0022-538X/82/030990-10$02.00/0
DNA
Sequence
of the
5' Terminus Containing
the
Replication
Origin
of
Parvovirus Replicative Form DNA
SOLON L. RHODE III* AND BELIAKLAASSENt
Institutefor Medical Research ofBennington, Bennington, Vermont05201 Received 6 August 1981/Accepted 13 October 1981
The nucleotide sequence of the 5' terminus of the parvovirus H-1 was determined. There are twoorientations of the 242-base-pair terminalpalindrome in nativereplicative form DNA, one inverted with respect to the other. Adjacent tothe terminalpalindrome is an AT-rich regionthat isnoncodingandcontainsa
55-base-pair tandem repeat. The addition mutant of H-1, DI-1, was also
se-quenced in this region and shown to have three copies of the tandem repeat sequence. Similarly, the related parvovirus H-3 contains only one copy of this repeat sequence. This region contains the replication origin for parvovirus replicative form DNA replication. Some of the implications of these results are
discussed.
Thenondefective parvoviruses are small ico-sahedral viruses that contain a linear single-stranded DNA of about 1.6 x 106 daltons (13,
34, 36). The virion DNA is predominantly, but
notentirely, minus strand and is the template for allvirus-inducedmRNA's that have been identi-fied (9, 30). During the replication process, the virion single-stranded DNA is converted to a linear duplex replicative form (RF) DNA (16, 35). This RF DNA undergoes amplification by a semiconservative modeofDNAreplicationthat requiresaviral RF rep geneproduct which has not been characterized (16, 22). The RF DNA serves as atemplate for viral RNAsynthesisand the synthesis of single-stranded minus strand DNA that becomes encapsidated into progeny virions (17, 37).
The virion DNA containsself-complementary sequences atboth its 3' and 5' termini,and RF DNAhas been shown tocontain these terminal palindromes in both extended and foldback
con-figurations (3, 5, 19). The 5'termini of both the plus and minus strandsofRFDNAare covalent-ly bound to a protein(s) whose function is un-known (15). The pools of RF DNA extracted from infected cells are also characterized by a
significantfraction(=25%)ofdimer-length mol-ecules that arelinked at the 3' terminus(18). The 3' termini of several parvoviruses have been sequenced, and evidence was discussed which indicated thatthe orientation ofthe3'-terminal
palindrome is unique, that is, not inverted, in both virions andRF DNA(1, 2).
Inthisreportwepresenttheprimarystructure
of the 5' terminus of the parvovirus H-1 and
t Laboratorium voor Fysiologische Scheikunde, Leiden,
The Netherlands.
portions of the terminal regions of DI-1, an
addition mutant of H-1, and H-3, aparvovirus related to H-1. This region of themolecule has previously been shown by us to contain the
replication origin for RF DNA replication (26,
27). Our evidence indicates that the5'-terminal
palindromeis present in two orientations inRF
DNA, one inverted with respect to the other. This ffip-flop relationship suggests that the mechanism for synthesis of 5' termini of DNA proposed by Cavalier-Smith applies to this end
ofparvovirusRF DNA(4).Finally,wepresenta
model forRF DNAreplicationthatis
compati-ble with the currentlyavailable factspertaining
tothis process.
MATERIAUSANDMETHODS
Materials.Restriction endonucleases were obtained
from New England Biolabs or Bethesda Research Laboratories andwereusedasdirected by the
suppli-er.Thelargefragment of Escherichia coli DNA poly-meraseI(pol I) was obtained from Boehringer-Mann-heim. T4 DNA ligase was from New England Biolabs. The deoxynucleotide triphosphates and dideoxynu-cleotide triphosphates were from P-L Biochemicals, Inc. The HindIII linker and theM13-specific primer werepurchased fromCollaborativeResearch, Inc. [a-32P]dATP was from New England Nuclear.
Cells and viruses. The parvoviruses used in this
study were wild-type H-1, H-1 DI-1, and H-3 (20). Virus and RF DNA were obtainedfrom infected NB
cellsaspreviously described (19). M13 mp2, mp5, and
mp7, and JM103, werekindlyprovidedby J.Messing.
Moleculardoning ofparvovirusDNA. Approximate-ly 20 ,ug ofparvovirusRF DNAwasdigestedwith the appropriate restriction endonucleaseaspreviously de-scribed (19). The reactions were terminated by the addition of 50 mMTris, pH 8.0, 0.1 MNaCl, 1 mM EDTA, and 0.2%Sarkosyl,to afinalvolume of 200
tl,
andwereextracted oncewith phenol andonce with 990
on November 10, 2019 by guest
http://jvi.asm.org/
phenol-chloroform-isoamyl alcohol (25:24:1). Phenol was removed by ether extraction; the solution was
adjustedto0.3 MNa+with 3 M sodium acetate, pH 5.5, and the DNA was precipitated with ethanol. HindII or HincIl digests were ligated to a l1-mer HindIll linker as described (8). In some cases the linkerswerephosphorylated withunlabeled ATP, and themobility shift of the fragmentduring its isolation by gel electrophoresis providedamonitorof the effective-nessof the linker ligation step. Reactionswerecarried
out in 66 mMTris, pH 7.4, 6.6 mMMgCl2, 10mM
dithiothreitol, 1 mMspermidine,0.5 mMATP,3,uM
linker,50,ugof bovineserumalbumin perml,and 10 to20 Uof T4 DNA ligase per mlat14°Cfor 20 h. The reactionswerestopped byheatingat65°Cfor 10 min. The reactionmixture was adjusted to100 ,ul and 60
mM NaCl-0.03% sodium azide and was incubated
with 10 U ofHindIIl for 16 h at 37°C. The digestion was terminated by the additionof EDTAto20mM, and the DNA was precipitated with ethanol. The restrictionfragments were isolated byelectroelution after fractionation by electrophoresison agelof 5%
acrylamide. Slightlybroadened bands wereobtained
atthegel positions expected after the addition of the linker sequences. After ethanol precipitation, the
DNA fragments were ligated to HindIII-cleaved
pBR322 or, in thecaseofHincIl 83/91, directly into M13mp5by standard methods (8). E.coli HB101 was
transformedbythe methodof Dagert and Ehrlich(6). Recombinant plasmids were identified by screening the Apr Tcscolonies for the proper size insertionsby
gelelectrophoresis.Plasmid DNAwasprepared bythe
extraction method ofGuerryetal. (10),concentrated
by precipitation with 10%o polyethylene glycol,
ex-tractedwithphenol, andpurified by centrifugation in
CsCVlethidium bromide gradients. Parvovirus
frag-ments weresubcloned in M13mp5 bydigestionof 0.5
to1 p.gof plasmid with HindIII and ligation of the
fragmentsto50to100 ngofHindIII-cleavedM13mp5
RF DNA. Wedidnot recoveranypBR322-M13mp5 recombinants.
