0022-538X/81/070075-07$02.00/0
Amplification of
a
Short Nucleotide Sequence
in the
Repeat
Units of Defective
Herpes
Simplex
Virus Type
1
Angelotti
DNA
H. C.KAERNER,* A. OTT-HARTMANN, R. SCHATTEN,C. H.SCHRODER, ANDC. P. GRAY
Instituteof Virus Research, German Cancer Research Center, 6900 Heidelberg, West Germany
Received31December1980/Accepted 8 April 1981
Ithas been shown earlier thatthe reiterated regionsTRsandIRsbracketing the
Ussegment ofherpes simplex virustype 1Angelotti DNAareheterogeneous in
size by stepwise insertion of one to six copies of a 550-base-pair nucleotide
sequence. Considerably higher amplification of this sequence was observed in
defective viral DNA:up to 14copieswere detectedtobe inserted in therepeat
units ofamajor class of defective herpes simplex virus type 1Angelotti DNA,
dDNA1, which originated from noncontiguous sites locatedinUL and the inverted
repeats of the Scomponentofthe parentalgenome. Physicalmaps were
estab-lished for the cleavage sites ofKpnI,PstI,XhoI, and BamHI restriction
endonu-cleasesontherepeatsof dDNA1. Themapposition of the insertionsequence was
determined. Itwasdemonstrated that theamplified insertswere notdistributed
at random among or within the repeats. A given total population of dDNA1
molecules consistedof differenthomopolymers, eachof which containeda
con-stant number of inserts in all of its repeats. Assuming that a rolling-circle
mechanism is involved in thegeneration of full-length defective herpes simplex
virustype 1AngelottiDNAfrom singlerepeatunits,these data suggestthat the
550-base-pairsequence isamplifiedin therepeatsbefore thereplicationprocess.
Virionscontainingdefective viral DNAappear
in the virus offspring in the course of serial
passagesof herpesviruses athighmultiplicities
of infection(1-7,9,11-14, 17,18, 20). The
defec-tive DNAhas, approximately,aviralDNA unit
lengthandis madeupby repetition ofrestricted
portions of the parental genome. The present
study is concerned with one major classof
de-fective herpes simplex virus type 1 Angelotti
(HSV-1 ANG) DNA, dDNA1, which has the
samebuoyant densityasthe parental viralDNA
(17). dDNAl hasarepeat sequenceof
approxi-mately7x
10'
daltons andoriginates
fromnon-contiguous sites of the parentalgenomelocated
between0.33to0.42 and either0.82 to0.857 or
0.963 to 1.0 mapunitsontheprototypeisomer
(9). As has been reportedearlier, the reiterated
regionsTRsandIRsof theScomponentof
HSV-1ANG standardDNAcontainone tosixtandem
insertionsofa550-base-pairsequenceunit.
Con-sequently, theS-terminal and the L-Sjoint
re-striction fragments appear as series of DNA
bandsin agarosegels, equidistant byabout 0.35
x
10'
daltons. By investigating the sequencearrangement ofHSV-1 ANGdDNA1,wefound
that, in some of the repeats of this class of
defective DNA, this sequence is amplified as
muchas14-fold. One questionarisingfrom this
findingwaswhether the insertsweredistributed
atrandom within andamongtherepeatunits of
individual dDNAl molecules or whether the
totalpopulation of dDNAl molecules consisted
of differenthomopolymers madeupofidentical
repeatscontainingconstantnumbers ofinserts.
To distinguish between these possibilities, we
partiallydigested dDNAl withanumber of
re-striction endonucleases and examined the
di-gestsfor theappearance andsizedistribution of
intermediatefragments. Most of theenzymes we
usedrenderedsmearswhichcouldnotbe
inter-preted. PstI, however, cleavedpreferentiallyon
one of its two cleavage sites on the dDNA1
repeats. From the fragmentpatterns ofpartial
PstI digests we concluded that dDNAl repre-sents a mixture of different homopolymers. Physical mappingof thecleavagesites of various
restriction enzymes on the repeat sequence of
dDNAl further revealed that all of the
homo-polymers have common left-hand and right-handtermini.
MATERIALS AND METHODS
Virus andcells.HSV-1 ANG(1,13,15)was
prop-agatedonAfrican greenmonkeykidneycells(RC-37
Rita,Italdiagnostics, Rome, Italy)asdescribedearlier
(17).
