0022-538X/94/$04.00+0
Copyright © 1994, American Society for Microbiology
Translational Frameshifting at the gag-pol Junction of Human
Immunodeficiency Virus Type 1 Is Not Increased
in Infected T-Lymphoid Cells
MICHEL
CASSAN,'
NATHALIEDELAUNAY,2t CATHERINEVAQUERO,2
AND JEAN-PIERREROUSSET3*Institut de Genetique etMicrobiologie, Universite Paris XI, 91405 Orsaycedex,1 Institut de laSanteet de laRecherche Medicale U152, Institut Cochin de Genetique Moleculaire, 75014 Paris,2 and Unite de Genetique
de la
Differenciation,
CNRS, URA 1149, Institut Pasteur, 75724 Paris cedex15,3 France Received 16 August 1993/Accepted 22 October 1993A frameshift event is necessary forexpression of the products of the pol gene in a number ofretroviruses, including humanimmunodeficiency virus type 1 (HIV-1). The basic signals
necessary
for frameshifting consist of ashiftysequence in which the ribosome slips and a downstreamstimulatory
structure which can be either a stem-loop or apseudoknot. In HIV-1, much attention has been paid to the frameshift site itself, and only recently has therole of thedownstream structure been examined. Here we used a luciferase-based experimental system toanalyze in vivo the cis and trans factors potentially involved in controlling frameshifting efficiency at thegag-pol junction of HIV-1. We demonstrated that high-level frameshifting is dependent on the presence of apalindromic region located downstream of the site where the frameshift event takes place. Frameshifting efficiencies were foundtobeidenticalinmousefibroblasts and the natural host cells of the virus, i.e., CD4+ humanlymphoid cells. Furthermore, no increase inframeshifting was observed upon virus infection. Previous observations have shownthatviral infection leads to specific alteration of tRNAs involved in translation of shifty sites (D. Hatfield, Y.-X. Feng,B.J. Lee,A.Rein, J. G.Levin, and S. Oroszlan, Virology 173:736-742, 1989). The results presented here stronglysuggestthatthesemodifications do notaffect frameshifting efficiency.
For most mRNAs, the encoded protein is a simple conse-quence ofordered reading of triplet codons. However, addi-tional instructionsarefound in thecoding sequences of certain messages, and these permit the generation of more than one translational product. For example, the meaning of a stop codon can be altered so that an amino acid is incorporated (readthrough), the reading frame may shift (frameshifting), and entire regions of the message can be skipped (hopping) during translation (1). These phenomena of reprogrammed genetic decoding have been recently termed recoding (20). Recoding often results in thesynthesisoftwodifferentproteins from thesamemRNA,asfound in the -1frameshifting in the dnaX gene of Escherichia coli (42). It can also provide fine-tuningof gene regulation, as in the E. coli RF2 translation terminationfactor,inwhichanobligatory +1frameshiftevent
is necessary for expression (9). However, it is in animal retroviruses(27,47)andplantRNAviruses(35),and in many bacterial (12, 36) and yeast (2, 10, 23) transposons, that recoding ismostoften found.
Theretroviral lifecyclerequires readthroughor frameshift-ing forsynthesis of the pol gene products(reviewed in
refer-ence 1). In human immunodeficiencyvirus type 1 (HIV-1), a
single AUG codon initiates translation of the polycistronic mRNAwhich encodes both theGagproteinand theGag-Pol polyprotein (26).Classicalreadingof the first frame
(gag)
leadstoastopcodon,preventingsynthesisof thepol geneproducts.
A -1 ribosomal frameshift, 200 nucleotides upstream of the gag stopcodon, is responsible for the synthesis ofa 160-kDa
*Correspondingauthor.Mailing address:Institut de Genetiqueet
Microbiologie,Universite Paris XI,91405Orsaycedex,France.Phone: 331 694172 64. Fax: 33 1 69 41 72 96.
tPresent address: Joint Diseases
Laboratory,
Montreal, Quebec,Canada H3G 1A6.
Gag-Pol fusion protein. The polyproteinis laterincorporated into virions and cleaved into its maturecomponents. Frame-shifting is thusresponsiblefor theproductionofapreciseratio ofGag-Pol comparedwithGag. This ismostprobablycrucial forbuilding ofafunctional virus (11, 18), making this step a
potential target fortherapeutic blocking of the viral life cycle. The -1frameshiftsignal hastwoelements:(i)a heptanucle-otide sequence,XXXY YYZ, where theribosome shifts and (ii) adownstream stimulatorystructurewhichcanbe either a
stem-loop or a pseudoknot (4, 5, 25). For HIV-1, such a
potential stem-loopis present 8nucleotides downstream of the heptanucleotide (29). However, dependingonthe experimen-tal system used, variations in the effect of this potential stem-looponframeshiftingefficiencyhave beendescribed.In a
yeastsystem (46), a21-nucleotide sequencelackingthe
stem-loopregionwasfoundtobesufficienttopromoteahigh level offrameshifting at thegag-poljunction. Similar results were
found withE. coli, in which disruptionof the stem-loopleads only to a slight decrease of frameshifting efficiency (45). However, inmammaliancells, in vivoanalysiswith frameshift-dependent luciferase constructs showed that a low level of frameshifting is obtained in the absence of the stem-loop region(6),whereas ahighlevel isobtained in its presence(38, 43). Similarconclusionsweredrawn from invitro and in vivo experiments with the whole gag-pol region of HIV-1 cloned
intoanexpressionvector(34).However, in thiswork,theeffect of the potential stem-loop structure was much more pro-nounced in vivothan invitroandthe magnitudeoftheeffect dependedonthe cell type used.