TheHincIl 95.7/99.9 fragment of H-1was isolated
from pBR322 by gel electrophoresis and restricted
with TaqI; the single-stranded endswerefilled inby
treatment with the large fragment ofE. coli pol l.
HindIll linkerswere addedas described above, and
thefragmentswereisolatedby gelelectrophoresisand
finallywerecloned in M13mp5.
The87/97.5fragmentof H-1wasobtainedbycloning
theTaqI 38/97.5 fragmentin M13mp7atthe AccI site.
RF DNA of this recombinant was restricted with Sau3A, whichcuts H-1 atmapposition 87 and M13 mp7atits Bamsite. The properfragmentwasisolated
by gel electrophoresis and clonedatthe Bam site of
M13mp7.
The strands represented in the M13 clones were
determined by incubating approximately 1 p.g of recombinant single-stranded DNA with 1 pgof H-1 minus strandDNAin SxSSC(SSC:0.15MNaClplus 0.015 Msodiumcitrate)at65°C for1to2h. Plus strand recombinants hybridize to H-1 DNA and show a
marked reduction of mobility in 1.6% agarose gels. Some clones contained tandem repeats of the
frag-ment.Inverted repeatswere notrecovered.Organisms
containing parvovirusrecombinantDNA were
propa-gated under Pl-EK1 conditions in accordance with the Guidelines for Recombinant DNA Research.
DNAsequencing. DNA sequencingwas done by the chain-terminating method of Sanger et al. (23, 24). We used either the 96-base-pair (bp) primer asdescribed (11) or a 12-bp synthetic primer. Electrophoresiswas in 0.3-mm gels of 8% acrylamide at about 50°C (29). Most gelswere 27 by 40 cm, buta few were 15 by60 cm.Samples were applied in two loadings 3 h apart.
RESULTS
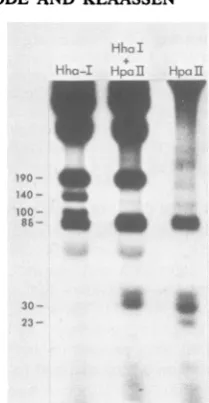
Orientation of the 5'-terminal palindrome. The physical map ofH-1 RF DNA was determined for therestriction endonucleases HhaI andTaqI by partial digestions and reciprocal digestions with enzymes having known sites, as previously described (19) (Fig. 1). These studies revealed two smallfragments generated by both of these enzymes that were present in less than equimo-lar amounts and which mapped to the 5' termi-nus of the molecule. This can be illustrated by the double digestion of RF DNA withHhaI and HpaII(Fig. 2). HpaII is known to have a number of cleavage sites, all located in a small area of the 5'-terminal palindrome (19). The twoHhaI
fragments of about 140 and 100 bp were both cleaved by HpaII, localizing them to the right end. Isolation of the HpaI 91/100 fragment and then digestion with HhaI produced four frag-ments of 100, 140, 340, and 380 bp (data not shown). The sums of the first plus fourth and the second plus third of these equal the original
480-bp 91/100fragment. The simplest interpretation of these results is that the 5' terminus has HhaI sites at either 140 or 100 nucleotides from the end in approximately equal proportions.
Fur-thermore, we would predict that although these
sites are within the terminal foldback they are not in a palindromic portion of it which has its axis of symmetry at nucleotides 115 through 120. This interpretation was confirmed by
demon-strating that HhaI did not cleave this region when in the foldback configuration (data not
shown) and more directly by DNA sequencing
as described below. A similar result was
ob-tained with TaqI except that its cleavage sites were located about 135 or 105 nucleotides from the 5' terminus. Thus, these data indicate that the 5'-terminal foldback region ofH-1 exists in
two orientations inverted with respect to each
Hh
I8
0.98.190
205TaqI 19.6
53.0 56.5
38 \0 / 72.5
I r/ 38.5
97.4or98.1 97 5
97.5or98.0
FIG. 1. Physical map ofwild-type H-1 RF DNA for the restriction endonucleases HhaI and TaqI. It is assumed thatthe terminalpalindromesareextended, with map position 0 at the 3' end of the minus strand and 100atthe 5' end. TheHhaI sitesat0.8 and 0.98
were obtained from the DNA sequence (1). The
re-maining sites were derived by partial digestion and
double digestions with enzymes with knowncleavage sites.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.491.254.443.564.598.2]ci
Hhoc-; tlp H1jeii
190-t40
-100- _
30- 1
23
-FIG. 2. Electrophoresis patternof32P-labeledH-1
RF DNA fragments produced by: HhaI, HhaI + HpaII, and HpaII. Fragment sizeswereestimated by
usingaHaeIII digest of+X174RFDNAinanadjacent
lane.Electrophoresiswasina15%acrylamide gel with
a 1-cm top layer of 5% acrylamide containing the
sample slots for 6 hat150 V. The gelwasstained with
ethidiumbromide, photographed, and then driedfor
autoradiography.
other such that there are two alternative
posi-tionsforaHhaIorTaqI site thatareequidistant
from the axis ofsymmetry.