75
on November 10, 2019 by guest
http://jvi.asm.org/
dDNA1. The development of virus particles
con-taining dDNA1 during serial virus passages at high
multiplicities of infection and the isolation of the
ECORI-, HpaI-, and HindIII-resistant dDNA1 from
maturevirions has been described in detailpreviously
(9, 17).In afinal step, defective DNAwaspurifiedas
DNAsedimentingatthesameapparentrateinneutral
sucrosegradientsasthatof viral standard DNA.
Restriction endonucleases. BamHIwas a
prod-uctof BRL,Inc.,Rockville,Md. All other restriction
endonucleases were purchased from New England
Biolabs, Boston, Mass. Enzyme digestionswere
per-formedby theprescription of theproducers.
Gelelectrophoresis of dDNA1 restriction
frag-ments.Electrophoresiswascarriedoutin0.6 or0.8%
vertical agarose (Seakem, Richmond, Calif.)slabgels
at 40V for18hat roomtemperature. Thegelswere
then stained with ethidium bromide andphotographed
under UVlight.
RESULTS
Evidence of multiple insertions of a
550-base pair nucleotide sequence dDNAl.
dDNA1 was digested with various restriction
enzymes, and the fragments were separated in
Xho
I
Pst I
agarose gels. Completion of the digestions was
carefully tested as follows: excess enzyme was multiply addedat 2-hintervals afterthe first
2-hdigestion period, and, in parallel assays, diges-tions were continued for 24 h as controls. The cleavage patterns obtained with XhoI, PstI, BamHI, and KpnI (Fig. 1) represented complete digestion in eachcase.
Allof the patternsdisplayed remarkable "step ladders" of DNA fragment bands, starting with base fragments of different molecular weights. The individual steps differedby 0.35 x 106
dal-tonsineachcase.Thisfinding suggests that the
repeat sequences of dDNAl contain multiple insertions of 550-base-pair sequences. The dif-ferent intensities of the individual step frag-ments further suggest that the number of dDNAl molecules present decreases with the increasing number of insertions that they con-tain.
Sequence or organization of the repeat
units of HSV-1 ANG dDNAl. In addition to
the majorstepladders, all of the fragment
pat-BamH
I
-A -B
5.71
5,35
5.0 * 4,77 *4.4 *4,01
-C
-E 1-t> 1.05
0.94
5,05 -4,7 -4
357
4,35----0 4,1
t *3,75
-A -o 3'4
t -.
3,05--B -o2,7
* 2,31 --f>2,08-.
1,6
-c -D
WV WV
'WV
t~~~~~~~~~1
&:'S:m:
-A 53
4.95
-4,6
-*4,05 *3,7
-C 2,65
--lo2,3 -D
1,95
41,6 6 _
1.4 _ - D
FIG. 1. DigestionofHSV1ANGdDNAIwiththe restriction endonucleases XhoI, PstI, BamHI, and KpnI.
Ethidium bromide-stained restrictionfragments electrophoretically separatedondifferentagarosegels (0.5 to 1%). The numbers represent the molecular weights x 106 of thefragments which were estimated by
calibratingthegelsusing HindIII restriction fragments ofADNA(19) and simian virus40DNA (15)and
HaeIIIrestrictionfragments of gX174RF DNA (10)asmarkers. Openarrows,left-handterminalfragments; Filledarrows, right-handterminalfragments.
5.25-4.9 4.55-*4.2 *3,85 *3.5
Xi>1.15
0.8
0.54
-A
-B
I
KnI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.58.448.307.588.2]REPEAT UNITS OF DEFECTIVE HSV-1 ANG DNA
ternsshown inFig. 1(andanumberofpattems
resulting from other restrictionenzymes which
are notshown) containeda second minorstep
ladder offragments (XhoI-B, PstI-B, BamHIB,
andKpnI-C)whichalsoareequidistantby 0.35
x
10'
daltons.Furthermore,each of thepatternscontainedoneminorfragment (XhoI-C, PstI-C,
BamHI-C, and KpnI-B) which when added
to-gether with the base stepof the corresponding
minor step ladder, equals the molecular weight
of the base step fragment of the major step
ladder. The molarities of the DNA fragment
bands were estimated from their molecular
weights andby optical scanning of the stained
gels. By taking the molarity of the discrete minor
fragment(indicated by theopen arrowinFig. 1)
for the individual pattern as one (i.e.,onlyone
fragment ispresent per dDNAl molecule), we
found the sum of the molarities of the minor
stepladderfragmentstobeapproximatelyone.
