaSiemens 101 electron micro- branal particles (described above) as well as a
scope. number of partially enveloped virions. At the
The triple-layered unit membrane seen in thin "ice level," where the exoplasmic part of the
sectionsissplit into two components by the cleaving bilayerhas beencutbythefracturing process,
process (17, 23, 24).Electron micrographsoffreeze- bothparts of the membranebilayer follow
on November 10, 2019 by guest
http://jvi.asm.org/
*,*. .*
4 .$ -d ~~~~~~~~~ " A ~wfK K
4 ~~~~~~~~-t
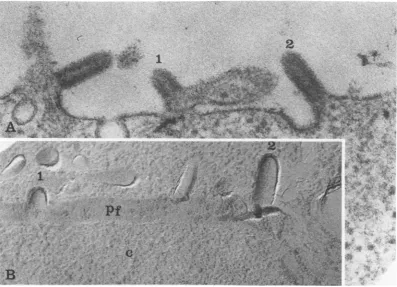
FIG. 1. Budding of VSV as seen by ultrathin sectioning andfreeze-fracturing.(A) Developing virions thin sectioned along their longaxis: (1) a virion at an early stage in budding; (2) a more completely developed
particle. x105,000. (B) Developing virions similar to those shown in (A) but observed in a replica of a freeze-fractured cell; (1) and (2) as described for (A). PF, Protoplasmic fracture face. x105,000.
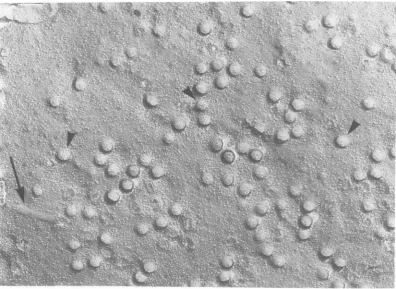
out interruption the contour of the developing have no intramembranal structure. The devel-virion. Themembranethatisformingtheviral oping viral envelopes viewed from above in envelope is, therefore, continuous with the both Fig. 3 and 4 are generally free of the plasma membrane from which it is derived. intramembranal particles, as were the develop-The portion of the membrane clearly incorpo- ing envelopes viewed from the side shown in rated into the viral bud is free of the intramem- Fig. 2. In a survey of 200 budding structures of branal particles that can be readily detected in the type shown in Fig. 3 and 4, only seven
the membrane at the base of the developing virions were found to have particulate
struc-viral envelope. In some instances there is an tures associated with their surface that were unusually large accumulation of the particles morphologically identical to the host intra-at the base of the developing virion, where the membranal particles. Most of the structure transition from host membrane to viral enve- seen in the developing viral envelopes could be lope takes place. In some instances a halo of attributed to fine structure of the platinum-material could be seen at the outer surface of carbon replica itself. In these images, as in Fig. the developing viral structures (Fig. 1, 2). This 2, an accumulation of particles at the base of halo of material may represent the appearance the developing virion can frequently be seen. of the viral spikes. Occasionally the normal distribution of intra-In Fig. 3 and 4 the PF of the infected cell membranal particles, in areas where budding
membrane is widely exposed, the axis being structures were not apparent, was disturbed in
perpendicular to the plane of section. Large such a way as to produce a bead-free area
hay-numbers of budding virions can be seen in both ing the size and shape of a VSV particle (Fig. 4) figures. Figure 3 has a number of nearly com- viewed on its long axis. Figure 5 shows an area pleted or possibly complete virions at the "ice of the PF of the membrane bilayer which has an level." These virions, like the budding forms, extraordinary number of these structures. The
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.49.447.68.355.2]_ '
'ir.
ci''
vre,2, n ow
V *~4S
-
Ni~~~ei
).415 ~. "' J.