DNA sequence of the 5' terminus of H-1.' To
determine
the
structureof the 5'-terminalpalin-drome and adjacent regions ofH-1, we cloned theHindll 95.7/99.9and91/95.7fragmentswith
Hindllllinkers into theplasmid pBR322. Subse-quent subcloning ofthe 95.7/99.9fragment into the single-stranded phage vector M13 mp5
re-sulted in a deletion of about 500 bp from the
recombinantsathighfrequency.Thisinstability
was circumvented by subcloning 95.7/99.9 in two fragments using the TaqI site at 97.5 (for
purposes of discussion only one orientation is
assumedhere),thusdividingthepalindromeinto parts. These fragments were sequenced by the
chain-terminating method ofSanger etal. (24), andarepresentative gelisshowninFig. 3.The
polymerizationreaction encountered avery
in-hibitory G-rich sequence in the palindrome
when the reaction was in the direction 97.5 to
95.7 or97.5 to99.9, and itwas notpossibleto
read the sequence past this point on these
strands. The sequencing reaction was often
slightly ambiguous atthis site on the opposite
strands as aresultofanunderrepresentation of
someof the Gbands,asshownin Fig.3a,buton some gels these regions were clearly deter-mined, e.g., Fig. 3b. Another complication is that the 97.5/99.9 fragment contains a small region with a9-bp palindrome with two HpaII
sites thatyieldedadifferentsequencedepending
onwhich strand of the subcloned fragmentwas
thetemplate (see Fig. 5). Threedifferentclones
were sequencedin the 5' to 3' direction 99.9 to
97.5 and two clones were sequenced from 97.5
toward99.9withthe same result. The sequence
derived in the 99.9 to 97.5 direction predicts a
HpaIIfragment of23 bpthat is cleaved by HhaI.
Thealternativesequence,whichis shown in the
legend of Fig. 5, would have HhaI cleaving a
HpaIIfragment of36bp. The results of such a
digestion (Fig. 2) indicate that the former
se-quence is the correct one. The other sequence
represents aninversionof 28 nucleotides (except
theone underlined inthelegend of Fig. 5). The
sequence change in the fragment must have
occurred during propagation of the original
HindII 95.7/99.9 fragment cloned in pBR322or
during the subsequent subcloning in M13 mp5. This inversion is not tobe confused with that occurring duringtheparvovirus RF DNA
repli-cationdiscussed above.
The HindII 91/95.7 fragments of H-1, the addition mutant, DI-1, and H-3 were similarly cloned inpBR322 and subcloned in M13 mp5. In addition,aRsaI siterevealedby sequencingwas
used to subdivide this region further for the sequencing of H-1 and DI-1. The sequencing strategy used is summarized inFig. 4, andthe sequence for wild-type H-1 minus strand is presentedinFig. 5.
The first seven nucleotides atthe 5' terminus werenotdirectly determinedbut are inferred for
reasonsofsymmetry sinceit ishighlylikely that
they belong tothe palindromic portions ofthe terminus which includes the HindII sites at
positions7and236. Because the 5' terminus was
FIG. 3. (a)Sequencing gelof theHindII-TaqIfragment95.7/98.0(nucleotides135to235)in both directions.
TheG-richregionat188to201producesastrongstoptothepolymerizationreaction,preventingconfirmation of
the sequencebeyond thatpoint(2). ThecomplementaryC-richregionwhich isadjacenttothepile-upbanddoes
notslowpolymerization(3).Reading from bottomtotop,panel1readsnucleotides170to135(plusstrand), panel
2reads 135to178(minusstrand),andpanel3reads 235to180(plus strand;numbersrefertothoseofFig. 5).On
panel2the H-1 sequencebeginsafter the TTGG of theHindIII linker.Inpanel3theoriginal HindIIIlinkerwas
filled.inbypolIandasecondlinkerligatedsothat theH-1 sequencebeginsafter the TTGGAGCTTG.Itwas confirmed by sequencingacrosstheHindll siteat235 andusingtheRsaIfragment (nucleotides157to313)that
oneGwaslost from the linkerduringthefirstcloningofHindII95.7/99.9.(b)Sequencefrom nucleotides 34to 105 across the palindrome at 41 to 57. Symbols: aC, arabinofuranosyl cytosine; A, deoxyadenosine; C,
deocycytidine; G, deoxyguanosine; T,thymidine.
VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.491.100.205.65.267.2]TERMINUS
2
3
a
CACOGT
Am~~~~~~~~~~
AG
|
K
s
a*G
TTTT^G9,~~~
-a
CACOGT CA C GT C A C G T
a
1
41,
I
on November 10, 2019 by guest
http://jvi.asm.org/
Toq I HindII Rso I
136 SS 235 312
HindEl Sou3A matedfor the 3'terminus of parvovirus mRNA
43' species(9, 30), buttwoother AATAAAsignals are present in this region. This region is also
FIG. 4. Sequencingstrategy used for determining
the 5'-terminal sequences of parvovirus H-1. The Hindll fragments: nucleotides7to235 and235to488
werecloned inpBR322 withHindIlI linkersand 488to
=880wascloned inM13mp5withEcoRIlinkers.The HindlI-TaqI fragments7-136 and 136-235and HindII-RsaIfragments235-312 and 312-488weresubcloned in M13mp5.The Sau3Afragment136-640wassubcloned
inM13 mp7fromaTaqIfragment136--3,270cloned atthe AccI site inM13 mp7. The symbolssindicates
theG-rich palindrome thatactedasastrong stoptothe
sequencing reactions. Theclone marked withan aster-isk had a region thatwas inverted within it, as
dis-cussedinthetext.
covalently bound to protein and remained blocked after deproteinization, we have not
identified the terminal nucleotide precisely. Its approximate location wasderived from the
ob-servationthat restriction of the terminal HpaII fragment with Hindllcauses itto shift down in mobility by approximately7bp(19).Therefore,
we are placing the 5' terminus tentatively at
sevennucleotides from the first HindII site.