Although the roleof the HIV-1
palindromic
sequenceas astimulator of frameshifting can be considered to be estab-lished, experiments are needed to
quantify
its effect moreprecisely. Furthermore, modifications ofcellular components, related to the differentiation or infection status of the
cell,
1501
on November 9, 2019 by guest
http://jvi.asm.org/
TABLE 1. Sequencesof theoligonucleotidesused in thisstudy'
Oligonucleotide Sequence
89(w)... CTAGCCAGGCTAATTTTTTAGGGAAGATC
89(c)... GATCGATCTTCCCTAAAAAATTAGCCTGG
90(w)... CTAGCCAGGCTAATTTTTTAAGGGAAGATC
90(c)... GATCGATCTTCCCTTAAAAAATTAGCCTGG
1789(w)... CTAGCCAGGCTAATTTTTTAGGGAAGATCTGGCCTTCCTACAAGGGAAGGCCAGGGAAG
1789(c)... GATCCTTCCCTGGCCTTCCCTTGTAGGAAGGCCAGATCTTCCCTAAAAAATTAGCCTGG
1815(w).CTAGCCAGGCTAATTTTTTAGGGAAGATCTGGCCTTCTTGCAACGCAAAGCACGCGAAG
185 w ...CT G A G T A T T T G G A A C G C T C T C A G A A C C C A G
1815(c)... GATCCTTCGCGTGCTTTGCGTTGCAAGAAGGCCAGATCTTCCCTAAAAAATTAGCCTGG
BLV(w)....CTAGCCAGGCTAATTTTTTAGGGAAGATCTGGGGGGACTTAGCGCCCCCCAGGAAG
BLV(c)... GATCCTTCCTGGGGGGCGCTAAGTCCCCCCAGATCTTCCCTAAAAAATTAGCCTGG
'Viral sequencesareinboldletters, andplasmidsequencesareinplainletters.
could interfere directly or indirectly with the frameshifting
mechanism, Forinstance, Hatfield andcoworkershaveshown
that tRNAsInvolved inframeshift signals lack specific hyper-modifications in the anticodon loop when isolated from
in-fected cells (2-t,. In addition to tRNAs interacting with the
shifty site, factors able to bind the downstream secondary
structure could alsobetrans-acting candidate molecules.
Here, we precisely evaluated the effect of the stem-loop
structure in vivo and assessed the role of cell type and virus infectiononframeshift efficiencyatthe HIV-1gag-pol junction.
To achieve this goal, we used a cloning-expression vector
carrying the lucgenefrom Photinuspyralisas areporter,which allows accurate quantification of frameshifting efficiency in
vivo(6).Afivefold increase in theframeshift levelwasinduced by thepresenceofthestem-loopstructure,althoughnoeffect of cell type orvirus infectionwasdemonstrated.
MATERIALSANDMETHODS
Cell lines and culture conditions. The A3.01 cell line is a
hypoxanthine-aminopterin-thymidine-sensitive derivative ofa
CD4+ T-cell line isolatedfrom achildwithacutelymphocytic leukemia (15). It is susceptible to HIV-1 infection (15, 16).
Two chronically infected sublines have been derived from A3.01: ACH-2, which carries a wild-type viral genome, and
8E5, which contains a Pol-defective genome (7, 16). These lines normally produce few (8E5), if any (ACH-2), viral
particles, but highlevels ofvirusreplicationandexpressioncan beinducedby various T-cellactivation stimuli(6, 7, 39, 40, 41). Suspension cultures of cells of the A3.01 lineage were
maintainedin RPMImedium containing10% fetal calfserum in open flasks at 37°C in a 10% CO2 atmosphere. NIH 3T3 mousefibroblastswerecultured asdescribed previously(6).
Plasmids. pCH110 carries the lacZ gene under control of
the simian virus 40 promoter-enhancer region (22). The pRSVLCAT vectorcarries the cat gene under control of the Rous sarcoma virus long terminal repeat promoter (21).
pRSVL, from which the pRSVL74 initialvector was derived,
carries awild-type luc gene(6).