4S1
j,W.~ ~ I
7'
N~~~~~~~~~9
I
4.
packed ring of theintramembranal particles at its base. A virion that was fractured in such a way as to reveal its external fracture face (ef) is also shown. x103,400.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 21, 1977 VSV DEVELOPMENT
605
it~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~,~
£"
-t~~~~~OC
JA.l- a
FIG. 3. Area of the protoplasmic fracture face of a VSV-infected cell. The view is from a position
perpendicular to the cell surface. A number of budding structures are viewed on their short axis. The intramembranalparticlesareabsentfrom the developingbuds, although theyareabundantinotherareasof thissurface. At theedge of the exposed cell surface (where the outer leaflet of the bilayeriscross-fracturedand thesurrounding mediumisexposed) are seen a number ofcompletedvirionswhich haveremainedonthe cell surface (arrows). A number ofbudding virions have beencutfrom the cellsurfacebythefracturingprocess (a).Occasionally this cross-fracture leaves the broken membrane witha moreorderlyappearance(arrow and insert). x57,000;insert, x168,000.
structureshave the sizeand
shape
of VSV viri- appearedto be surroundedby
the beads(Fig.
ons and, like the partially
developed
virions 5C).showninFig. 2through4,lacktheintramem- Because the freeze-fracture technique is so
branal particles that can readily be demon- ideally suitedtostudy of thecontours of
mem-stratedadjacenttothevirusforms. Thesestruc- branes, it was hoped that this study might
tures occurred rarely compared with the bud- provide information on how the virion is
re-dingstructuresshowninFig. 2through4 (1 in leased from the cell surfaceafterattaining its
80 to 100budding forms) butdosuggest that a characteristicshape. Althoughalarge number
process ofbudding can atleast be initiated in ofbuddingformswereexaminedinthis study, which the viral core structure interacts with noindication of an infolding of the membrane the modified cell plasma membrane in such a atthe baseof the virion wasdetected. Theonly way as to deform it into a shape with the di- structures seen that might be related to some mensionsof a virion viewed perpendicular to its kind of release mechanism were of the type long axis. Accumulations of the intramem- shown in Fig. 3. The ring-shaped structure
branalparticles canoccasionallybedetectedat shownin thisfigure does not have the rough-the square end ofthoseparticles havingabullet edged appearance of a virion that has been
shape (Fig. 5B), whereasother suchstructures broken from the cell surface bythe fracturing
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.51.446.69.388.2]'-7.'~~~~~~~~~~~~~~~~~~~~~~~~Z
(~~~~~~~
..L
tzo~~ ~ ~
[image:6.501.61.458.76.365.2]FG4.Protoplasmicfracture face ofa VSV-infected cell. A large number of budding virions are seen, as In Fig. 3. The arrowheads point out a few viral structures that appear to have intramembranal particles. The arrow points out an area of the inner fracture face where the interior membrane beads are not present. This area has dimensions similar to those of a VSV virion viewed on its longaxcis. x63,OOO.
process. Such structures may represent the of development. Like influenza, but unlike process of structural reorganization occurring Sindbis virus, VSV budding structures were at the base of a virion as it is released from the found to be generally free of intramembranal host cell plasma membrane. particles as soon as the buds could be morpho-Membrane fracture faces complementary to logically distinguished (1). Sindbis virus was those presented here (exoplasmic fracture found to possess intramembranal particles in faces) were also found and were examined in its envelope early in development, but these this study (Fig. 6). The exoplasmic fracture face structures were completely lost before the viral of the developing and mature viral envelopes core was half enveloped (3). VSV differed from were found to be free of intramembranal parti- both Sindbis and influenza viruses in that an clesor other detectable structure. It should be increase in the density of the intramembranal pointed out, however, that because the analo- particles could be frequently found at the base gous surface of the host plasma membrane has of the developing VSV envelope. This area is itself very few associated intramembranal par- the region where the transition from host
mem-tidles, the morphological change described brane to virus envelope takes place and may
above could not be detected. imply some special topological event
accompa-DISCUSSION
~~nying
the generation of the viral membrane.DISCUSSION
~~The
viral N and M proteins must have some This freeze-fracture study of VSV-infected mechanism for identifying the presence of G cells revealed similarities and differences in the protein on the outer surface of the cell plasma development of this rhabdovirion compared membrane in order for formation of the enve-with previously studied influenza and togavi- lope to take place and tobe able to incorporate ruses. VSV, like both influenza and Sindbis the glycoprotein into the mature virion in a viruses, was found to be free of intramem- relatively uniform ratio to the internally lo-branal particles when in a nearly mature state cated N and M proteins. The absence ofon November 10, 2019 by guest
http://jvi.asm.org/
- .y
1"--I-A-4~~~~~~~~~~~~~~~~~~~~~~~~~~~~~-(
4
~
~
~
-A
V~~~~~~&
Ad~~~~~~~~~~~~~~~~~
~~~~~~
e~~~~~~~
- - A~~~~~~~~~~ -NI4
A%~~ ~ ~ ~ ~ ~ p
FI.5.Poopamc rctr ac fa SV-nete el aig arenmbro srctrsthtapert
beviinsdeeopn wthtei ln ai prall oth cl mmbae.Te reswhrete iusstutue
607
on November 10, 2019 by guest
http://jvi.asm.org/
B~~~~~~~~~~~~~~~~
FIG 6 Exoplasmic fracture faces ofVSV-infected BHK cells. (A) Virions maturing with long axis
perpendicular to cellsurface (fracture face complementaryto thosepresentedin Fig 1) x100000. (B) A
depressionintheexoplasmic fracture face (bordered byarrowheads)possiblycreatedbyavirionbuddingas
showninFig.5.AsinFig.5, the intramembranalparticles(seenonlyinB)areabsentfromthatpartofthe membrane interiorassociated with the virion. x1OO,000.