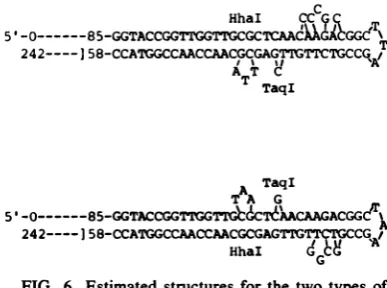
The most likely foldback structure that we
predict for the 5'-terminal palindrome in virion
DNA, usingthe methods of Tinocoetal.(32),is
shown inFig. 6. Thestemof thehairpin,
nucleo-tides 1 to 100 and 143 to 242, is a perfectly
matchedpalindrome, and only the portion
con-taining the asymmetries is shown in Fig. 6. A T-shapedstructureis also possible if thesequences
from 109to 119are pairedto those at89to94 and149to154, but this is lesslikely since it hasa
lower freeenergy.Also, therestriction ofnative
RF DNA with KpnI, which cleaves at 76 and within thepalindrome, produced onlyoneband onagarosegelsattheposition expectedfor the
76/97.1 fragment (data not shown). If the
fold-back were inthe T-shaped structure, the KpnI
sitewouldbeatthe branchpoint ofthe T andnot
cleaved.Thiswouldresult intwobands,namely
the 76/97.1 and 76/100*, as found with the
HaeIII 83.5/100 bands (19). Thesequence
con-firms thepredictedasymmetries oftheHhaI site
at100 andthe TaqI site at136.
The 88/95 region immediately adjacenttothe terminal foldback is a noncoding region ofthe
genomebecause therearechain-terminating
co-dons inallthreereadingframes between
nucleo-tides 600 and 640. A canonical polyadenylate
signalof AATAAA(plus strand) followedby CA
21 nucleotides downstream is present at 277.
This is compatible with the map position
esti-HindII 10 20 30 40 50
51-TTCAGTTGAC CAACTGAACC TATAGTATCA CTATGTTTTT AGGGTGGGGG 60 70 80 K_nI9O HnaII 100
GGTGGGAGAT ACATACGTTC GCTATGGACC AAGTGGTACC GGTTGGTTGC
HhaI 110 HpaII 120 130 TaqI 140 150
GCTCAACCAA CCGGACCGGC TTAGCCGTCTTGTTCGAGCT TAGCAACCAA
HpaII
KpnI 160 170 180 190 200
CCGGTACCAC TTGGTCCATA GCGAACGTAT GTATCTCCCA CCCCCCCACC 210 220 230HindrI 240 250
CTAAAAACAT AGTGATACTA TAGGTTCAGT TGGTCAACTGAATAGACAGT
260 270 280 290 300
AGTCCTTGTT CATTTTAATT ATTTTATTAT TTTTTTGGTCCCTAACATTC
310 RsaI 320 330 340 3S0
AGTCTAAGGG AGTACCAACC AACCACCCAA CCACCCTTTTTATTTCCC>A
360 370 380 390 400
ATACAAAATC TTTTTATATT CTATCTAACA TTAACCAACT AACTATATTT
410 420 430 440 450
ATTTTAAAATACAAAATCTTTTTATATTCT ATCTAACATT AACCAACTAA
460 470 480 HpaI 490 500
CTATATTTAT TCCAAAAGTTCTATTATTTTTTACAGTTAA CCTACCATAT
-j
510 520 530 540 550
TATGTTATTT TTAATAATCTATTATTAAGT GTATATTTTT ATGTTGTAGT
560 570 580 590 600
CTAGTTAGTT TAAGTATTAT GTGAAGCAAACAGAGAAACATAGTTGGTTG
610 620 630 Sau3A
[image:5.491.56.250.56.120.2]GTTAGTATGT CATGTGAGGC ACAGGTCTAC AAATCAATGGATC-3'
FIG. 5. The nucleotidesequenceof the 5' terminus of theparvovirus H-1. Thefirst fournucleotides are
inferredfrom the expected symmetryof the inverted
repeatencompassing nucleotides 0to100and 143to 242,andwerenotdirectlydetermined. The5'-terminal nucleotide was bound to a protein; its removal by protease digestion leaves a 5' terminus blocked to labeling by polynucleotide kinase. Its location was
estimatedbythemobilityshifttotheHpaIIterminal
fragmentafterdigestionwithHindll. Thesequencesin
thebracketsarethe55-bp tandemrepeatsofwild-type H-1. The viable defective mutantofH-1, DI-1,was
sequenced for themapregioncorrespondingtoHindII
236to 488. DI-1 containedthree copiesof the 55-bp
repeatsequencewithtwointerveningTTA.Similarly,
the parvovirus H-3 was shownto contain only one copyofthis sequence. H-3 also differed fromH-1 at three otherpositionsinthisregion: nucleotides291 =
T, 362 = G, and 479= G. The alternative sequence
obtained from twoclones marked in Fig. 4 withan
asteriskwas
100 HpaII HpaII
GCGCTCA-ACAAGACGGCTAGCCGGTCCGGTT-TaqI GGT-TCGA
This sequence (between the dashes) is the inverted complementof thatinFig.5exceptfor the I,which is
the same. The chain terminators in three reading
frames between 600 and 630 are underlined. The
terminator at 602 ends the only large open reading frame,soitverylikely marks the carboxy terminus of
the H-1coatproteins (Rhode, unpublished data).
Hind 0
7 SS
5'1
J. VIROL.
4 lb64 lo
ke ---w .4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.491.264.451.119.382.2]PARVOVIRUS 5' TERMINUS 995
5'-O---85-GGTACCGGTTGGT
242----] 58-CCATGGCCAACCAI
5' -o---85-GGTACCGGTTGGT1 242----]58-CCATGGCCAACCAJ
FIG. 6. Estimated structure
5'-terminal palindromes in th4
tions(32). The nucleotides 0tc
completely base paired. This s
by a reviewer of this manusc
lowerfreeenergy than the one
us.
distinguished by its richne4
two regions of entirely A-I
and471 to 481. H-1 contain
55 bp (348 to 402, 406 to 460
TTA. The addition mutant
tandem repeats of this seqi intervening TTA, and H-3
copyand is missing the inte
thanthe repeat sequence,H
gous to H-1 since it differe
tionsbetween 235 and488.