Itwasassumed that the fragments of the minor
stepladders and the 1 M fragmentsin eachof
theindividualpatternsshown inFig.1represent
left-hand and right-hand terminalfragments of
thecomplete dDNA1 molecules. This
assump-tion implies that the left-hand terminal
frag-Left Hand Terminus
mentscreatedby XhoI, PstI, BamHI,and KpnI
do not contain multiple 550-base-pair inserts,
whereas all of thecorresponding right-hand
ter-minal fragments do, and, hence, each of the
latter is displayed as a step ladder of bands in agarosegels.
The fragments XhoI-D, XhoI-E, PstI-D,
BamHI-D, and KpnI-D, all ofwhich were more
than10 M asjudgedbyopticalscanning, were
consideredto representinternalsegments of the
dDNA1 repeat sequenceunits. The same high
molarity was determined for the sum of the
major step ladder fragments (XhoI-A, PstI-A,
BamHI-A,andKpnI-A), which apparently also
representinternal DNA fragments.
From the above data, the sequence
arrange-mentof the dDNA1 repeatsand themap
posi-tions of the KpnI, XhoI, PstI, and BamHI
cleav-agesitesonthe repeatunitswere derived. The
resulting physicalmaps(Fig. 2) were confirmed
byaseries of double digestions of dDNAlwith
different restriction enzymes. Four
representa-tive cleavagepatternsobtained are shown in Fig.
3. Each of them agreed withthe corresponding
double cleavage pattern predicted from the
physicalmaps(Fig.2).
(1-14x) (nx)
RightHand Terminrus
(1-14x)
(nx) ;
(nx) i
0.0 01 0.2 0.3 0,4 0.5 0.6 07 0.8 0:s1,0
0
0.1 0.2 0;3 04 0.5 0;6 0;7 0.8 0,9 1.0
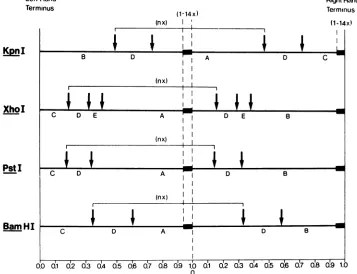
FIG. 2. Scalemapsof KpnI, XhoI, PstI,and BamHIcleavagesitesontherepeatsequenceofHSV-1ANG dDNA1. The blackbarsrepresentthe550-base-pairinserts. nequalsthe numberof repeatsperunit-length moleculeofdDNAIand variesfrom9to17dependingonthe copy numberofinsertsperrepeat.
I
XhoI
PstI
Bam HI
VOL. 39,1981 77
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.73.430.342.616.2]78 KAERNER ET AL.
C
0 - I(A
.c i a, m
s coE ye sU. 4
_ x (A
4 - cc.
a _ E E C
a- 0. c c U)
_ _
a. a.
0 _
o c
x IC
_CL
E v
m
_ E
E-a. c
I
3.3x106d
Bam HI B BamHi D Pst C
_ Pst D
!36X106d
Bam Hi C
t
Kpn C
0.95x1O6d
0.72X106d
0.63X1O6d
_. bCLc
cL vd
0
_C _
x cL
4.5X106d
t Xho B
Pst C _ Xho D
0.5X106d
I
2.5X106d
Kpn IC
Kpn D
Pst C
0.98X1O6d
Pst D
A B C D
FIG. 3. PstI-BamHI, BamHI-KpnI, XhoI-Pst,andPstI-Kpn I double digestions of HSV-1 ANG dDNA1.
Ethidiumbromide-stained restrictionfragments electrophoreticallyseparatedonagarosegels. The
nomen-clatureofthefragmentsisasdescribedfor Fig.1and2. The molecularweight calibrationwasperformed in
eachofthetwogels(AandB) by usingHindIII restrictionfragments ofX DNA(19), simian virus 40 DNA(15),
and HaeIII restriction fragments of4OX174RF DNA (10) as markers and is shown in gel A. For better
illustration thefragmentpatternsofthe PstI-BamHI and theBamHI-KpnI double digestswerecompared
with thefragmentpatternsof single digestswithPstI, BamHI, and KpnI in gel A. The fragment patterns of
thedoubledigestsin A and Bareshownschematicallyin C andD,respectively.