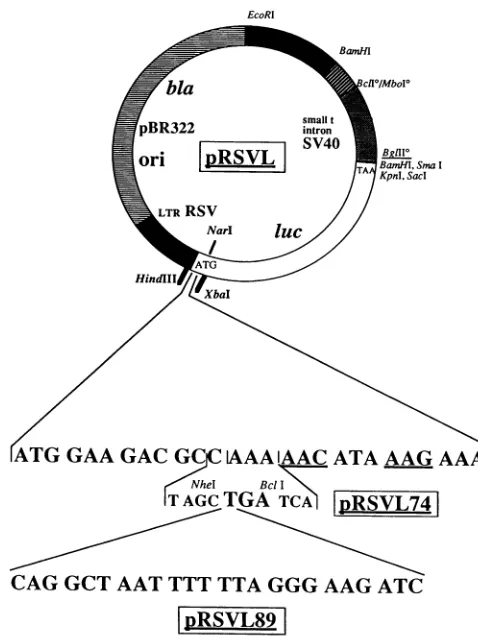
The testplasmidsareallderivedfromthepRSVL74 cloning and expression vector (reference 6 and Fig. 1). This vector
contains an oriented cloning site composed ofa uniqueNheI site and a unique BclI site, including a TGA stop codon,
spanning the fourth to the seventh codons of the luciferase
gene. These unique sites allow insertion of sequences to be tested for frameshifting efficiency. Pairs of complementary
oligonucleotides including an NheI site and Bcll-compatible
stickyends were allowedtoreassociate and then insertedinto
thepRSVL74vector.Insertion leadstodestruction of theBcll site andconsequentlytolossof thestopcodonincludedinthis site. The sequences of the oligonucleotides used are listedin Table 1. Theresultingvectors aretermed pRSVL, followedby thenameof theoligonucleotide.To constructthepRSVL1790
and pRSVLTBLV control vectors, the newly created NheI-BglII fragment from either pRSVL1789 or pRSVLBLV was
replaced by the appropriate 90 (w) and 90 (c) paired oligo-nucleotides. Figure 1 illustrates the cloning strategy by using the construction of the pRSVL89 vector as an example. Positiveclones were screened by usingoneofthe oligonucle-otides as a probe, and the nucleotide sequence of a 100-bp regionsurrounding the insertwasverified.
Transfections. Plasmid DNAs used in transfections were
submitted to two successive CsCl-ethidium bromide equilib-rium centrifugationsteps.
It wascriticaltoachievehigh transfection efficiencytodetect
lowlevels offrameshift-dependent luciferase activity.For NIH
3T3 cells,we havepreviously shownthattheclassical calcium phosphate coprecipitation techniquegavesatisfactory transfec-tionefficiency (6).NIH3T3cellswerecotransfected with10
Rg
of thetestplasmidsand 5 ,ugofthepCH110calibrationvector. For the A3.01 cell line family, high transfection efficiencies
were obtained by modifyingthe DEAE dextran technique of Fregeau and Bleackley (17). Briefly, 1.25 x 107 cells were
refedwith fresh medium3 hbefore transfection andseededat
10'/ml.
For transfection, cells were centrifuged at low speedand the cell pelletwas rinsed in30 ml of serum-free medium
and then suspended in 1 ml of transfection buffer (0.5 ml of TBS, which contained 25 mM Tris HCl [pH 7.4], 137 mM
NaCl, 5 mM KCl, 0.6 mMNa2HPO4, 0.7 mM CaCl2, and0.5
mMMgCl2, plus 0.5ml ofserum-freeRPMI) containing3
jig
of thepRSVCATcalibrationplasmid,7jig
of the testplasmid,and500
jig
ofDEAEdextranperml.The cellswereincubatedat room temperature for 15 min, rinsed twice with 10 ml of serum-free medium, and resuspensed in 10 ml of culture medium containing 7.5 mM sodium butyrate. No further increase in transfection efficiency was observed when the amountoftheplasmidwasincreasedorwhenchloroquinewas included during the transfection orexpression step.
At 48 (A3.01 family) or72 (NIH 3T3) h aftertransfection, cells were harvested for enzyme assays. Crude extracts were obtained by lysing the pelleted cells in 100 ,ul of luciferase buffer (1% Triton X-100, 25 mM Tris phosphate [pH 7.8], 8
mM MgCl2, 1 mM dithiothreitol, 1 mM EDTA, 1% bovine serum albumin, 15% glycerol).The cell extract wasclearedby
on November 9, 2019 by guest
http://jvi.asm.org/
EcoRI
BamHII
IATG
GAA
GAC
GC
AAAIAAC
ATA AAG AAAiNheI BclI \
T
AGC
TGA
TCA pRSVL741
CAG GCT AAT TTT TTAGGG AAG ATC
|
PRSVL89
FIG. 1. Filiation ofthe differentvectors used in this study,
illus-tratedwith theconstruction ofthepRSVL89vector. The upperpart showsthe mainfeatures of the initial pRSVLvector. The lowerpart showsthesequencesofthe amino-terminal parts ofthelucgenesfrom
the pRSVL wild-type vector,from thepRSVL74 cloningvector, and
from the pRSVL89 test vector. SV40, simian virus 40; LTR, long
terminal repeat.
centrifugation at 12,000 x gfor 20 min, and the supernatant
waskept frozen.
Stimulation of viralexpression.Instimulationexperiments,
cellswereallowed torecoverfor 6 h aftertransfection and then split into two aliquots; one wasstimulated, and the otherwas
used as acontrol. Cells were continuouslystimulated with 50 nM phorbol myristate acetate (PMA) until harvesting at48 h after transfection. These conditions induce both HIV-1
tran-scription and protein expression in infected ACH2 and 8E5 cells but do not trigger significant cytokine expression (41).
Uninfected A3.01control cellsweretransfected andstimulated by following thesameprotocol.
Enzyme assays. Luciferase was assayed by following the protocol ofNguyen (32), witha Berthold LUMAT LB 95501 luminometer forlightdetection. Chloramphenicol
acetyltrans-ferase (CAT) activitywas determined in accordance with the two-phase partitionmethod(31). ,B-Galactosidasewasassayed in accordance with the classical method ofMiller(30).
