The prototype foamy virus (PFV) glycoprotein, which is essential for PFV particle release, displays a highly unusual biosynthesis, resulting in posttranslational cleavage of the precursor protein into three particle-associated subunits, i.e., leader peptide (LP), surface (SU), and transmembrane (TM). Glycosidase digestion of metabolically labeled PFV particles revealed the presence of N-linked carbohydrates on all subunits. The differential sensitivity to specific glycosidases indicated that all oligosaccharides on LP and TM are of the high-mannose or hybrid type, whereas most of those attached to SU, which contribute to about 50% of its molecular weight, are of the complex type. Individual inactivation of all 15 potential N-glycosylation sites in PFV Env demonstrated that 14 are used, i.e., 1 out of 2 in LP, 10 in SU, and 3 in TM. Analysis of the individual altered glycoproteins revealed defects in intracellular processing, support of particle release, and infectivity for three mutants, having the evolutionarily conserved glycosylation sites N8 in SU or N13 and N15 in the cysteine-rich central “sheets-and-loops” region of TM inactivated. Examination of alternative mutants with mutations affecting glycosylation or surrounding sequences at these sites indicated that inhibition of glyco-sylation at N8 and N13 most likely is responsible for the observed replication defects, whereas for N15 surrounding sequences seem to contribute to a temperature-sensitive phenotype. Taken together these data demonstrate that PFV Env and in particular the SU subunit are heavily N glycosylated and suggest that although most carbohydrates are dispensable individually, some evolutionarily conserved sites are important for normal Env function of FV isolates from different species.
The envelope glycoprotein (Env) of retroviruses is synthe-sized as a precursor molecule in infected cells (reviewed in reference 29). It assembles into oligomeric complexes in the rough endoplasmic reticulum (RER), is extensively modified, and then is cleaved by a cell-encoded protease into surface (SU) and transmembrane (TM) subunits during transport to the cell surface before being incorporated into the budding retroviral particle. Some modifications, such as addition of N-linked sugars, start while the nascent protein is still cotrans-lationally translocated across the membrane and into the lu-men of the RER. The sites of covalent attachlu-ment are aspar-agine (N) residues within the canonical sequence for N-linked carbohydrates, N-X-S/T, where X is any amino acid except proline. During the process of oligomerization of the Env proteins and their intracellular transport to the cell surface, the carbohydrate chains are heavily modified by removal and ad-dition of specific sugar residues. In general there are three types of N-linked carbohydrate chains, i.e., high-mannose, hy-brid, and complex, which can be distinguished experimentally by digestion with specific glycosidases. The number and posi-tion of N-glycosylaposi-tion sites vary widely among retroviruses.
For example, the Env protein of human immunodeficiency virus (HIV) type 1 has approximately 30 glycosylation sites in the SU and TM subunits (6, 13), whereas the Env protein of murine leukemia virus (MuLV) harbors only 7 to 8 in SU and none in TM (5, 12). At least some of the oligosaccharides must have important roles in the proper folding of Env, because treatment of cells with the glycosidase inhibitor tunicamycin results in unglycosylated Env molecules that are trapped in the ER. However, in some cases (e.g., MuLV and HIV) individual sites and in a few cases (e.g., MuLV and simian immunodefi-ciency virus) multiple sites of N-linked carbohydrate chains can be eliminated by mutagenesis without affecting the transport of the Env protein (5, 12), whereas in other cases (e.g., HIV and MuLV), mutagenesis of specific sites results in a block only after transport to the RER or Golgi (5–7, 12, 26). In addition to their role in protein folding, carbohydrates of retroviral glycoproteins have also been implicated in other functions such as masking of immunodominant epitopes (e.g., HIV and sim-ian immunodeficiency virus) or regulation of coreceptor usage (e.g., HIV) (23, 24, 26, 28).
Recently retroviruses have been regrouped into two subfam-ilies, theOrthoretrovirinaeandSpumaretrovirinae. As an expres-sion of their unique replication strategy, foamy viruses (FVs) constitute the only genus of the subfamilySpumaretrovirinae
(reviewed in reference 27). The particle-associated glycopro-tein of FV is unique compared to other retroviral envelope proteins because its coexpression is strictly required for the FV particle release process and its function cannot be replaced by heterologous viral glycoproteins (reviewed in reference 18).
* Corresponding author. Mailing address: Institut fu¨r Virologie, Medizinische Fakulta¨t “Carl Gustav Carus,” Technische Universita¨t Dresden, Fetscherstr. 74, 01307 Dresden, Germany. Phone: 49-351-458-6210. Fax: 49-351-458-6314. E-mail: [email protected] -dresden.de.
† Present address: Rudolf-Virchow-Zentrum fu¨r Experimentelle Biomedizin, Universita¨t Wu¨rzburg, 97078 Wu¨rzburg, Germany.
7664
on November 8, 2019 by guest
The FV envelope precursor protein has a highly unusual bio-synthesis for a retroviral glycoprotein. It is translated as a full-length precursor protein into the RER and initially has a type III protein configuration, with both its N and C termini located intracytoplasmically (10, 19). Only during its transport to the cell surface is it posttranslationally processed by cellular, most likely furin-like proteases, and not the signal peptidase complex, into at least three subunits (4, 9). The N-terminal signal or leader peptide (LP) has a type II conformation, whereas the C-terminal TM subunit has a type I conformation. The internal SU subunit presumably associates with extracel-lular domains of TM on the luminal side (19, 32). Image reconstruction analysis from cryoelectron microscopy pictures of the characteristic prominent Env spike structures on FV particles indicates that the heterotrimeric Env protein com-plexes form trimers, similar to those reported for other viral glycoproteins (31). For the FV budding process at least two essential interactions between Env and Gag are required (19, 21). One of these is the contact of the N-terminal cytoplasmic region of the FV Env LP, the so-called budding domain, con-taining an essential conserved WXXW sequence motif, with the N terminus of the FV Gag protein (19, 32). The LP of FV Env is glycosylated, and cleavage products are viral particle associated (19). In the present study we intended to determine the extent of prototype FV (PFV) Env N glycosylation, and we analyzed the requirement of individual N glycosylation sites for PFV particle release and infectivity.