Forexample, the double digestion ofdDNAl
withPstI andBamHI created the left-hand
ter-minal fragment PstI-C, the fragments PstI-D
andBamHI-D,and the series ofright-hand
ter-minalfragmentsBamHI-B.Themajor step lad-der offragments in this digest had a base
frag-ment of 3.3 x 106 daltons, as expected, and
mappedbetween the BamHI cleavage site D-A
and the PstI cleavage site -A-D. BamHI-KpnI
digestion created the left-hand terminal
frag-ment BamHI-C and the series of right-hand
terminal fragments KpnI-C, together with the
otherfragments predicted fromthe maps shown
inFig.2.Similar conclusionscanbe drawn from
theXhoI-PstI andthePstI-KpnI cleavage
pat-terns.
Asshownin Fig.2, it isconceivable that the
550-base-pairsequenceinserts (1to 14copies [at
least]) cluster between the map positions of
about0.74and1.0;these map coordinatesfollow from the size of the left-hand terminal fragment
KpnI-B (3.1 x
10'
daltons) and the basefrag-ment of the right-hand terminal fragments
KpnI-C (1.6 x 10'daltons), assuming that the
latter containsatlast one of the inserts. In the mapsshown inFig. 2, the clusters of inserts are
tentatively placedatthe right-hand end of the
dDNAlmolecules, speculatingthat part of the insert sequence might function as a signal for
cutting the repeat concatemers to viral DNA
unitlength. Accordingtothe sequence
arrange-mentshowninFig. 2,all dDNA1 molecules had
m
n
m
L
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.54.449.52.403.2]REPEAT UNITS OF DEFECTIVE HSV-1 ANG DNA
identical left-hand and right-hand ends. The
total sequence complexity of thedDNAlrepeat
unitwas calculated fromthephysical maps to
be about 6 x 106daltons (KpnI pattern, 6.0 x
106; BamHI pattem, 5.95 x 106; PstI pattern,
5.94 x 106;XhoIpattern,5.9 x 106 daltons).
Nonrandom distributionof the
550-base-pair insertions in the dDNAl repeat
se-quence units. The modelof the sequence
ar-rangementindDNAl (Fig. 2)raisesthequestion
of whether individual dDNAl molecules are
made up at random of repeats with different
copy numbers of the insertion sequence or
whethereach of thecompletedDNA1 molecules
consists ofacertain number of identical repeats,
each of which has the samenumber of inserts.
As an experimental approach to answer this question, dDNA1 was partially digested witha
series of restriction endonucleases, andthe
ob-tained fragment patternswereanalyzedon
aga-rosegels.Mostofthe enzymes assayedforthis
purpose rendered smears of fragment bands.
However, PstI endonuclease created partial
digestion fragment patterns which allowed a
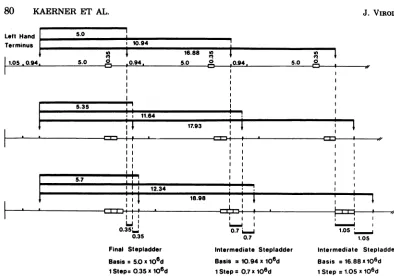
conclusive interpretation. Figure4shows a
se-quential series of"snapshots" representing
ini-tialstatesof the PstIdigestion. Beside the
cre-ation of the final majorstepladder (0.35x 106
daltons[Fig.1]),twointermediate fragmentstep
ladders withstepsof0.7 x106andof 1 x 106to
1.1 x
106
daltons can be detected. Thecorre-sponding base step fragments have molecular
weights of about11 x 106and 17 x 106,
respec-tively. Afurther group of bands which are not
clearly resolvedonthegelstartwith fragments
of about23x106daltons. Theinterpretationof
these results is shown in Fig. 5; the different
intermediate fragmentstep laddersapparently
represent theoriginationofdimers, trimers, and
ofmultimers ofahigherorderof therepeats. In
each of the multimers, the terminal0.94 x
106-dalton fragment PstI-D was cut off. The fact
that the step ladders of dimers and trimers,
which areclearlyresolved in the gelsshownin
Fig. 4,have stepsizes of0.7 x 106 (2 x 0.35 x
106) daltons and1x106(3x 0.35 x
10r)
daltons,respectively, stronglysuggests that the
550-base-pair inserts are not interspersed atrandom in therepeatsofindividualdDNAl molecules. It is
conceivable that, in the case of random
distri-bution, all intermediate step ladders would
dis-play acommonstep size of0.35 x 106 daltons.