RESULTS
To measure the in vivo level offrameshifting at the HIV-1
gag-poljunction accurately and to assess the role ofpotential
effectors actingeither in cis or intrans, we useda previously described system ofexpression vectors (6). Sequences
poten-tially subject to frameshifting are inserted at the beginning of the luc gene coding region, such that the appearance of functional luciferase is dependent on a frameshift event. The vectors are transiently transfected into mammalian cells, and the luciferase activity is adirect reflection of theframeshifting
efficiency. This system ishighly sensitive,allowing detectionof frameshift levels as low as 10-5.We previouslydemonstrated
that theluciferaseactivity observedis notdue to reinitiation at the downstream AUG or to accumulation of mutants in the plasmid preparation used for transfections (6).
Since the amino acid sequence of luciferase changes be-tweendifferent constructs, variation of luciferase activitycould reflect eitherdifferent frameshift efficiencyor different specific activity of the modified luciferase. It was therefore crucial to compare the activity driven by a frameshift-dependent con-struct with the activity obtainedwith an appropriate control. The site where the ribosome shifts is precisely known (26),
allowing the construction of in-frame controlvectorsby addi-tion of oneadenine after the UUAleucinecodon (Fig. 1). In these vectors, the luciferase sequence corresponds exactly to thatsynthesized from the test vectors after frameshifting.
Quantification of the HIV-1 frameshift level in vivo. In a first set of experiments, we analyzed in vivo the effect of the
potential stem-loop structure on the efficiency ofthe HIV-1
frameshift.One construct (pRSVL89) carriesa 24-nucleotide sequencecontainingthe minimalslipperyregion of the HIV-1
gag-poljunction defined by Wilson et al. (46), and the other
(pRSVL1789) contains the palindromic sequence as well. Figure 1 describes thegeneral strategyforconstruction of the
different vectors, illustrated with the pRSVL89 vector. The constructs were transfected into NIH 3T3 cells, and the
luciferase activities measured were standardized for
transfec-tion efficiency by using ,B-galactosidase expression directed by
the pCHI 0vector(see Materials andMethods).The results,
expressed as the ratio of the luciferase activity of a given construct to that of the corresponding in-frame control
(pRSVL90 or pRSVL1790), are shown in Fig. 2. A fivefold
increase oftheframeshifting efficiencywasobserved whenthe
palindromic sequencewaspresent.
The palindromic region could be crucial for two reasons:
either becauseofthe potential stem-loop secondarystructure orbecause of the sequence context. To discriminate between these two hypotheses, we replaced the HIV-1 palindromic region by the analogous palindromic region from the bovine leukemia virus (BLV) (pRSVLBLV construct). In another
vector, we introduced several mutations which affect the potential stem of the HIV-1 palindromic sequence
(pRSVL1815). The residual base-pairing potentialinthe latter construct corresponds to a free energy of -8 kcal (1 cal = 4.184 J)/mol (calculated with the MFOLD program by using
Zucker's rules), compared with -20 kcal/mol for the HIV-1
wild-type sequence. Results shown inFig. 3 demonstrate that
the BLV palindromic region can increase frameshifting as
efficiently as the wild-type HIV-1 stimulatorystructure. With
the pRSVL1815 vector, the frameshifting level dropped only twofold.This suggeststhat the nucleotidecontextinfactplays
a role, although we cannot exclude the possibility that the
remainingbasepairingis sufficient in vivotodirect part of the
effect of thewild-type stem-loop structure.
The frameshift level isnotincreased inCD4+ lymphocytes.
Since both the frameshift efficiency and the effect of the
stem-loop structure have been found tovary greatlybetween
different cell types (compare, for example, references 19 and
34), it was essential to analyze the frameshifting efficiency in natural host cells of thevirus, i.e., CD4+lymphocytes.
This analysis necessitated a high transfection efficiency of
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.54.293.69.390.2]pRSVL90 in frame control
+A
t
ATGGAAGACGCTAGCCAGGCTAATTTTTTAGGGAAGATCGATCAAACATAA... Luciferase HIV-1 Luciferase
C A
A A
T.G C-G
C-G
T-A
T-A C-G
pRSVL1790 in frame control C-G
G-C
+A G-C
T-A
AACACAAGC-G
ATGGAGACGCTAGC AGGCTAATTTTTTAG,GGAAGAT*GGAAGGATcAAAcATAA. Luciferase HIV-1
Frameshifting efficiency
pRSVL89 = 0.39
(+/-
0.15) %pRSVL1789 = 2.1
(+/-
0.5) %Luciferase
FIG. 2. Amino-terminalpartoftheengineeredlucgenewith the HIV-1frameshiftregionunderlined. The first stop codonencounteredinthe
normal frame of theengineered lucgeneisin boldletters.NIH3T3cellsweretransfectedwith thedifferentvectorsand harvested 3 dayslater forenzymaticassays. Therelativeexpressionofeachconstructisexpressedasthe ratio of theluciferaseactivity directed by the -1frametest construct(pRSVL89orpRSVL1789) tothat directedbythe in-framecontrol (pRSVL90orpRSVL1790). Results werestandardizedby using
P-galactosidaseexpressionas aninternalcontrol for transfectionefficiency. Each valuerepresentsthemeanofatleast sixindependenttransfection experimentswithtwoindependent plasmid preparations.