MATERIALS AND METHODS
Cells.The human kidney cell line 293T (3) and the human fibrosarcoma cell line HT1080 (25) were cultivated in Dulbecco’s modified Eagle’s medium sup-plemented with 10% fetal calf serum and antibiotics.
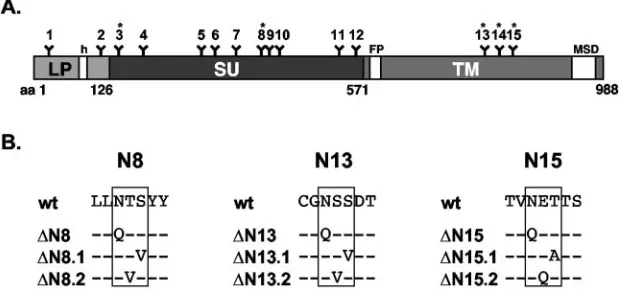
Expression constructs. The PFV retroviral vector pMH118 expressing an enhanced green fluorescent protein (EGFP) from an internal spleen focus-forming virus U3 promoter has been described previously (14). The pMH120 vector is a variant thereof having the EGFP marker gene replaced by the LacZ gene. The expression constructs of the individual PFV envelope mutants with
mutations affecting the first three potential N-glycosylation sites (⌬N1, pczHFV
envEM058 [N25Q];⌬N2, pczHFVenvEM077 [N109Q]; and⌬N3, pczHFVenv
EM078 [N141Q]) and of the SU/TM cleavage site mutant (pczHFVenvEM020
[R571T]) were described earlier (19, 22). All further N-glycosylation site mutants
were generated by recombinant PCR techniques (11) and are based on the
prototype foamy virus gp130Env
expression construct pczHFVenvEM002 de-scribed earlier (21). All PCR-derived fragments were sequenced to confirm the desired mutations and exclude further off-site mutations. Details on the cloning procedures for the individual mutants are available on request. The following
new single-site mutants were generated:⌬N4, pczHFVenvEM105 (N183Q);⌬N5,
pczHFVenvEM106 (N286Q);⌬N6, pczHFVenvEM107 (N311Q);⌬N7, pczHFV
envEM108 (N346Q); ⌬N8, pczHFVenvEM109 (N391Q); ⌬N8.1,
pczHFVen-vEM131 (S393V);⌬N8.2, pczHFVenvEM151 (T392V);⌬N9, pczHFVenvEM110
(N405Q); ⌬N10, pczHFVenvEM111 (N423Q); ⌬N11, pczHFVenvEM112
(N527Q); ⌬N12, pczHFVenvEM113 (N556Q); ⌬N13, pczHFVenvEM114
(N782Q); ⌬N13.1, pczHFVenvEM132 (S784V); ⌬N13.2, pczHFVenvEM152
(S783V); ⌬N14, pczHFVenvEM115 (N808Q); ⌬N15, pczHFVenvEM116
(N833Q);⌬N15.1, pczHFVenvEM133 (T835A); and⌬N15.2, pczHFVenvEM153
(E834Q). The positions of the potential N-glycosylation sites in the subdomains
of the PFV Env protein are shown schematically in Fig. 1.
Biochemical characterization of PFV Env proteins.For biochemical analysis of the PFV Env mutants, the individual expression constructs were cotransfected together with FV vector pMH118 into 293T cells by the calcium phosphate or polyethylene imine method essentially as described previously (19). Western blot analysis was performed as described previously (19). Polyclonal antisera used were specific for PFV Gag (1), the LP domain of PFV Env (amino acids [aa] 1 to 86) (19), or the LP and SU domains of PFV Env (aa 1 to 571) (20). Further-more, a hybridoma supernatant (clone P3E10) specific for the SU subunit of PFV Env was employed in some experiments (4). Metabolic labeling of trans-fected cells and viral particles secreted into the supernatant, particle purification, and immunoprecipitation with FV protein-specific antisera were done as de-scribed previously (19). For glycosidase treatment of viral proteins, immune complexes were eluted from protein A-Sepharose pellets after the final washing
step by boiling in 60l 1⫻denaturation buffer (0.5% sodium dodecyl sulfate
[SDS], 1%-mercaptoethanol) for 10 min at 98°C. Subsequently, the
superna-tant was split in three equal parts and transferred to fresh tubes, and 4l 10⫻
G5 buffer (0.5 M sodium citrate, pH 5.5) or 4l 10⫻G7 buffer (0.5 M sodium
phosphate, pH 7.5) and 4l 10% NP-40 were added for endoglycosidase H
(endo H) orN-glycosidase F (PNGase F) digestion, respectively. Following
volume adjustment to 40l with H2O and addition of 250 units endo H or
PNGase F (New England Biolabs), the tubes and corresponding mock-treated controls lacking the enzymes were incubated for 4 to 6 h at 37°C. Cell surface biotinylation of 293T cells transiently transfected with the individual Env expres-sion constructs was carried out essentially as described previously (22). Briefly,
293T cells were transiently transfected and metabolically labeled with [35
S]me-thionine and [35S]cysteine. At 36 h after the addition of the DNA, cell surface
[image:2.585.138.450.73.218.2]protein was labeled with NHS-biotin (Calbiochem) at 1 mg/ml in phosphate-buffered saline for 30 min. Subsequently, the biotinylation reaction was stopped by adding phosphate-buffered saline containing 100 mM glycine prior to cell lysis in radioimmunoprecipitation assay buffer. Lysates were precipitated with an FV-positive chimpanzee serum as described earlier (8, 20), separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE), and blotted onto nitrocellulose
FIG. 1. Schematic outline of the PFV glycoprotein domain organization. A) Schematic outline of the PFV Env domain organization. B) Alignment of wild-type and mutant protein sequences around potential glycosylation sites 8, 13, and 15. Amino acids altered in the mutant constructs are shown, and the corresponding construct names are given on the left. FP, fusion peptide; MSD, membrane-spanning domain; h, hydrophobic domain of the leader peptide. Potential N-linked carbohydrate chain attachment sites are indicated by Y and numbered 1 to 15. Asterisks mark evolutionarily conserved sites.
on November 8, 2019 by guest
http://jvi.asm.org/
48 to 72 h after infection. All transduction experiments were performed at least three times, and in each independent experiment the values obtained with wild-type (wt) PFV Env were arbitrarily set to 100. Alternatively, LacZ-expressing viral supernatants were titrated by infection of target cells with 10-fold serial
dilutions in 12-well plates and subsequent histochemical-galactosidase staining
48 h postinfection.