membranal
particles
inthecleavedenvelope
of ical structure. (ii) Thenucleocapsid
is also ca-the mature andpartially
mature virions sup- pable oforganizing
itself into the form that it ports the notion that the structuralproteins
of will ultimately assume in the virion without theviralenvelope
do notpenetrate
intoor crossbeing
completely
in contact with the modified thenonpolar region of theenvelopebilayer (10). membrane (the otherside of such a nucleocap-It ispossible, however, that at some very early sid would be exposed to the cell cytoplasm), aspointintheir beingincorporated intothe viral is the case in the former situation. It is not
envelope, the VSV membrane proteinsinteract possibletodetermineby electronmicroscopy if
with one anotheracrossthe nonpolarregionof such laterally arranged core structures ever
the membrane bilayer. As the proteins become bud to a completed virion. The morphological
a fixed part of the envelope, this transmem- changes occurring within the membrane
bi-branal interaction may be lost, producing the layeraccompanying this lateral budding
proc-structurally smooth envelope interior and the essare,however,identicaltothoseoccurring in
appearanceof thevirusspikes onthe surface of the predominating "normal"process.
the developing envelope. In such a case one ACKNOWLEDGMENTS
might propose that the dense accumulation
mightramembranal
parat
thedethe
base of theofACNWEGThis
research wassupported by
NTtheDeutsche
For-intramembranal particles at the base of the schungsgemeinschaftthrough SFB74.
developing envelope "C" fracture face
repre-sentssuchacomplex ofvirusenvelope proteins LITERATURE CITED
(G and M), which have been accumulated by 1. Bachi,T., W. Gerhard, J. Lindenmann, and K.
Muhle-the presence of the nucleocapsid just prior to thaler. 1969. Morphogenesisof influenza A virusin suchastructural transition. Ehrlichasites tumor cellsasrevealed bythin
section-Thelongitudinal association of the VSVcore ing andfreeze-etching.J.Virol.4:739-776.
2. Branton,D.,S.Bullivant, M.J.Karnovsky,H.Moor, withtheplasma membrane (Fig.5)issimilarto K.
Muhlethaler,
D. H. Northcote, L. Packer, B. Sa-that ofstructuresobservedin thinsections by tir, P. Satir, V. Speth, L. A. Staehlin, R. L.Steere, Orensteinetal. (16) andsuggestthat the form- and R.S. Weinstein.1975.Freeze-etchingnomencla-determining
deterinincharacteristicschaacterstic of theof he nuleocpsidnucleocapsid
3. Brown,ture.Science 190:54-56.D. T., M. R. F. Waite, and E. Pfefferkorn.canfunctionin morethanone way.(i)Oneway 1972. Morphology and morphogenesis of Sindbis virus isthe classical development of thevirus struc- as seen with freeze-etching techniques. J. Virol. ture along its long axis (which is by far the 10:524-536.
predominatingprocess seen), with the nucleo- 4. Cartwright, B.,S. J. Smale, F. Brown,and R. Hull.
predominating process 1972. Modelforvesicular stomatitis virus. J. Virol.
capsid being
progressivelyorganized into a hel- 10:256-260.on November 10, 2019 by guest
http://jvi.asm.org/
5. David,A.E. 1973. Assembly of the vesicular stomatitis proteinof vesicular stomatitis virus: role of the neu-virus envelope: incorporation of viral polypeptides raminic acid in infection. J.Virol. 14:270-281. intothehost plasma membrane. J. Mol. Biol. 76:135- 19. Schloemer, R. H., and R. R. Wagner. 1975. Cellular 148. adsorption function of the sialoglycoproteinof vesicu-6. Eagle, H. 1959.Amino acid metabolism in mammalian lar stomatitis virus and its neuramicacid. J. Virol.
cell cultures. Science 130:432-437. 15:882-893.