CD4+lymphoidcellsfor detection ofalowlevel of
frameshift-dependent expression. We used a DEAE dextran-derived
transfection technique (17) adapted for the A3.01 CD4+ family ofcell lines(seeMaterials and Methods). By usingthis
method, activities ofupto106light units/107 cells/,ugofvector can be obtained with thewild-type pRSVLvector. Since the
backgroundoftheassayformock-transfectedcells isbetween 80 and 150 light units, it allows detection and accurate
pRSVL1790 inframe contro
quantification of frameshift levels as low as 10-4 in typical experimentswith 7,ug ofatestplasmid.
In transfections involving cells of the A3.01 family, the
pRSVCATvector, which bearsthe cat geneunder control of the Rous sarcoma virus long terminal repeat promoter, was
usedasacalibrationvector.ThepCH110vectorused in NIH 3T3cellswasinoperativein theA3.01-derivedcell lines for the
following two reasons: (i) because of a low but systematic
C A
A A
T.G C-G C-G T-A T-A C-G C-G G-C
+A G-C
T-A
A C-G
T T.G
ATGGAAGACGCTAGCCAGGCTAATTTTTAGGGAAGA GAAGGATCAAACATAA... Luciferase HIV-1 Luciferase
Frameshifting efficiency
pRSVL1789 = 2.1
(+/-
0.5) % T AT G
C C
pRSVLTBLVin fram control
+A
ATGGAAGACGCTAGCCAGGCTAATTTTTTiG
Luciferase HIV-J1 G-C G-C
G-C
G-C BLV
G-C G-C T-A C-G
;GGAAGA GAAGGATCAAACATAA...
Luciferase pRSVLBLV
= 2.6 (+/- 0.4) %
C A G A
T C
T.G
C C
T-A T-A C A
C-G
pRSVL1790 inframe control G-C
+A G A
T C
C-G
ATGGAAGACGCTAGCCAGGCTAATTTTTT GGGAAGA
GAAGGATCAAACATAA-Luciferase HIV-1 Luciferase
[image:4.612.130.497.76.232.2]pRSVL1815 = 0.9
(+/-
0.2) %FIG. 3. SameasFig. 2, but in thiscasethreeindependent experimentsweredone.
A
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.129.485.425.700.2]TABLE 2. Frameshiftingefficienciesinuninfectedandinfectedlymphoidcells
Cell line Construct Frameshifting efficiencies(%)a Mean frameshifting
efficiency(%; ±SEM)
A301 pRSVL89 0.15,0.19, 0.29, 0.21, 0.43 0.25(±0.11)
pRSVL1789 0.90,0.90, 1.02, 0.81, 2.00 1.13(±0.49)
ACH2 pRSVL89 0.80,0.12, 0.21, 0.26, 0.52, 0.19, 0.14 0.32(±0.25)
pRSVL1789 4.00, 1.50, 1.20, 1.10, 1.40, 1.10,1.70 1.71(± 1.03)
8 E5 pRSVL89 0.22,0.17, 0.29, 0.26, 0.86 0.36(±0.28)
pRSVL1789 1.00,0.97, 2.50, 2.40, 1.90 1.75(±0.74)
a Resultsofindividual transfectionexperimentsareexpressedas the ratio ofluciferase activity obtainedwith a testvectorto thatobtained withthecorresponding
in-frame control. Resultsare standardizedfortransfectionefficiency with respectto CATexpression.
background of 3-galactosidase activity in these cells and (ii) becauseno ,B-galactosidase activity above the backgroundwas
detected,evenin experimentsin whichacotransfectedpRSVL vectorwasabletodriveahighlevel of expression. Thiswasnot duetolack ofexpression of the simian virus40promoter,since
thesameresults have beenobtained withvectorscarrying the
lacZgeneunder control of either the Roussarcomavirus long
terminal repeat promoter, or the cytomegalovirus promoter
(data notshown).
pRSVL89 and pRSVL1789 and in-frame control plasmids pRSVL90 and pRSVL1790weretransfected into A3.01 cells.
Luciferase activities were standardized for transfection
effi-ciency by using CATas aninternal control. Results expressed as the luciferase activity obtained with the test vectors
com-pared withthat obtained with the in-frame controlsareshown
in Table 2. Although for thetwo vectorsthe frameshift level
observedinA3.01 cellswasslightly lower than that observedin
NIH 3T3 cells (0.25 versus 0.39% with pRSVL89 and 1.1
versus 2.1% with pRSVL1789), this difference is not
signifi-cant. In the presence of the stem-loop structure, a fivefold
increase in frameshifting efficiencywasobserved inA3.01, as
well asin NIH 3T3, cells. Thus,weobserved no influence of
cell typeonframeshifting efficiency. Inaddition, theseresults
illustrate thatNIH3T3 cellsaregoodrecipients for analysis of
ciselements involved in frameshifting inmammalian cells.
Theframeshiftlevel isnotincreasedinchronically infected cells. Hatfield and coworkershave reportedthat HIVinfection leads to tRNA hypomodification. They suggested that these
alterations could beresponsiblefor anincreasein
frameshift-ingby direct perturbation of codon-anticodon recognition (24). Frameshifting could also be regulated by virus infection
through interaction of the viral RNA with virus-encoded or
virus-induced factors, agood potential target beingthe
stem-loop structure. In our vectors, since the target shifty site is uncoupled from viral genome expression, analysis of
frame-shifting efficiency can be carried out independently of the
presence oftheviralgenome.