Analysis of temperature sensitivity.For assaying the potential temperature sensitivity of specific mutants, virus was produced in parallel at either 37°C or 30°C, by shifting one plate per construct of duplicate transfections to 30°C during the sodium butyrate induction step and leaving it at this temperature until virus was harvested 24 h later. Subsequently, the HT1080 target cells were incubated with viral supernatant or dilutions thereof for 6 h at 30°C. After removal of the viral supernatant, the target cells were incubated for an additional 16 h at 30°C before they were shifted to 37°C and assayed by fluorescence-activated cell sorting 48 h later.
RESULTS
Analysis of N glycosylation of cell- and particle-associated FV Env proteins by glycosidase digestion.FV particles contain three different processed glycoprotein subunits, LP, SU, and TM (4, 19). A schematic outline of the PFV gp130Env
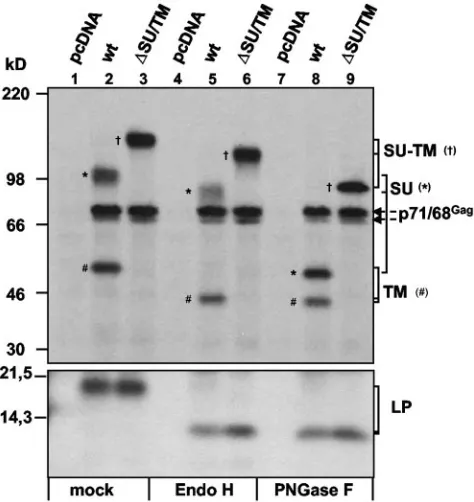
precur-sor organization and the locations of putative N-glycosylation sites within the individual subunits are depicted in Fig. 1. First we determined which of these subunits contain N-linked car-bohydrate chains. Therefore, PFV particles were generated by cotransfection of 293T cells with the PFV Gag/Pol-expressing vector pMH118 and the wild-type PFV Env protein expression construct pczHFVenvEM002 or the PFV Env SU/TM cleavage mutant expression construct pczHFVenvEM020 and subse-quently metabolically labeled. Following immunoprecipitation of cell- and PFV particle-associated viral proteins by using a mixture of anti-PFV Env and Gag antisera, the immune com-plexes were digested with endo H or PNGase F or mock incubated. In Fig. 2 the result of an SDS-PAGE analysis of such samples is shown. All three viral particle-associated PFV Env subunits of the wild-type protein contain N-linked carbo-hydrates, as their mobility increased after PNGase F treatment (Fig. 2, lane 8) compared to a mock-treated control (Fig. 2, lane 2). Similarly, both Env subunits of a SU/TM cleavage mutant (⌬SU/TM), LP and SU-TM, showed a decrease in molecular weight upon PNGase F treatment (Fig. 2, lane 9). Interestingly, a similar mobility shift of the LP and TM sub-units of the wild-type protein was seen in endo H (Fig. 2, lane 5)- and PNGase F (Fig. 2, lane 8)-treated samples. In contrast, the SU subunit showed only a small mobility shift after endo H treatment (Fig. 2, lane 5) but much larger shift after PNGase F treatment (Fig. 2, lane 8). Thus, this analysis indicated that all viral particle-associated PFV Env subunits are N
glycosy-lated. Furthermore, the differential sensitivity of the individual subunits to endo H and PNGase F suggests that that the N-linked carbohydrate chains of the LP and TM subunits are of the high-mannose or hybrid type, whereas at least some if not most of the carbohydrate chains of the SU subunit are of the complex type.
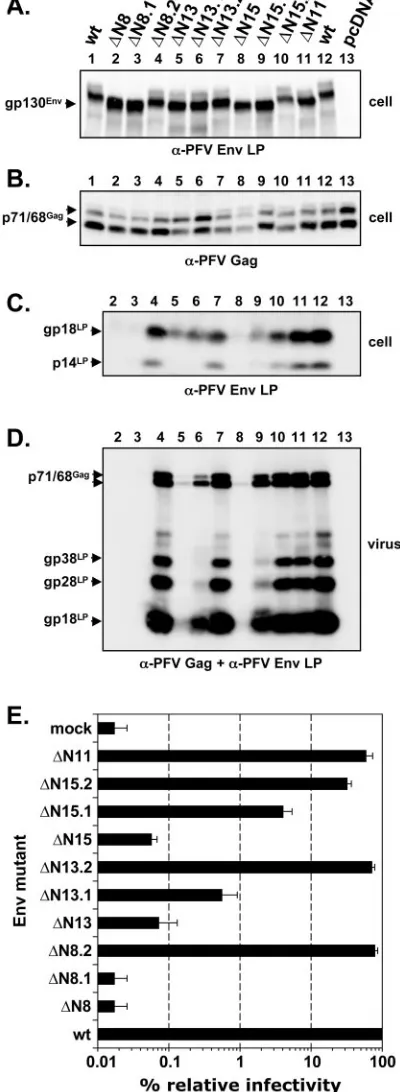
[image:3.585.303.540.70.321.2]Expression analysis of mutant PFV Env proteins with indi-vidually inactivated potential N-glycosylation sites.The results presented above demonstrated that all PFV Env subunits are N glycosylated. To identify at which of the 15 potential N-linked glycosylation sites (N1 to N15) found in the PFV Env protein sequence carbohydrate chains are indeed added, 15 PFV Env mutants, termed⌬N1 to⌬N15, were generated, in which one individual potential glycosylation site at a time was inactivated by changing the N-X-S/T consensus sequence to Q-X-S/T. After cotransfection of the different single mutants together with pMH118 into 293T cells, samples of cell lysates were separated by SDS-PAGE and analyzed by Western blot-ting using a PFV Gag (Fig. 3A)- or PFV Env LP (Fig. 3B)-specific antiserum or a PFV SU-3B)-specific monoclonal antibody (Fig. 3C). All mutants were expressed at similar levels intra-cellularly (Fig. 3B). Separation of the precursor protein
FIG. 2. Glycosidase digestion of particle-associated FV proteins. FV particles were generated by cotransfection of 293T cells with pMH118 and the indicated Env expression constructs. Thirty-two hours posttransfection, cells were metabolically labeled with [35
S]me-thionine and [35S]cysteine for 16 h. Subsequently, viral particles were
purified by ultracentrifugation through 20% sucrose and digested with endo H or PNGase F as indicated or mock incubated before separation of viral proteins by SDS-PAGE and exposure to X-ray film. The indi-vidual PFV proteins are indicated on the right, and the assignment of the specific bands to individual subunits is indicated by different sym-bols: SU (*), TM (#), and SU-TM (†). Cells were cotransfected with pMH118 and the following: lanes 1, 4, and 7, pcDNA3.1⫹zeo (pcDNA); lanes 2, 5, and 8, pczHFVenvEM002 (wt); lanes 3, 6, and 9, pczHFVenvEM020 (⌬SU/TM).