DISCUSSII
The fortuitous asymmetr TaqI sites in the
5'-termin;
DNA have demonstrated t]contains approximately eqi
types ofterminal palindron
complement of the other. was derived from a freshl
finding indicates that thefi ofthe 5'-terminal palindron ess, but it does not prove
t}
each round of replication. been established for the c
adeno-associated virus (12
strongly support the
foldbE
forgenerating5' termini de Smith (4). In the case of the
viruses,this mechanism
app
one end of the DNA, the minus strand. Because oftithe mere existence of coval toplus strands at the 5' tern ingevidence in favor of a I nism ofinitiation of RFD? 31).
Ahypothetical replicatior
HhaI CCC ratesthe knownfeatures of
parvovirus
RFrepli-rGCGCTC_GA
GGC
cation is outlined in Fig. 7. In particular, it CGCCGAGTTTCTGCCG depends on thefinding
ofY-shaped replicative
AT
T C intermediates described forH-1
(26, 27). TheseTaqI data
conffict
with the failure of Tseng et al. todetect Okazaki fragments in nascent
H-1
DNA (33). Resolution of thiscontradictionwill
require further study. The synthesis of parental RFTaqI DNA is hypothesized to initiate on the native 3'
TNA
G terminus of virion DNA, step 2 (3). Directrk6CT&ACAAGACGGc!"\
evidence of this is lacking, but it has been shown',CGCGAGTTTCTGCCG,
that virion DNA chases intoRF
DNA with aHhaI
d
9G
covalently
closed3'-terminal
foldback(35).
The transcription of parental RF DNA has beenesr
foldthe
totnfigeuraf
inferredbyindirectmethods through itssensitiv-D85 and 158 to 242 are
ity
toinhibition by
substitution ofbromode-.tructurewas suggested
oxyuridine
forthymidine during parental
RF:ript
and has a slightlysynthesis
(17, 37).
originally
proposed by One principal feature ofthis model is that it utilizes a Cavalier-Smith type foldback transfer process to generate the 5' termini of minusstrands,
which is shown at step 5 or 9. A second ss in AT (73%) and feature is that an asymmetrical processing of the pairs at 261 to 285 minus strand3'-terminal
palindrome is executedIs
a tandem repeat of on a dimer substrate for generating the 5' termini ) with an intervening of plus strands. In this way the unique orienta-DI-1 contains three tion of the3'-terminal
palindrome of minus uence, including the strands in virion DNA and RF DNA is con-possesses only one served as previously reported (1), steps 5 and 6.~rvening
TTA. Other At step 3 replication is presented as an1-3is highly homolo-
internally
primed event on the plus strand d at only three posi- template ofRF DNA with the 5' foldback region in its extended duplex configuration. There is no direct evidence to support or refute thishypo-'ON thesis at this time. This step may be analogous
ies of the
HhaI
and to the D loop of mitochondria. In fact, we are al region ofH-1
RF proposing that the function of theG-rich
se-hat native RF DNA quence in the plus strand shown as f in Fig. 7 ual amounts of two prevents minus strand synthesis on the 5' side of ne, one the inverted this site as suggested for mitochondrial DNA Since theRF
DNA replication (25). It is interesting that this site in y cloned virus, thisH-1
is a G-rich palindrome, AGGGTGGGGlip-flop configuration GGGTGGGA, that is remarkably
similar
to the ne is an active proc- sequence AGGGGGTGGGGGGGT found adja-iat it happens during cent to the origin of H-strand synthesis in rat This result has also mitochondrial DNA (25). By inhibiting DNAlefective parvovirus synthesis, this signal sequence would ensure ), and both results that the plus strand palindrome remains free to ack transfer process renature as a foldback structure when initiation scribed by Cavalier- and rightward chain extension on the minus nondefective parvo- strand template drives the separation of the ears to apply to only parental foldback region, as shown in steps 3
5' terminus of the and 4.
Progression
of the replication fork toward he foldback transfer, the covalently closed 3' foldback converts thelentlinkage of minus monomer
RF
to a dimer, step 5. Synthesis of the ninus is not convinc- plus strand would be by a discontinuous method DNA-primed mecha- that presumably uses Okazaki fragments as NA replication (e.g., intermediates.As an
alternative,
a DNA-primed initiationamodel that incorpo- using the plus strand 3' terminus as primer
VOL.41, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.491.46.240.73.217.2]1.
b
2.
cQa:
5.
5'-f f' f' f
6. 5'
f f'
f' f a' b' c' c d' a 7. 5'
5f f -a b c c' d a'
IRFrep
f', f
o f f'
L
b a
I
f f _d" 5'afi
doa
f fU8.
[image:7.491.110.402.70.437.2]9.