Thus, onemustconcludethat eachofthesteps
ofanindividualstep ladderindicates the
exist-ence of a certain class of dDNA1 molecules,
made up by identical repeats, each of which
containsaconstantnumberof inserts.This
im-plicates that different dDNAl molecules are
madeupby different numbers of repeats which
23s00 11,00
5,53
4,4
t
4,0130 1,05 0,94
D C B A
FIG. 4. Partial digestion ofdDNA1 withPstI
re-striction endonuclease. Amounts (8 g) of dDNAI
weredigested with0.4Uof PstI (the definition of one
enzymeunitwasasgiven by the suppliers[BRLI).
Portionsof the digests containing 1.5 ug ofDNA were
takenfrom the reaction mixtureat 4(D),6(C), 8 (B),
and 12min (A) after thestartof the reaction and
wereanalyzedon anagarosegel. Thenumbers
rep-resent molecular weights x 106 of the fragments,
whichweredetermined by using HindIII fragments
ofXDNA, simian virus40 DNA, and HSV-1 ANG
viral standard DNAasmarkers. Filled arrows,
right-handterminalfragments(PstI-B,Fig.1);open arrow:
left-handterminalfragment (PstI-C,Fig.1).
areheterogeneousin size.Assuminganaverage
size of the complete molecules ofabout 100 x
106
daltons, therepeatnumbersmustvaryfrom17 (1 insertperrepeat, 6x 106daltons)to9 (14
insertsperrepeat, 10.6x
10'
daltons).Similardataon thecomposition ofrepetitive
defective HSV-1 (Justin) DNA has been
pro-videdbyFrenkeletal (5).Bypartial
denatura-tion studies, these authors could demonstrate
twotypes of DNAmoleculesbuilt up eitherby repeatunitswithoneadenine-plusthymine-rich
regionorbyrepeatunitswithtwoadenine-plus
thymine-rich regions.
79
VOL. 39,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.251.447.61.379.2]LeftHand Terminus
I.2 na
5.0
in c,
d
10.94
I l1e-a In I
41% o -( A 0n 0 I
IIIl
FII
i11.64a
i
1
17.93 iII
I
I I a r-Im we5.7
0 I3
0.35'
18.98
0.7 _.
0.7
1.05 ',_
1.05
Final Stepladder Intermediate Stepladder Intermediate Stepladder Basis:5.0x10d Basis 3 10.94 x 106d Basis =16.88x106d
[image:6.493.53.447.45.321.2]1Step:0.35x106d 1Step: 0.7x106d 1Step=1.05 x 106d
FIG. 5. Schematicalinterpretationofthepartialdigestion ofdDNAIwith PstI. The scheme shows three
differentdDNAI molecules with one, two, and three inserts per repeat. The insertsarerepresentedbyopen
bars.Preferentialcleavagesitesaremarkedbyarrows.The blackbars indicate thepreferentially originating
fragments.Thenumbersaremolecularweightsx106 DISCUSSION
A550-base-pairsequenceof theHSV-1ANG
genome wasfound earliertobeamplifiedtosix
copies in regions TRsand IRs of standard viral
DNA(9). Locker and Frenkel (11) andWagner
and Summers (21) have detected variable,
strain-specific sizeheterogeneitiesof the L ends
and the L-Sjoints of the DNA of HSV-1 strains
KOS, F, and Justin and have proposed that this
is duetotheamplificationofashortnucleotide
sequence. These authors, however, have
ana-lyzed viral DNAfrominfectedcells,whereas in
the study presented here, viralDNAextracted
frommaturevirionswasinvestigated. It should
be mentioned thatweobtainedthe sameresults
by using HSV ANG dDNAl isolated from
in-fectedcells.