on November 8, 2019 by guest
gp130Env by low-percentage SDS-PAGE revealed small but
detectable mobility shifts for mutants⌬N2 to⌬N15 (Fig. 3B, upper panel, lanes 2 to 15) compared to the wild-type protein (Fig. 3B, upper panel, lane 16). This indicated that carbohy-drate chains are added at all potential N-glycosylation sites of the PFV Env protein except the first and confirmed our pre-vious glycosylation analysis of the PFV Env leader peptide gp18LP(19). Furthermore, a mobility shift of the LP cleavage
products was observed only for the ⌬N2 mutant (Fig. 3B, middle panel, lane 2), although both⌬N2 and⌬N3 showed a mobility shift of the gp130Envprecursor protein (Fig. 3B, upper
panel, lanes 2 and 3). This implies that LP cleavage occurs between PFV Env aa 109 and 141, which we recently confirmed by N-terminal protein sequencing of PFV Env subunits (4). In
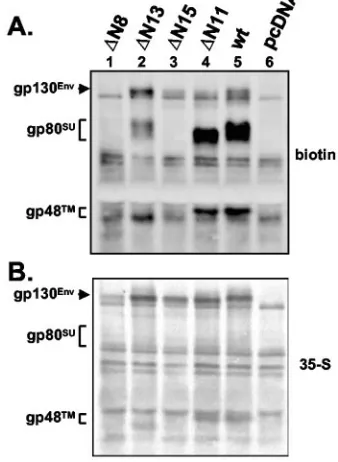
addition, the Western blot analysis of cell lysates revealed that the individual glycosylation mutants showed a differential pro-cessing of the LP. Mutants⌬N13 and⌬N14 (Fig. 3B, middle panel, lanes 13 and 14) displayed a moderately reduced pro-cessing and mutant ⌬N15 (Fig. 3B, middle panel, lane 15) displayed a strongly reduced processing, whereas no processing at all could be detected for⌬N8 (Fig. 3B, middle panel, lane 8). Cell surface biotinylation analysis of selected mutants revealed intracellular transport defects for mutants ⌬N8, ⌬N13, and
⌬N15 (Fig. 4). For wild-type PFV Env (Fig. 4A, lane 5) and the control glycosylation mutant⌬N11 (Fig. 4A, lane 4) significant amounts of gp80SU and gp48TM glycoprotein subunits and
small amounts of gp130Envprecursor were detected at the cell
surface. Mutant⌬N8 displayed the strongest transport defect, since no Env precursor or processed subunits could be de-tected at the cell surface (Fig. 4A, lane 1), although total protein expression was only slightly reduced compared to the other samples (Fig. 4B, lane 1). For mutant⌬N15 only small amounts of unprocessed gp130Envprecursor were detectable at
[image:4.585.46.283.72.382.2]the cell surface (Fig. 4A, lane 3), whereas mutant ⌬N13 showed cell surface expression of both precursor and pro-cessed subunits (Fig. 4A, lane 2); however, the relative amount
[image:4.585.336.505.77.308.2]FIG. 3. Biochemical analysis of PFV Env N-glycosylation mutants. Western blot analysis of 293T cell (cell) and purified PFV particle (virus) lysates using (A) anti-PFV Gag, (B) anti-PFV Env LP (aa 1 to 86)-specific polyclonal rabbit antiserum, or (C) anti-PFV-SU (P3E10) monoclonal hybridoma supernatant is shown. The individual PFV pro-teins are indicated. 293T cells were cotransfected with the PFV Gag/ Pol-expressing, replication-defective retroviral vector pMH118 and the respective PFV Env expression construct as indicated: lane 1, pcz HFVenvEM058 (⌬N1); lane 2, pczHFVenvEM077 (⌬N2); lane 3, pcz HFVenvEM078 (⌬N3); lane 4, pczHFVenvEM105 (⌬N4); lane 5, pczHFV envEM106 (⌬N5); lane 6, pczHFVenvEM107 (⌬N6); lane 7, pczHFV envEM108 (⌬N7); lane 8, pczHFVenvEM109 (⌬N8); lane 9, pczHFV-envEM110 (⌬N9); lane 10, pczHFVenvEM111 (⌬N10); lane 11, pc-zHFVenvEM112 (⌬N11); lane 12, pczHFVenvEM113 (⌬N12); lane 13, pczHFVenvEM114 (⌬N13); lane 14, pczHFVenvEM115 (⌬N14); lane 15, pczHFVenvEM116 (⌬N15); lane 16, pczHFVenvEM002 (wt); lane 17, pcDNA3.1⫹zeo (pcDNA). For lane 18, transfection was with only empty expression vector.