FIG. 7. Hypothetical model for thereplicationofparvovirusDNA. Theviral DNA is notdrawn to scale. Complementarysequencesarerepresentedbysmalllettersandtheirprimes.Abbreviations: RF rep,transacting
viralproductrequired for RFreplication; VP,viralproteins;N1orN2site-specific single-strandendonucleaseor
nickase; Pol,DNA polymerization; L, ligation; f,thepalindrome sequence GGGTGGGGGGGTGGG. All or
nearlyallofthe5' terminiareboundtoaprotein(15).Step1depictsthestructureof virion DNA. Atstep2the synthesisofparentalRF DNA hasoccurred,andtheparentalRF istemplatefor viral mRNAsynthesis. When RFrep orvirionproteins,orboth,aresynthesized,RFreplicationproceedsatstep3. Initiation isproposedto begin with minus strand DNAjust inboard of the terminal inverted repeat sequences. An early replicative intermediateisshownatstep3.Progressionof thereplicationforktothe 3'hairpinconvertstheparentalRFtoa
dimer RF. A foldback transferprocess proceedsatone ofthedaughter molecule's 5' termini toproduce a5'
terminus for thenascentminus strand insteps 5 and6. Thisisrepresentedbynickase N1cleavingtheplusstrand
toproducea3'-OHprimersite for stranddisplacement synthesiswhich extends the foldback. Inasimilarmanner
nickaseN2cleavesattheboundaryof the 3'hairpinsequences(step 5),but stranddisplacementsynthesisoccurs
atonlyoneof thesesites(step 6).Instep7 thedimer has converted to twomonomerRF DNAmolecules,and the
orientation of the 3'hairpinremains thesame asfor theparentalvirion DNA. RF DNAwiththe extended3' end replicatesasinsteps3 and 4except that twomonomerRFmoleculesaregenerated(steps 7-9). MonomerRF
DNA with the3' end in thehairpin configurationmustrecyclethroughthe dimer intermediatewhen itreplicates.
Thismodelwasformulatedtofit thefollowingexperimentaldata:(i)thereplicativeintermediates ofRF DNAare
double-stranded branched molecules(26); (ii)theorientation of the 3'hairpinisunique (1); (iii)therearetwo
orientations of the 5'hairpin (this work; 1); (iv)dimer moleculesareprominentmembers(25%)of thepoolof replicatingRF DNA(18, 26, 27); (v)asignificantfractionof the 3'foldback structuresarecovalentlyclosed(18); (vi) onlythe extended andfoldbackconfigurationsof the 5' terminus have been identified(19).Arabbit-eared5'
terminushas notbeen detectedby gel electrophoresis of 5'-terminal restrictionfragments. _f'
5 f
_ f
If
J.VIROL.
f 5
f'
on November 10, 2019 by guest
http://jvi.asm.org/
997
would require a mechanism for converting the extendedstructureof the palindrometo a "rab-bit-eared"structure. Wehaveproduceda termi-nal fragment tentativelyidentified as a
"rabbit-eared" structure after denaturation and renaturation of the native terminal fragment. There is no evidence for such a terminal frag-ment in native RF DNA (S. L. Rhode, unpub-lished data).
The foldback transfer processes which are symmetrical at the 5' terminus, that is, indepen-dentof the ffip or flop orientation, and
asymmet-rical for the 3' terminus are shown as a site-specific nicking reaction by nickase N1 or N2
followed by strand
displacement
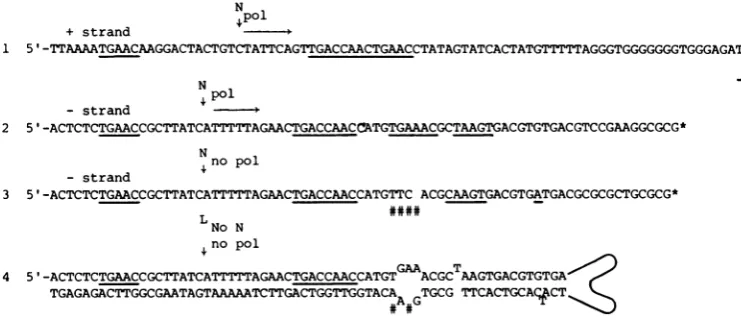
by the DNA replication apparatus,!°.Therefore, DNA repli-cation itself would drive the strand separations necessary for these reactions. Examination of the reported sequence for the various 3'-termi-nal structures of H-1 and comparison with the appropriate region of the 5' terminus suggests how the template could direct these reactions(Fig. 8). Sequence 1 is the plus strand sequence attheborder of the 5'foldback with the foldback
turnaroundtothe right. Sequences 2 and 3 are the twoportions of minus strand sequences from thedimer junction, that is, the abc and ad'c of
Fig. 7, and sequence 4 is the 3' terminus in foldback configuration. The minus strand 3'-terminal sequences at the monomer junctions in the dimer are shown one above the other for easeofcomparison with the axis of symmetry at therightindicated by an asterisk. The tentative
nickingandprimer sites for strand displacement
are again indicated by N !$ A most striking feature of all four sequences is thehomologous positions of the possible recognition signals
TGAAC, TGACCAAC, and
TGAA(k)C
withrespect to thepostulated nicking sites. Equally
important, the 3' foldback region of the dimer, which we have suggested is nicked but is not a substrate for strand displacement synthesis, is missing the TGAAAC sequence present in the other two templates. Similarly, the same signal
site in thefoldbackconfiguration(4) is located at an interior loop rather than a duplex site, as
found in the dimer. This model proposes that
this configuration isprocessed byligationrather thannicking. Therefore,themodelpredictsthat a duplex
TGAA(k)C
signal is required for the strand displacement DNA synthesis. Thus, these regions contain the required symmetriesandasymmetries of sequence necessaryto regu-late the foldback transfer mechanism as de-scribed in the model. It is possible that the nicking enzymes N1 and N2 are the same even though the nick sites do not seem to have identicalrelationships to thepostulated
recogni-tion signals. It is alsopossible that the terminal boundproteins are the nickases (15).