7. Harrison, S. C., A. David, J. Jumblatt, and J. E. Dar- 20. Schloemer, R. H., and R. R. Wagner. 1975. Association nell. 1971. Lipid and protein organization in Sindbis of vesicular stomatitis virus glycoprotein with mem-Virus. J. Mol. Biol. 60:523-528. branes: isolation and characterization of a lipophilic 8. Kang, C. Y., and L. Prevec. 1971. Proteins ofvesicular fragment ofthe glycoprotein. J. Virol. 16:237-249.
stomatitis virus. III.Intracellular synthesis and ex- 21. Sefton, B. M., and B. J. Gaffney. 1974. Effect of the tracellular appearance of virus specific proteins. Vi- viral proteins on the fluidity of themembrane lipids rology 46:678-690. in sindbisvirus. J. Mol. Biol. 90:343-358.
9. Kelly,J. M., R. R. Wagner, and S. U. Emerson. 1972. 22. Singer, S.J. 1974. The molecular organization of mem-The glyco-protein of vesicular stomatitis virus is the branes.Annu. Rev. Biochem. 43:805-833.
antigenthat gives rise toandreacts withneutraliz- 23. Tillack, T. W., and V. T. Marchesi. 1970. Demonstra-ingantibody. J. Virol. 10:1231-1235. tionof the outer surface offreeze-etched red blood cell 10. Lenard, J., and R. W. Compans. 1974. The membrane membranes. J. Cell. Biol. 45:694.
structureoflipidcontainingviruse$. Biochim. Bio- 24. Tillack, T. W., R. E. Scott, and V. E.Marchesi. 1972. phys. Acta 344:51-94. The structure of the erythrocyte membranesstudied 11. Luft,J. H. 1961. Improvements in epoxy resin embed- by freeze-etching. II. Localization of receptors for ding methods. J. Biophys. Biochem. Cytol. 9:409. phyto hemagglutininand influenza virus to the intra-12. Moore, N. F., J. M. Kelly, and R. R. Wagner. 1974. membranous particles. J. Exp.Med. 135:1209-1227.
Envelope proteins of vesicular stomatitis viruses: ac- 25. Wagner, R. R. 1975.Reproduction of rhabdoviruses. In cessibility toiodination. Virology 61:292-296. H.Fraenkel-ConratandR. R.Wagner, (ed.), Com-13. Mudd, J. A. 1974. Glycoprotein fragment associated prehensive virology, vol. 4. Plenum,New York.
with vesicular stomatitis virus after proteolytic 26. Wagner, R. R.,J. W. Heine, G. Goldstein,and C. A. digestion. Virology 62:573-577. Schnaitman. 1971. Use of antiviral-antiferritin hy-14. Nakai, T., and A. F.Howatson.1968. The fine structure brid antibody for localization of viral antigen in
ofvesicular stomatitis virus.Virology35:268-281. plasma membrane. J. Virol. 7:274-277.
15. Orenstein,J., L. Johnson, E. Shelton, and R. A. Lazza- 27. Wagner, R. R.,C. A. Schnaitman, and R. M.Snyder. rini. 1976. The shape of vesicular stomatitis virus. 1969. Structural proteins of vesicular stomatitis
vi-Virology71:291-301. rus.J. Virol. 3:395-403.
16. Orenstein, J., E. Shelton, and R. A. Lazzarini. 1975. 28. Walter, G.,and J. A. Mudd. 1973. Iodination of
vesicu-Associationof ribosomes withintracellularvesicular lar stomatitis virus withlactoperoxidase. Virology
stomatitis virusparticles.J. Virol. 16:447-452. 52:574-577.
17. Pinto de Silva, P., and D. Branton. 1970. Membrane 29. Zee, Y. C., A. J.Hackett,and L. Talens. 1970. Vesicu-splitting in freeze-etching. J. Cell. Biol. 45:590-605. lar stomatitis virus maturation sitesinsixdifferent 18. Schloemer, R. H., and R. R. Wagner. 1974. Sialoglyco- hostcells. J. Gen. Virol. 7:95-102.