FIG. 4. Cell surface expression of selected PFV Env N-glycosyla-tion mutants. Transfected 293T cells were metabolically labeled, and subsequently cell surface proteins were specifically biotinylated. (A) Following immunoprecipitation with an FV-positive monkey se-rum, SDS-polyacrylamide gel electrophoresis, and blotting to nitrocel-lulose membranes, biotinylated PFV Env proteins were chemilumines-cently detected by incubation with streptavidin-horseradish peroxidase, ECL-Plus. (B) Subsequently, total cellular Env expression was visualized by PhophorImager analysis after the chemilumines-cence signal was allowed to fade overnight. 293T cells were cotrans-fected with the PFV Gag/Pol-expressing, replication-defective retrovi-ral vector pMH118 and the respective PFV Env expression construct as indicated: lane 1, pczHFVenvEM109 (⌬N8); lane 2, pczHFVenv EM114 (⌬N13); lane 3, pczHFVenvEM116 (⌬N15); lane 4, pczHFV envEM112 (⌬N11); lane 5, pczHFVenvEM002 (wt); lane 6, pcDNA3.1⫹zeo (pcDNA).
on November 8, 2019 by guest
http://jvi.asm.org/
rified by ultracentrifugation from supernatants of 293T cells cotransfected with the FV vector pMH118 and the individual Env expression constructs. The protein composition analysis of purified PFV particles by Western blotting using antisera spe-cific for PFV Gag, PFV Env LP, and Env SU subunits is shown in Fig. 3. No particle-associated Gag or Env proteins were detected for mutant⌬N8 (Fig. 3A to C, lane 8). In particle preparation of cells cotransfected with mutant⌬N13 or⌬N15, only small amounts of particle-associated Gag and Env pro-teins were detected (Fig. 3 A to C, lanes 13 and 15). Mutants
⌬N2,⌬N12, and ⌬N14 (Fig. 3A to C, lanes 2, 12, and 14) displayed a somewhat reduced physical particle release, whereas all other mutants revealed no obvious phenotype and showed a particle release comparable to that of wild-type PFV Env (Fig. 3A to C, lanes 1, 3 to 7, and 9 to 11). Furthermore, in the Western blot analysis a mobility shift in FV particle-associated LP cleavage products was observed only for the
⌬N2 mutant (Fig. 3B, lower panel, lane 2), whereas a shift in the SU subunit was detectable for mutants⌬N3 to⌬N7 (Fig. 3C, lanes 3 to 7) and⌬N9 to⌬N12 (Fig. 3C, lanes 9 to 12). The higher-molecular-weight LP cleavage products gp28LP and
gp38LP(Fig. 3B, lower panel) that we observed previously are
generated not by alternative proteolytic processing or differ-ential glycosylation but by additional posttranslational modifi-cation of the gp18LP cleavage product (4; N. Stanke and D.
Lindemann, unpublished results).
Taken together, these data demonstrate that the PFV LP subunit is N glycosylated at a single site at N109; that the SU
subunit is N glycosylated at 10 sites at N141, N183, N286, N311,
N346, N391, N405, N423, N527and N556; and that the TM subunit
is N glycosylated at 3 sites at N782, N808, and N833.
Further-more, they show that individual inactivation of glycosylation at a single site, N391(N8), in SU and at all three sites, N782(N13),
N808(N14), and N833(N15), in TM results in the failure of
these proteins to efficiently support PFV particle release.
Infectivity of PFV Env N-glycosylation mutants correlates with physical particle release.To determine the infectivity of PFV particles containing the mutant glycoproteins, superna-tants of the 293T cells cotransfected with pMH118 described above were further analyzed in an EGFP marker gene transfer assay on HT1080 cells. The results of this analysis are summa-rized in Fig. 5A. All mutants except three showed infectivities within a threefold range of that of wild-type PFV Env (Fig. 5A). Mutants⌬N13 and ⌬N15 showed 65-fold and 400-fold reductions in infectivity, respectively, whereas no infectivity could be detected for mutant⌬N8. To analyze the infectivities
of selected mutants in more detail, viral particle preparations of mutants ⌬N8, ⌬N11, ⌬N13, and ⌬N15 were titrated on HT1080 cells by using a more sensitive LacZ marker gene transfer assay (Fig. 5B). Supernatants of mutant⌬N8 showed an infectivity that was at least 4⫻104-fold lower than that of
wild-type FV Env-containing samples, whereas the infectivities of mutants ⌬N13 and ⌬N15 were reduced 600- to 700-fold. Similar to what was observed in the EGFP transfer assay, mutant⌬N11 showed an infectivity comparable to that of the wild type.
[image:5.585.321.516.69.443.2]Taken together, these data demonstrate a correlation be-tween the physical particle release and infectivity of specific supernatants. This suggests that those N-glycosylation mutants that support PFV particle release display normal envelope
FIG. 5. Infectivity of viral supernatants. The infectivities of super-natants of 293T cells cotransfected with the indicated PFV Env ex-pression constructs and A) the EGFP-exex-pression FV vector pMH118 or B) the-galactosidase-expressing FV vector pMH120 are shown. For calculation of the relative infectivities by the EGFP transfer assay, the values for EGFP-positive target cells obtained by using the wild-type PFV Env expression plasmid were arbitrarily set to 100%. The mean values and standard deviations for two independent experiments with a total of 8 to 12 values for each individual Env protein are shown.
on November 8, 2019 by guest
functions such as receptor binding, fusion, and entry into target cells.