Thedefective derivative HSV-1ANG dDNAl
is ofspecial interest as it originates from
non-contiguoussites of the parentalDNA.dDNA1,
besides beingpart ofTRs or IRS, including the
550-base-pair sequence, consists of sequences
from UL. Surprisingly, the 550-bapair
se-quence wasamplifiedtomuchhighercopy num-bers inthe dDNA1repeats than in the parental DNA. Thisfactisimportantinconnection with another result ofthe presentstudy which sug-gests that the totalpopulationofdDNAl
mole-cules consists of different homopolymers, each
of which is madeupof identical repeatswitha
constant copy number of the 550-base-pair
in-sertion sequence.Consideringresults of Locker
and Frenkel (12)whichsuggestthat individual
repeatunits of defective HSV-1 DNA are
elon-gatedtofull-size defective viralDNAbya
roll-ing-circlemechanism, it isconceivable that the
amplification of the insertion sequence in the
dDNAl repeatoccursbefore theonsetof
repli-cation.
Roizman (16) and Jacob etal. (8) have
pre-sented a hypothesis of the replication ofHSV
DNAviaarolling-circlemodeland the
genera-tion of all four isomers of HSV DNA from one
isomer. Onecentralfeature of this hypothesis is
the regeneration of the "a" sequence, which is
present at both ends of the molecule and is
reiterated in the L-Sjointin opposite
orienta-tion. Circularization of the DNA molecules by
ligationof cohesive ends would leadto theloss
ofonecopy of the "a" sequence whichhas to be
regeneratedto achievecompleteviralDNA
mol-ecules either before circularizationoraftertheir
excision from concatemeric DNA. The above
authorsproposedin their modelthatthe
regen-eration of the "a" sequence would occur after
replication via a rolling-circle mechanism and
that inthecourseof theregeneration of the "a"
I .--- I
-,-I ..O%db I T. I
c.n
I i I
on November 10, 2019 by guest
http://jvi.asm.org/
VO9%91REPEAT UNITS OF DEFECTIVE HSV-1 ANG DNA
sequence,theother isomers could beformned.
One conceivablespeculationfrom thepresent
datais that the550-base-pairsequencein
HSV-1 ANG standard DNA andindDNAl could be
(or could contain) the "a" sequence. The
ob-servedamplification of thissequence instandard
anddefective HSV-1 ANG DNA could then
re-flect one step of the viral DNA replication in
terms of the above-mentioned hypothesis,
which, forsomereason, doesnotstopafter the
regeneration of one copy of the "a" sequence.
Thefact that the550-base-pairsequenceis
am-plified to a higher degree in the repeats of
dDNA1 than in theparental DNAsuggeststhat
aplificationcanoccurin standard DNAaswell
asin the dDNAl repeatsthemselves, i.e., after
their formation from UL and the repeats from
the S component. On the other hand, higher
copy numbers of the amplified sequence are
evidently ofnoadvantage for the replication of
the repeats, as can be concluded from the
in-creasing number of insertions which correlate withadecreasing number of dDNA1molecules (Fig. 1).
Molecular cloning of dDNA1 restriction
en-zyme fragments and DNA sequencingare now
in progress to elucidate the sequence
arrange-mentofdDNA1. It should be mentionedfinally
that another type of defective HSV-1 ANG
DNA, dDNA2 (9) which closely resembles the
HD DNAderived from HSV-1 Justin by Frenkel
and co-workers (5, 6, 11), also shows multiple
insertions of the550-base-pairunit described in
thispaper(Kaerner,unpublisheddata).
LITERATURE CITED
1. Ben-Porat,T., J. M. Demarchi, and A.S. Kaplan. 1974.Characterization of defectiveinterfering viral par-ticles present in apopulationofpseudorabiesvirions. Virology60:29-37.
2. Bronson,D.L.,G. R.Dreesman,N.Biswal,and M.
Benyesh-Melnick. 1973.Defective virions ofherpes
simplexviruaes. Intervirology 1:141-153.
3. Campbell,D. E.,M. C. Kemp,M. LPerdue, C. C.
Randall,and G.A.Gentry.1976.Equine herpesvirus
in vivo:cyclicproductionofaDNAdensityvariant with repetitive sequences.Virology60:737-750.