This modelpredictsarecyclingofsomeofthe monomer RF DNAs through adimer molecule and allows someof the monomer RF DNAs to
replicatewithout a dimerintermediate, steps 7-9. Dimers arepostulated to beformed by
repli-cation and notrecombination.This is supported
by our finding that the dimersproducedincells doubly infected with standard virus anda
defec-+ strand
Npol
1 5'-TTAAAATGAACAAGGACTACTGTCTATTCAGTTGACCAACTGAACCTATAGTATCACTATGTTTTTAGGGTGGGGGGGTGGGAGATA
Npo1
- strand
2 5'-ACTCTCTGAACCGCTTATCATTTTTAGAACTGACCAACCATGTGAAACGCTAAGTGACGTGTGACGTCCGAAGGCGCG*
Nno pol
- strand
3 5'-ACTCTCTGAACCGCTTATCATTTTTAGAACTGACCAACCATGTTC ACGCAAGTGACGTGATGACGCGCGCTGCGCG*
No N
+no pol
4 5'-ACTCTCTGAACCGCTTATCATTTTTAGAACTGACCAACCATGT ACGC AAGTGACGTGTGA
TGAGAGACTTGGCGAATAGTAAAAATCTTGACTGGTTGGTACAA
TGCGTTCACAQPCT
FIG. 8. Comparison of the 5'-terminal (1) and3'-terminal(2-4) sequences in theregion of thehypothetical sitesfor the nickasesN1and N2ortheligation (L) of thereplication model of Fig. 7. Sequence1is the plus strand sequenceattheboundary of the5'-terminal palindrome. Sequences 2 and 3 are the minus strand sequencesatthe dimer RFjunction brought into vertical alignment with the axis of symmetry indicated by the asterisk. Sequence
4is the 3' terminus in itsfoldback configuration (1). The # signs below the line mark points of asymmetry where theoneside of the dimer
sequence
(3) or thefoldback sequence (4) does not haveaduplexTGAAAC which we postulatetoberequired for the strand displacement, P°, to occur. The exact site of nicking for sequence 1 awaits theprecise placement of the 5'-terminal nucleotide.VOL.41,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.491.50.421.438.600.2]RHODE AND KLAASSEN
tive deletion mutant are of two types: standard length x2 or deleted length x2 (Rhode, unpub-lished data). This conclusion should be tem-pered by the fact that we cannotprove that the replicating pools of standard and deleted mole-cules are not compartmentalized within the nu-cleus, thus precluding recombination between them(28).
Because of the symmetry within 100 nucleo-tides of the nick site at the 5' terminus, one might expect that the nicking-strand displace-mentmechanism might not distinguish between 5' termini in foldback and extended configura-tions. If this is the case, this process acting on extended termini would strand displace free copies of the 5' hairpin as a wasteful side reac-tionof the replication process, and we intend to search for these. Otherpredictions of this model which can be tested are that the 5' termini of
dimermolecules willbe in the same orientation (step 6, Fig. 7) and that inhibition of DNA
synthesis will block the interconversion of the various terminal structures.
Finally, the model does not show progeny
minus strand synthesis. Virion DNA synthesis couldoccur by aninteractionof anappropriately mature viral capsid with an early replication intermediate (justbefore step 3 or 8) in such a way as to block theantipolar plus strand synthe-sis. Evidence has beenpresented which suggests thatencapsidation proceeds in closeconjunction with the displacement of minusstrand from the RFtemplate (7).
The sequence derived for the 5' terminus of H-1 includes the replication origin which maps to this site by electron microscope analysis of replicative intermediates(26, 27). The sequence data haveconfirmed ourearlierhypothesisthat the addition in DI-1 is a tandem repeat of a sequence that is repeated already in wild-type
H-1 andis present inonlyonecopy in H-3(20).
A similar relationship has been suggested by physicalmapping data for minute virus of mice (MVM) and MVM(i) (14). Itis thusverylikely
that the further additions reported in H-1 DI genomes between map positions 91and95.7are
tandemrepeatsof this same sequence (21). What role this sequence has in DNA replication is unknown, but it is clear fromtheexample of H-3 that only one copy is required for efficient RF DNAreplication. The tentativeorigin region has many featuresreported for otherorigins ofDNA
replication. The noncoding region inboard of the terminalpalindrome 244 to 640 isA-Trich(73%)
and contains a 25-nucleotideATblockat 262 to
286 andan11-nucleotideATblockat472to 482. Italsocontains a23-nucleotide AC stretchat314
to 336. There are true palindromes of eight
nucleotides orlargerat3 to10,25to35,41 to57,
111to119,186 to202,208 to218,233 to240,267
to 278, and 313 to 324. We are attempting to develop a functionalassay for origin activity so thatstructure-function studies can be carried out todetermine which features of this sequence are important in the initiation of RF DNA replica-tion.
ACKNOWLEDGMENTS
We gratefully acknowledge Jeanne Helft for expert techni-calassistance,Virginia Haas forsecretarial duties, and Elea-norSpicer for helpful discussions.
This work was supported by Public Health Service grants CA-25866andCA26801 fromtheNational CancerInstitute.
LITERATURE CITED
1. Asteil, C., M. Smith, M. Chow, and D. Ward. 1979. Structureof the 3'hairpintermini of four rodent parvovi-rusgenomes:nucleotide sequence homology at origins of DNAreplication. Cell 17:691-703.
2. Asteil, C., M. Smith, M. Chow, and D. Ward. 1979. Sequenceof the3'-terminusof the genome fromKilham rat virus, anon-defective parvovirus. Virology 94:669-674.
3.Bourgulgnon,G., P.Tattersall, and D. Ward. 1976.DNA of minutevirusofmice:self-priming, nonpermuted, sin-gle-strandedgenomewith a5-terminal hairpinduplex. J. Virol.20:290-306.
4. Cavalier-Smith, T. 1974.Palindromic basesequencesand replicationof eukaryote chromosome ends. Nature (Lon-don)250:467-470.
5. Chow, M. B., and D. C. Ward. 1978.Comparison ofthe terminal nucleotide structures in the DNA ofnondefective parvoviruses, p. 205-217. In D. C. Ward and P. Tattersall (ed.), Replication of mammalian parvoviruses. Cold SpringHarborLaboratory, Cold Spring Harbor,N.Y. 6. Dagert, M., and S. D.Ehrlich.1979.Prolongedincubation
in calcium chlorideimproves the competence of Esche-richiacoli cells.Gene 6:23-28.
7. Gautschi, M., and P. Reinhard. 1978. Synthesis of a
mammalian parvovirus onBrij 58-lysedcells. J. Virol. 27:453-456.
8;Goodman, H. M., and R. J. MacDonald. 1979.Cloningof hormone genes from a mixture of cDNA molecules. MethodsEnzymol.68:75-89.