Analysis of the⌬N8,⌬N13, and⌬N15 mutant phenotypes by alternative mutations.Next we examined whether the lack of glycosylation at N8, N13, and N15 per se is responsible for the strong phenotypes observed for corresponding mutants or whether the regions surrounding these sites influence Env function, similar to what was observed for Friend murine leu-kemia virus (12, 15). Therefore, we generated additional mu-tations, either abolishing glycosylation by altering the third amino acid in the N-X-S/T signal sequence (⌬N8.1,⌬N13.1, and⌬N15.1) or introducing a different amino acid at the vari-able second position (⌬N8.2,⌬N13.2, and⌬N15.2) that should not interfere with N glycosylation (Fig. 1B). Mutant ⌬N11, lacking one N-glycosylation site but showing no obvious phe-notype, was used as an additional control in the following experiments. Western blot analysis revealed a mobility shift of the gp130Envprecursors in cell lysates of mutants⌬N8,⌬N8.1,
⌬N13,⌬N13.1,⌬N15,⌬N15.1, and⌬N11 (Fig. 6A, lanes 2, 3, 5, 6, 8, 9, and 11) compared to wild-type protein (Fig. 6A, lanes 1 and 12). In contrast, no mobility shift was observed for mutants⌬N8.2,⌬N13.2, and⌬N15.2 (Fig. 6A, lanes 4, 7, and 10), indicating that unlike the other mutants, they still con-tained N-linked carbohydrate chains at the indicated glycosyl-ation sites. LP processing of mutants⌬N8.1 and⌬N13.1 was similar to that of mutants⌬N8 and⌬N13, respectively (Fig. 6C, lanes 2, 3, 5, and 6), whereas that of mutant ⌬N15.1 was significantly improved compared to that of mutant⌬N15 (Fig. 6C, lanes 8 and 9). The other mutants,⌬N8.2,⌬N13.2,⌬N15.2, and ⌬N11, showed LP processing similar to that of the wild type (Fig. 6C, lanes 4, 7, 10, 11, and 12). Subsequently, the infectivity of the supernatants of the transfected 293T cells was examined using the EGFP transfer assay (Fig. 6E). Mutant
⌬N8.1 showed a similar infectivity as mutant⌬N8, and mutant
⌬N13.1 showed a marginal eightfold increase compared to mutant⌬N13, whereas mutant⌬N15.1 displayed an infectivity that was 70-fold higher than that of mutant⌬N15 but still at only about 4% of the level of the wild type. In contrast, mu-tants ⌬N8.2, ⌬N13.2, and ⌬N15.2 showed dramatically in-creased infectivities. Similar to the control mutant⌬N11, mu-tants⌬N8.2 and⌬N13.2 reached levels of 70 to 80% of wild type, whereas the infectivity of⌬N15.2 was only about 20% of wild type. Analysis of particle release revealed in general a good correlation to the infection analysis (Fig. 6D and E). Particle secretion of mutants ⌬N8.2, ⌬N13.2, ⌬N15.2, and
[image:6.585.62.262.79.625.2]⌬N11 into the supernatant was indistinguishable from that of the wild type (Fig. 6D, lanes 4, 7, 10, 11, and 12). No particle release could be detected for mutants⌬N8 and ⌬N8.1 (Fig. 6D, lanes 2 and 3), whereas that of mutants⌬N13 and⌬N15 (Fig. 6D, lanes 5 and 8) was strongly reduced. In contrast,
FIG. 6. Analysis of selected PFV Env mutants. A to D) Western blot analysis of 293T cell (cell) and purified PFV particle (virus) lysates using anti-PFV Gag and anti-PFV Env LP (aa 1 to 86)-specific poly-clonal rabbit antisera. The individual PFV proteins are indicated. E) Infection analysis of supernatants of 293T cells cotransfected with pMH118 and the individual PFV Env expression constructs as indi-cated. For calculation of the relative infectivities by the EGFP transfer assay, the values of EGFP-positive target cells obtained by using the wild-type PFV Env expression plasmid were arbitrarily set to 100%. The mean values and standard deviations for two independent
exper-iments with a total of 4 to 12 values for each individual Env protein are shown. Cotransfection was with pMH118 and the following: lane 1, pczHFVenvEM002 (wt); lane 2, pczHFVenvEM109 (⌬N8); lane 3, pczHFVenvEM131 (⌬N8.1); lane 4, pczHFVenvEM151 (⌬N8.2); lane 5, pczHFVenvEM114 (⌬N13); lane 6, pczHFVenvEM132 (⌬N13.1); lane 7, pczHFVenvEM152 (⌬N13.2); lane 8, pczHFVenvEM116 (⌬N15); lane 9, pczHFVenvEM133 (⌬N15.1); lane 10, pczHFVen-vEM153 (⌬N15.2); lane 11, pczHFVenvEM112 (⌬N11); lane 12, pc-zHFVenvEM002 (wt); lane 13, pcDNA3.1⫹zeo (pcDNA).
on November 8, 2019 by guest
http://jvi.asm.org/
particle release of mutants⌬N13.1 and⌬N15.1 (Fig. 6D, lanes 6 and 9) was increased but still lower than that of the wild type (Fig. 6D, lane 12). Interestingly, significantly higher amounts of secreted particles of mutant⌬N13.1 compared to mutant
⌬N13 could be detected (Fig. 6D, lanes 5 and 6), although the infectivities of these mutants were only marginally different (Fig. 6E).
Taken together, these data suggest that the N glycosylation at N391(N8) in SU and N782(N13) in TM by itself is important
for PFV Env function, whereas at N833(N15) a combination of
N glycosylation and surrounding sequences seems to influence glycoprotein function.
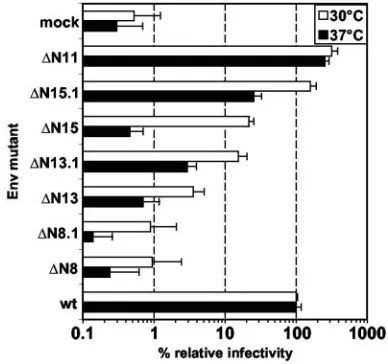
Analysis of temperature sensitivity of the mutant pheno-types.Several viral envelope mutations have been reported to have temperature-sensitive phenotypes (reviewed in reference 2). In particular, mutation analysis of Moloney MuLV N-gly-cosylation sites identified one mutant that displayed a cell type-specific, temperature-sensitive phenotype (5). Therefore, we examined whether some of the PFV Env N-glycosylation mutants showing a decreased infectivity or no infectivity at 37°C could be rescued at lower temperatures. Mutant viruses were produced in parallel by transient transfection of 293T cells at either 37°C or 30°C. Subsequently, target cells were incubated for 6 h with viral supernatants and for an additional 16 h at the corresponding temperature of virus production before shifting of all cells to 37°C and analysis by flow cytom-etry 72 h after infection. The analysis (Fig. 7) revealed a gen-eral two- to fivefold increase in the relative infectivity of most mutants compared to the wild type, although the sensitivity of the assay was somewhat reduced at 30°C due to generally lower infection efficiencies. Nevertheless, mutant ⌬N15 showed a 40-fold increase, and infectivity could be rescued from back-ground levels to 20% of that of the wild type (Fig. 7). The
biosynthesis. Similar to other retroviral glycoprotein precursor proteins, it is proteolytically processed into mature subunits during its transport to the cell surface. However, unlike as reported for other retroviral glycoproteins, two cleavage events, which result in the generation of LP, SU, and TM subunits, take place only late during intracellular transport, most likely by furin-like proteases (4). In this study we analyzed the extent of N glycosylation of the different PFV Env subunits and the function of individual N-glycosylation sites for viral replication. We demonstrate that all potential N-glycosylation sites except one that are present in the PFV Env protein sequence are indeed modified. The LP subunit contains only one carbohydrate chain, whereas the TM and SU subunits have oligosaccharides attached at 3 and 10 sites, respectively. Inter-estingly, the pattern of digestion by different glycosidases sug-gests that the LP and TM subunits contain only carbohydrate chains of the high-mannose or hybrid type, whereas the ma-jority of the oligosaccharides attached to the SU subunit are of the complex type. Recent results on the binding of HIV and Ebola virus GPs to the C-type lectins SIGN and DC-SIGNR suggest a role for high-mannose-type carbohydrate structures in determining the specificity of the interaction (16). Therefore, it might be possible that the specific carbohydrate composition of individual PFV Env subunits, in particular that of SU, may influence binding to target cells and interaction with cellular receptors.