Totestthe effect of virus infectiononframeshifting,weused twochronicallyinfected lines derived from A3.01 cells(15, 16).
TheACH-2line carriesawild-typeviralgenome,and the8E5
line contains a Pol-defective genome (7, 16). The cells were
transfected under the conditions described above with the minimal pRSVL89 construct and the pRSVL90 control and
with thepalindromecontaining the pRSVL1789vectorand the
pRSVL1790 control. Results summarized in Table 2 showa
slight increase of the frameshift level in infectedversus
unin-fectedcells. Althoughobservedwith these two independently
infected cell lines, this effect is not significant. The same
fivefoldincrease inframeshifting efficiencyduetothepresence
of thepalindromic regionwasalso found with thetwoinfected
cell lines (Table 2). The frameshift efficiencies observed in uninfected and infectedT-lymphoid cells are thus not different. However, we cannot exclude thepossibility that an effect of virusinfection escaped detectionbecause full virusexpression needs lymphocyte stimulation. Indeed, the ACH2 line pro-duceslittle, if any, infectiousparticles and undetectable levels ofviral mRNA andproteins, and in the 8E5 line the viral RNA and proteins can only be detected at a low level (41). Viral transcription and proteinsynthesis, leading to a burst of virus particles, are induced in these cells by various treatments, includingPMAstimulation(7, 14, 37, 39-41, 44).
We analyzed the effect of the stimulation of virus gene expression by PMAon the frameshift level in theACH2 and 8E5 lines. As a control, we determined the effect of PMA stimulation on frameshifting efficiency in uninfected A3.01 cells. Cells were transfected with thepRSVL1789vector and split into two flasks 6 h later. One sample was subjected to stimulation with PMA, and the otherwas used as a control. Results areexpressed as the ratio of luciferaseactivitytoCAT activity in stimulated and control cells. No significant
differ-ence in frameshifting was observed between stimulated and control cells (Table 3). Consequently, induction of virus
ex-pression doesnotinfluenceframeshifting efficiency. DISCUSSION
HIV-1, like many otherretroviruses,usesaprogrammed -1 frameshift to express thepol geneproducts (26). Two
struc-tural elements are involved in controlling frameshifting effi-ciency:ashiftysite where the ribosomeslipsandadownstream stimulatorysequencewhichcan
potentially
adoptastem-loop conformation. Although much attention has beenpaidtothe frameshift site itself, only recently has the role of thedown-streamstructure been examined(34).
TABLE 3. Effect ofstimulationwithPMAonframeshifting efficiencyin uninfected and infectedlymphoidcells
Cell line Luciferase Meanluciferaseactivity (treatment) activitiesa (±SEM)
A301 31, 12,14 19(±11)
A301 (PMA) 25, 34,23 27(±6)
ACH2 26, 15,52 31(±19)
ACH2(PMA) 46, 94,53 64(± 26)
8E5 31, 27,87 29(33)
8E5(PMA) 16,57 36(29)
aResults areexpressedinarbitraryunitsof luciferaseactivityobtained with thepRSVL1789 vector,corrected for transfectionefficiencywithrespecttoCAT expression.The differentvaluescorrespondtoindependenttransfection exper-iments.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.319.555.603.684.2]In this study, we used frameshift-dependent expression
vectorsbasedonluciferasetoquantify preciselyinvivo the role of thepotential stem-loopstructureofHIV-1.With this system
we obtained both a high level of frameshifting (2.1%) and significantdependence onthepalindromic region(fivefold)in
NIH 3T3 mouse fibroblasts. This is in contrast to in vitro translation systems or in vivo bacterial or yeast systems, in
which no oronlyaslight effect ofthe palindromic region has beenfound (34, 45,46).
A fivefold increase was also obtained when the analogous palindromic region derived from BLV (pRSVLBLV vector)
wasused instead ofthe HIV-1palindrome. This is reminiscent of thefindingof Parkinet al.(34),who showed thatreplacing the HIV-1 stem-loop by the pseudoknot from mouse
mam-mary tumorvirus, which would give a morestable secondary structure, resulted in afivefold increase in frameshifting effi-ciency. Altogether, theseresults demonstratethat, invivo, the palindromic regionis importantfor ahigh-levelframeshift to
takeplace. The palindromicregionmostprobablyactsthrough formation of a stem-loop structure. However, we cannot
exclude the possibilitythat thenucleotide sequence itself also playsarole, sinceonlyatwofolddecrease in frameshiftactivity
was obtained when most of the pairing capability was de-stroyed by mutation. Constructs inwhich an anonymous pal-indrome replaced that ofHIV-1 would help in the testing of this hypothesis.
Anotherproblemwhich isonlybeginningtobe investigated is the involvement of trans-acting factors in controlling the frameshift mechanism. As mentioned above,the effect of the potential stem-loop is more important in vivo than in vitro. This could reflect the need for some trans-acting molecule, onlypresent (or enriched) inliving cells,which interacts with the stem-loopstructure. Since the effects of thestem-loopare
different in different cell types (compare references 6, 34, 38, and 43), such trans-actingfactors could be differently
distrib-uted indifferent tissues. Accordingly, ahigherlevel of frame-shifting mighttakeplace in the naturalhost cells ofthevirus, i.e., CD4+ lymphocytes.