9. Green, M., R. Lebovitz, and R. Roeder. 1979.Expression
ofthe autonomousparvovirusH-1 genome: evidencefora
single transcriptional unit and multiple spliced polyad-enylatedtranscripts.Cell17:967-977.
10. Guerry, P., D. J. LeBlanc, and S. Falkow.1973.General methodfortheisolation ofplasmiddeoxyribonucleic acid. J.Bacteriol. 116:1064-1066.
11. Heidecker, G., J. Mesdng,andB.Gronenborn. 1980. A versatile primer for DNA sequencing in the M13mp2 cloningsystem.Gene 10:69-73.
12. Lusby, E., K. H.Fife,andK. I.Berns.1980.Nucleotide sequence of theinverted terminalrepetitionin adenoasso-ciated virus DNA. J. Virol. 34:402-409.
13. McGeoch,D. J.,L.V. Crawford, andE. A.C. Foilett. 1970. TheDNAs of threeparvoviruses. J. Gen. Virol. 6:33-40.
14. McMaster, G. K.,P.Beard,H. D.Engera,andB.Hirt. 1981.Characterization ofanimmunosuppressive
parvovi-rusrelated to minute virus ofmice. J. Virol. 38:317-326.
15. Revie, D., B. Tseng, R. Grafstrom,and M.Goullan.1979. Covalent association ofproteinwith thereplicativeform DNAofparvovirus H-1. Proc. Natl.Acad. Sci. U.S.A. 11:5539-5543.
16. Rhode, S.L.1974.Replicationprocess of theparvovirus H-1.II.Isolation andcharacterizationof H-1 replicative form DNA. J. Virol. 13:400-410.
17. Rhode, S.L.1974.Replicationprocess of theparvovirus H-1. III. Factors affecting H-1 RF DNA synthesis. J. Virol. 14:791-801.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
18. Rhode, S. L. 1977.Replicationprocessoftheparvovirus H-1. VI.Characterizationof areplicationterminus of H-1 replicative-form DNA synthesis. J. Virol. 21:694-712. 19. Rhode, S. L. 1977.Replication process of the parvovirus
H-1. IX. Physical mappingstudies of the H-1 genome. J. Virol.22:446 458.
20. Rhode, S. L. 1978. Replication process of the parvovirus H-1. X.Isolation of a mutant defective in replicative-form DNAreplication. J. Virol. 25:215-223.
21. Rhode, S. L. 1978. Defectiveinterfering particles of parvo-virusH-1. J. Virol. 27:347-356.
22. Rhode, S. L. 1977. H-1 DNA synthesis, p.279-2%.In D. Wardand P. Tattersall(ed.),Replication of mammalian parvoviruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
23. Sanger, F., A. R.Coulson,B. G.Barrell, A. J. H. Smith, andB. A. Roe. 1980.Cloning in single-stranded bacterio-phage as anaidtorapidDNAsequencing. J.Mol. Biol. 143:161-178.
24. Sanger, F.,S. Ndcklen, and A. R.Coulson. 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad.Sci.U.S.A. 74:5463-5467.
25. Sekiya, T., M. Kobayashi, T.SekI, and K. Kolke. 1980. Nucleotide sequence of a clonedfragment ofrat mito-chondrial DNA containingthe replication origin. Gene 11:53-62.
26. Shiger,I.I., and S. L. Rhode. 1977.Replication process of theparvovirus H-1. VII. Electron microscopy of replica-tive form DNA synthesis. J. Virol. 21:713-723. 27. Singer,1.I., and S. L. Rhode.1977.Replicationprocessof
the parvovirusH-1. VIII. Partialdenaturation mapping andlocalization of the replicationoriginof H-1replicative formDNA with electron microscopy. J. Virol. 21:724-731.
28. Shiger, I. I., and S. L. Rhode. 1978.Ultrastructural studies
of H-1 parvovirus replication. VI. Simultaneous
autora-diographicand immunochemical intranuclear localization of viral DNA synthesis and protein accumulation. J. Virol. 25:349-360.
29. Smith, A.J.H.1980. DNA sequence analysisbyprimed synthesis. Methods Enzymol. 65:560-579.
30. Tal, J., P. Ron, P. Tattersall, S.Bratosin, and Y. Aloni. 1979. About 30%o of minute virus of mice RNA isspliced out followingpolyadenylation. Nature (London) 279:649-651.
31. Tattersall, P., and D. C. Ward. 1976. Rolling hairpin model for replication of parvovirus and linear chromo-somalDNA. Nature (London)263:106-109.
32. Tlinoco, I., Jr., P. N. Borer, B. Dengler, M. D. Levine, 0. C.Uhlenbeck,P. M. Crothers,andJ.Gralla. 1973. Im-proved estimation of secondary structure in ribonucleic acids.Nature(London) 246:4041.
33. Tseng,R.Y., R. H.Grafstrom,D.Revie, W. Oertel, and M.Goulian. 1979. Studies on early intermediates in the synthesisof DNA in animal cells. Cold SpringHarbor Symp. Quant. Biol. 43:263-270.
34. Usategul-Gomez, M., F. Al-Lami, S. M. Hopkins, N. Ledinko, and H. Toolan. 1969. Single-stranded DNA from theparvovirus H-1.Virology 39:617-621.
35. Ward, D. C., K. Dlnshaw,and D. K.Dadachanj. 1978. Replication of minute virus of mice DNA, p. 297-313. In D.Ward and P. Tattersall (ed.), Replication of mammalian parvoviruses. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
36. Ward, D. C., and P. Tattersall (ed.). 1978.Replicationof mammalian parvoviruses. Cold Spring Harbor Labora-tory,Cold Spring Harbor, N.Y.
37. Wolter, S., R. Rkhards, and R. W. Armentrout. 1980.Cell cycle-dependentreplication of the DNA ofminutevirusof mice,aparvovirus.Biochim. Biophys.Acta 607:420-431.
on November 10, 2019 by guest
http://jvi.asm.org/