[image:7.585.65.259.71.253.2]Sequence comparison with glycoproteins from other FV spe-cies revealed the conservation of 2 out of 10 N-glycosylation sites in SU and all three sites in TM (data not shown). Muta-tion analysis of the individual glycosylaMuta-tion sites demonstrated that all the nonconserved sites are dispensable for PFV enve-lope function in vitro when inactivated. Interestingly, only mu-tation of some of the conserved carbohydrate attachment sites resulted in detectable phenotypes in our analysis, suggesting that these sites might be important for glycoprotein function of all FV isolates. Mutation of the first (N13) and third (N15) glycosylation site in the TM subunit, which are located in the unique cysteine-rich, prolonged central “sheets-and-loops” re-gion of unknown function (30), resulted in greatly diminished particle release. In addition, both mutants showed a reduced cell-associated LP processing and cell surface expression, in-dicating that the intracellular transport of these proteins is less efficient than wild type. Since the extracellular domain of ret-roviral TM subunits normally harbors the oligomerization do-mains and correct oligomerization of glycoproteins is essential for proper intracellular transport, the results might indicate
FIG. 7. Temperature sensitivity analysis of selected PFV Env mu-tants. The infectivities of supernatants of 293T cells cotransfected with the indicated PFV Env expression constructs generated at 37°C (solid bars) or 30°C (open bars) was determined on HT1080 cells by using the EGFP transfer assay. For calculation of the relative infectivities, the values of EGFP-positive target cells obtained by using the wild-type PFV Env expression plasmid were arbitrarily set to 100%. The mean values and standard deviations for two independent experiments with a total of 4 to 8 values for each individual Env protein are shown.
on November 8, 2019 by guest
that these two mutations affect oligomerization, although this was not addressed directly in our analysis. The results with alternative mutants affected in both glycosylation sites (⌬N13.1 and⌬N15.1) or neighboring sequences (⌬N13.2 and⌬N15.2) suggest that in case of N13 the glycosylation at this position itself is important for Env function. For N15, alternative inhi-bition of glycosylation (⌬N15.1) led to a significant improve-ment of particle release and infectivity, and an amino acid change not affecting glycosylation (⌬N15.2) resulted in a slight reduction in the infectivity of the released particles. Further-more, analysis of the temperature dependence of the pheno-types observed for individual mutants suggests that only the
⌬N15 mutant displays a significant temperature-sensitive phe-notype. Therefore, in this case the sequence surrounding N15 rather than the attached carbohydrate chain, or a perhaps a mixture of both, seems to be important for normal glycopro-tein function. This resembles the case for Friend ecotropic murine leukemia virus Env protein, where only one of eight signals for N-linked glycan attachment is critical for envelope function (12). Subsequently, a more detailed mutational anal-ysis of this MuLV N-linked glycosylation site and surrounding sequences revealed that N glycosylation per se is not required for MuLV Env function but that the region surrounding this glycosylation site mediates envelope folding and the stability of the interaction between SU and TM (15).
In the PFV Env SU subunit, only inactivation of the con-served carbohydrate attachment site N8 impaired Env func-tion. Similar to what was observed for the ⌬N13 and ⌬N15 mutants, the⌬N8 mutant protein was not properly transported intracellularly, as indicated by the lack of LP cleavage of the mutant precursor protein and cell surface expression. How-ever, unlike the two mutants with mutations in TM, neither
⌬N8 nor the alternative glycosylation site mutant ⌬N8.1 showed any detectable infectivity or particle release. In con-trast, the⌬N8.2 mutant, retaining glycosylation but having the variable second amino acid of the N-X-S/T glycosylation site signal sequence mutated, displayed a wild-type phenotype. This strongly suggests that the carbohydrate chain itself serves an important role in PFV Env function, most likely in folding, as both glycosylation mutants showed no proteolytic processing of the precursor protein. However, since this glycosylation site is conserved in the SU subunits of FV isolates from different species, it is possible that it may be involved in other Env function, such as interaction with the cellular receptor, al-though this apparently cannot be addressed in the context of the full-length PFV Env protein due to the transport defect of these mutants. However, preliminary data obtained using PFV Env immunoadhesins indicate that glycosylation site N8 or surrounding sequences might be important for receptor inter-action (A. Duda and D. Lindemann, unpublished observa-tions). Since the digestion pattern of the SU subunit with different glycosidases suggests that the majority of the carbo-hydrate chains in SU are of the complex type, it would be interesting to know which sites are of the high-mannose or hybrid type and in particular what type of carbohydrate chain is added to N8.
ACKNOWLEDGMENTS
This work was supported by grants from the DFG (Li621/2-1, Li621/ 2-3, and Li621/3-1) and the BMBF (01ZZ0102) to D.L.
REFERENCES
1.Baunach, G., B. Maurer, H. Hahn, M. Kranz, and A. Rethwilm.1993. Functional analysis of human foamy virus accessory reading frames. J. Virol.