Here,wequantifiedtheframeshiftinglevel andanalyzedthe effect of the stem-loop structure in cells of the A3.01 family, derived fromaCD4+ lymphoidtumor.Comparableframeshift levelswereobserved in the A3.01cell lineand NIH3T3mouse
fibroblasts. This is consistent with other studies, in which
HIV-1 frameshifting was estimated in different cell types by using frameshift-dependent reporter expression (38, 39a, 43). The frameshift levelsrangedbetween 1 and
2%,
reflectingthe inherent variations of transfection techniques. Also, weob-served the same fivefold increase due to the palindromic sequence inA3.01 and NIH3T3 cells.Together,these results indicate that similar mechanisms control frameshift levels in
different cell types and provide nosupport for thehypothesis
that specific trans-activating factors exist in the natural host cells ofHIV-1.
Hatfield and coworkers showed that tRNAs involved in
frameshift signals from different viruses lack specific hyper-modifications in their anticodon loop when isolated from infected cells (24). This might be relevant for frameshifting, because only tRNAs involved in decoding of the HIV-1 frameshift site are undermodified in HIV-1-infected cells, whereasonlytRNAs involved indecoding of the BLV frame-shift site are undermodified in BLV-infected cells. More generally, one can ask whether virus infection would affect frameshifting whatever the mechanism involved, i.e., either through tRNA modifications or by any other alteration of cell-encoded molecules. We carriedout thisanalysis with our
in vivo system, in which the frameshift target site and viral expression canbeuncoupled.
Weexamined the effect of HIV-1 infectiononframeshifting by transfection ofchronically infected cells. Both the frame-shift level and the effect of the stem-loop structure were
equivalent in infected and control cells. No effect on frame-shiftingwasobserved uponstimulation of virusexpressionwith
PMA.Nevertheless,we cannotexcludethepossibilitythatsuch
an effect exists. Forexample,the highnumber of target RNA molecules per cell may lead to titration of a putative
trans-acting factor in transfection experiments. The effect of virus infection could also be dependent on the presenceofaviral
RNAtargetregionwhich isnotincludedin theconstructsthat
we used. Finally, the posttranscriptional modifications of the
tRNAscouldconceivablybedifferent inourcell system and in the cells usedby Hatfield and coworkers.
However, thesimplest interpretation ofour results is that virus infection hasnoeffectonframeshifting. Thisconclusion is confirmedby the work of Reiletal.(37a),who usedahuman fibroblastic cell linestably transfected witha frameshift-depen-dent construct and rendered susceptible to HIV-1. In this experimentalsystemalso,HIV-1infection didnotsignificantly modify the frameshifting efficiencyattheHIV-1gag-pol junc-tion.
It has been proposed that in Moloney murine leukemia virus, virus infection increases theamountofaminor
suppres-sortRNAresponsibleforreadthroughat thegag-pol junction (28), but this result has been disputed (13). Furthermore, experiments of the type presented here have never revealed anyeffect ofMoloney murine leukemia virus infectiononthe readthroughlevelatthegag-pol junctionof type C retroviruses (3, 33). Thus, in programmed readthrough as well, viral infection hasnoeffect.
Upto now, the evidence for modification of cell-encoded molecules upon virus infection has come from biochemical analysis; however,functional studies have failed toreveal any effect of infection on frameshifting at the gag-pol junction. Since theexpressionofthe TopenreadingframeofHIV-1 is probably dependenton a -1 frameshift(8),tRNAalterations may act atthis step. Anotherpossibility is thattheyinterfere withthe viruscycle at alevel different fromframeshifting.
Differentapproaches have been used toanalyze thesignals that modulate HIV-1 frameshifting: in vitro translation of synthetic RNA orinvivo systemsusingeitherbacterial,yeast,
or mammalian cells as a host. In parallel, frameshifting effi-ciencyhas been evaluatedby assaying the level of expression of
a frameshift-dependent reporter (,B-galactosidase or lucif-erase) orthrough direct quantification of theprotein synthe-sized. Comparison of these different approaches leads to
apparently contradictory results. For instance, in yeasts and bacteria, no effect of the palindrome was observed (45, 46),
whereas in a mammalian cell system increased frameshifting
wasobserved in the presence of the palindromic region(34).
Butin thislastexperiment, the overall efficiency of frameshift-ingwas low. Last, in experiments conducted invitro, only a
slight effect or no effect at all of thepotential stem-loop was
observed (34, 46).
Invivo systems likethe one usedhere,inwhichexpressionof
a reporter gene is dependent on a frameshift event, give
accurate and consistent results. Different groups using such systemshavereportedresultswhich areinperfect agreement, bothqualitativelyandquantitatively (6, 38).Furthermore, they alsopermit studyofthe effect ofpotential trans-acting factors. Therefore, today theyconstitute the most powerful approach
toanalysisof theframeshift mechanism inhigher eukaryotes.
on November 9, 2019 by guest
http://jvi.asm.org/
ACKNOWLEDGMENTS
We thankMarguerite Picard and Mary C. Weiss for support during the course of this work. Roger Karress, Anne Plessis, and Mary C. Weiss greatly contributed to theimprovement of successive versions of the manuscript and are gratefully acknowledged.