4. Fleckenstein,B., G.W.Bornkamm,and H.Ludwig.
1975.Repetitivesequences incompleteand defective genomes ofHerpesvirussaimiri.J.Virol.15:398-406. 5. Frenkel,N.,R.J.Jacob,R. W.Honess,G. S.
Hay-ward,H.Locker,and B.Roizman.1975.Anatomyof herpessimplexvirus DNA.III. Characterization of
de-fective DNA molecules and biological properties of
virus populations containing them. J. Virol.16:153-167. 6. Frenkel, N., H. Locker, W. Batterson, G. S. Hayward, and B. Roizman. 1976. Anatomy of herpes simplex virus DNA. VI. Defective DNAoriginatesfrom the S component. J. Virol. 20:527-531.
7.Graham,B.J., Z.Bengali,and G. F. Vande Woude. 1978. Physical map of the origin of defective DNA in herpes simplex virus type 1 DNA. J.Virol. 25:878-887. 8. Jacob, R. J., L S. Morse, and B. Roizman. 1979. Anatomyof herpessimplex virus DNA. XII. Accumu-lation ofhead-to-tail concatemers in nuclei of infected cells and their role in the generationof the four isomeric arrangements of viral DNA. J. Virol.29:448-457. 9.Kaerner, H. C., L. B.Maichle, and C. H. Schrdder.
1979.Originof two different classes ofdefectiveHSV-1 AngelottiDNA.Nucleic Acids Res. 6:1467-1478. 10.Lee, A.S.,and R. LSinsheimer.1974. Acleavage map
ofbacteriophageOX174genome.Proc. Natl. Acad. Sci. U.S.A.71:2882-2886.
11. Locker,H., and N.Frenkel.1979.Structureand origin ofdefective genomes contained in serially passaged herpessimplexvirustype1(Justin).J.Virol. 29:1065-1077.
12.Locker,H., and N.Frenkel.1979.BamI,KpnI,andSall restriction enzyme maps of the DNAs ofherpes simplex virus strains Justin andF: occurrence of heterogeneities indefined regions of the viral DNA. J. Virol. 32:429-441.
13. Murray,B.K., N.Biawal,J. B.Bookout,R. E. Lan-ford,R. J.Courtney,and J.L. Melnick.1975.Cyclic appearanceof defectiveinterfering particlesofherpes simplex virus and the concomitant accumulation of early polypeptideVP175.Intervirology5:173-184. 14.Ott,A.,B.Fohring,and H.C. Kaerner. 1979.Herpes
simplex virus type 1 Angelotti and a defectiveviral genotype: analysis of genome structures andgenetic relatedness by DNA-DNA reassociation kinetics. J. Virol.29:423-430.
15. Reddy,V.B.,B.Thimmappaya,R.Dhar,K. N. Sub-ramanian,B.S.Zain,J.Pan, P.KL Ghosh,M.L. Celma,and S. M. Weissman. 1978.The genome of Simian virus40.Science 200:494-502.
16. Roizman, B.1979. The structure and isomerization of herpes simplexvirus genomes. Cell16:481-494. 17. Schrdder,C.H.,B.Stegmann, H. F. Lauppe, and H.
C. Kaerner. 1975/76. An unusual defective genotype derivedfromherpes simplex virus strain ANG. Inter-virology6:270-284.
18. Stegman, B., H. Zentgraf, A. Ott, and C. H. Schr6der. 1978. Synthesis and packaging ofherpes simplexvirus DNA in thecourseof viruspassagesat highmultiplicity.Intervirology10:228-240.
19. Thomas, M., andR. W. Davis. 1975. Studiesonthe cleavage ofbacteriophage lambda DNA with EcoRI restriction endonuclease. J. Mol. Biol. 91:315-328.
20.Wagner, M., J. Skare, and W. C. Summers. 1974. Analysis of defectiveherpes simplex virus type 1by restrictionendonucleasecleavageandnucleic acid hy-bridization. ColdSpringHarborSymp. Quant.Biol. 39:
683-686.
21. Wagner,M.J.,and W.C. Summers. 1978. Structure of thejointregionand the terminiof the DNA ofherpes simplexvirustype1.J.Virol. 27:374-387.
81 39,1981
on November 10, 2019 by guest
http://jvi.asm.org/