67:5411–5418.
2.Doms, R. W., R. A. Lamb, J. K. Rose, and A. Helenius.1993. Folding and
assembly of viral membrane proteins. Virology193:545–562.
3.Du Bridge, R. B., P. Tang, H. C. Hsia, P. M. Leong, J. H. Miller, and M. P. Calos.1987. Analysis of mutation in human cells by using an Epstein-Barr
virus shuttle system. Mol. Cell. Biol.7:379–387.
4.Duda, A., A. Stange, D. Lu¨ftenegger, N. Stanke, D. Westphal, T. Pietsch-mann, S. W. Eastman, M. L. Linial, A. Rethwilm, and D. Lindemann.2004. Prototype foamy virus envelope glycoprotein leader peptide processing is mediated by a furin-like cellular protease, but cleavage is not essential for
viral infectivity. J. Virol.78:13865–13870.
5.Felkner, R. H., and M. J. Roth.1992. Mutational analysis of the N-linked glycosylation sites of the SU envelope protein of Moloney murine leukemia
virus. J. Virol.66:4258–4264.
6.Fenouillet, E., J. C. Gluckman, and I. M. Jones.1994. Functions of HIV
envelope glycans. Trends Biochem. Sci.19:65–70.
7.Fenouillet, E., and I. M. Jones.1995. The glycosylation of human immuno-deficiency virus type 1 transmembrane glycoprotein (gp41) is important for the efficient intracellular transport of the envelope precursor gp160. J. Gen.
Virol.76:1509–1514.
8.Fischer, N., M. Heinkelein, D. Lindemann, J. Enssle, C. Baum, E. Werder, H. Zentgraf, J. G. Muller, and A. Rethwilm.1998. Foamy virus particle
formation. J. Virol.72:1610–1615.
9.Geiselhart, V., P. Bastone, T. Kempf, M. Schno¨lzer, and M. Lo¨chelt.2004. Furin-mediated cleavage of the feline foamy virus Env leader protein. J.
Vi-rol.78:13573–13581.
10.Geiselhart, V., A. Schwantes, P. Bastone, M. Frech, and M. Lo¨chelt.2003. Features of the Env leader protein and the N-terminal Gag domain of feline
foamy virus important for virus morphogenesis. Virology310:235–244.
11.Higuchi, R.1990. Recombinant PCR, p. 177–183.InM. A. Innis, D. H. Gelfand, and T. J. White (ed.), PCR protocols; a guide to methods and applications. Academic Press, San Diego, Calif.
12.Kayman, S. C., R. Kopelman, S. Projan, D. M. Kinney, and A. Pinter.1991. Mutational analysis of N-linked glycosylation sites of Friend murine
leuke-mia virus envelope protein. J. Virol.65:5323–5332.
13.Leonard, C. K., M. W. Spellman, L. Riddle, R. J. Harris, J. N. Thomas, and T. J. Gregory.1990. Assignment of intrachain disulfide bonds and charac-terization of potential glycosylation sites of the type 1 recombinant human immunodeficiency virus envelope glycoprotein (gp120) expressed in Chinese
hamster ovary cells. J. Biol. Chem.265:10373–10382.
14.Leurs, C., M. Jansen, K. E. Pollok, M. Heinkelein, M. Schmidt, M. Wissler, D. Lindemann, C. Von Kalle, A. Rethwilm, D. A. Williams, and H. Hanen-berg.2003. Comparison of three retroviral vector systems for transduction of nonobese diabetic/severe combined immunodeficiency mice repopulating
human CD34⫹cord blood cells. Hum. Gene Ther.14:509–519.
15.Li, Z., A. Pinter, and S. C. Kayman.1997. The critical N-linked glycan of murine leukemia virus envelope protein promotes both folding of the C-terminal domains of the precursor polyprotein and stability of the
postcleav-age envelope complex. J. Virol.71:7012–7019.
16.Lin, G., G. Simmons, S. Po¨hlmann, F. Baribaud, H. Ni, G. J. Leslie, B. S. Haggarty, P. Bates, D. Weissman, J. A. Hoxie, and R. W. Doms.2003. Differential N-linked glycosylation of human immunodeficiency virus and Ebola virus envelope glycoproteins modulates interactions with DC-SIGN
and DC-SIGNR. J. Virol.77:1337–1346.
17.Lindemann, D., M. Bock, M. Schweizer, and A. Rethwilm.1997. Efficient pseudotyping of murine leukemia virus particles with chimeric human foamy
virus envelope proteins. J. Virol.71:4815–4820.
18.Lindemann, D., and P. A. Goepfert.2003. The foamy virus envelope
glyco-proteins. Curr. Top. Microbiol. Immunol.277:111–129.
19.Lindemann, D., T. Pietschmann, M. Picard-Maureau, A. Berg, M. Heinke-lein, J. Thurow, P. Knaus, H. Zentgraf, and A. Rethwilm.2001. A particle-associated glycoprotein signal peptide essential for virus maturation and
infectivity. J. Virol.75:5762–5771.
20.Lindemann, D., and A. Rethwilm.1998. Characterization of a human foamy virus 170-kilodalton Env-Bet fusion protein generated by alternative splicing.
J. Virol.72:4088–4094.
21.Pietschmann, T., M. Heinkelein, M. Heldmann, H. Zentgraf, A. Rethwilm, and D. Lindemann.1999. Foamy virus capsids require the cognate envelope
protein for particle export. J. Virol.73:2613–2621.
22.Pietschmann, T., H. Zentgraf, A. Rethwilm, and D. Lindemann.2000. An evolutionarily conserved positively charged amino acid in the putative mem-brane-spanning domain of the foamy virus envelope protein controls fusion
activity. J. Virol.74:4474–4482.
23.Polzer, S., M. T. Dittmar, H. Schmitz, B. Meyer, H. Mu¨ller, H. G. Kra ¨uss-lich, and M. Schreiber.2001. Loss of N-linked glycans in the V3-loop region of gp120 is correlated to an enhanced infectivity of HIV-1. Glycobiology
11:11–19.
24.Polzer, S., M. T. Dittmar, H. Schmitz, and M. Schreiber.2002. The N-linked