This work wassupported by the CEE (Bridge Programme) and the Association pourla Recherche contre le Cancer.
REFERENCES
1. Atkins, J. F., R. B.Weiss, andR. F. Gesteland. 1990.Ribosome gymnastics-degree of difficulty 9.5, style 10.0. Cell 62:413-423. 2. Belcourt, M. F., and P. J.Farabaugh. 1990.Ribosomal
frameshift-ing in the yeast transposon Ty: tRNAs induce slippage on a 7 nucleotide minimal site. Cell 62:339-352.
3. Berteaux, V., J. P. Rousset, and M. Cassan. 1991. UAG readthrough is not increased in vivo by Moloney murine leukemia virus infection. Biochimie 73:1291-1293.
4. Brierley, I., P.Digard, and S. C.Inglis. 1989.Characterizationof anefficient coronavirus ribosomal frameshift signal: requirement for an RNApseudoknot. Cell 57:537-547.
5. Brierley, I., A. J. Jenner, and S. C. Inglis. 1992. Mutational analysis of the "slippery-sequence" component of a coronavirus ribosomal frameshift signal. J. Mol. Biol. 227:463-479.
6. Cassan, M., V.Berteaux, P.0. Angrand, and J. P. Rousset. 1990. Expression vectors forquantitating in vivo translational ambiguity: their potential use to analyse frameshifting at the HIV gag-pol junction. Res. Virol. 141:597-610.
7. Clouse, K. A., D. Powell, I. Washington, G. Poli, K. Strebel, W. Farrar, P.Barstad, J.Kovacs,A. S.Fauci, and T. M. Folks. 1989. Monokine regulation of human immunodeficiency virus-1 expres-sion in a chronically infected human T cell clone. J. Immunol. 142:431-438.
8. Cohen, E. A., Y. Lu, H. Gottlinger, G. Dehni, Y. Jalinoos, J. G. Sodroski, and W.A.Haseltine. 1990. The T open reading frame of humanimmunodeficiency virus type 1. J. Acquired Immune Defic. Syndr. 3:601-608.
9. Craigen, W. J., and C. T. Caskey. 1986. Expression of peptide chain release factor 2 requires high-efficiency frameshifting. Na-ture(London) 322:273-275.
10. Dinman, J. D., T.Icho, and R. B.Wickner. 1991. A -1 ribosomal frameshift in a double-stranded RNA virus of yeast forms a gag-pol fusion protein. Proc. Natl. Acad. Sci. USA 88:174-178. 11. Dinman, J. D., and R. B.Wickner. 1992. Ribosomalframeshifting
efficiency andgag/gag-pol ratio are critical for yeast MI double-stranded RNA virus propagation. J. Virol. 66:3669-3676. 12. Fayet,O., P. Ramond, P. Polard, M. F. Prere, and M. Chandler.
1990. Functional similarities between retroviruses and the IS3 family ofbacterial insertion sequences. Mol. Microbiol. 4:1771-1777.
13. Feng, Y.-X., D. L. Hatfield, A. Rein, and J. G. Levin. 1989. Translational readthrough of the murine leukemia virus gag gene amber codon does not requirevirus-induced alteration of tRNA. J. Virol. 63:2405-2410.
14. Fernie, B. F., G. Poli, and A. S. Fauci. 1991. Alpha interferon suppresses virion but not soluble human immunodeficiencyvirus antigen production in chronically infected T-lymphocytic cells. J. Virol. 65:3968-3971.
15. Folks, T., S. Benn, A. Rabson, T. Theodore, M. D. Hoggan, M. Martin, M. Lightfoot, and K. Sell. 1985. Characterization of a continuous T-cell line susceptible to the cytopathiceffects of the acquiredimmunodeficiency syndrome-associated retrovirus. Proc. Natl.Acad. Sci. USA82:4539-4543.
16. Folks, T. M., D. Powell, M. Lightfoot, S. Koenig, A. S. Fauci, S. Benn, A.Rabson, D.Daugherty, H. E. Gendelman, M. D. Hogan, S.Venkatesan, and M. A. Martin. 1986. Biological and biochem-ical characterization of a cloned Leu3- cell surviving infection with theacquired immune deficiencysyndrome retrovirus. J. Exp. Med. 164:280-290.
17. Fregeau, C. J., and R. C. Bleackley. 1991. Factors influencing transientexpression in cytotoxic T cells followingDEAE dextran-mediated gene transfer. Somat. Cell Mol. Genet. 17:239-257. 18. Fujimura, T., J. C. Ribas, A. M. Makhov, andR.B. Wickner. 1992.
Pol of gag-pol fusion protein required forencapsidation of viral
RNAofyeast L-Avirus. Nature (London) 359:746-749. 19. Garcia, A., J. van-Duin, and C. W. Pleij. 1993. Differential
response to frameshift signals in eukaryotic and prokaryotic translational systems. Nucleic Acids Res. 21:401-406.
20. Gesteland, R.F., R. B.Weiss,andJ. F.Atkins. 1992. Recoding: reprogrammedgeneticdecoding. Science 257:1640-1643. 21. Gorman,C.M.,L. F.Moffat,and B. H. Howard. 1982.
Recombi-nantgenomeswhich express chloramphenicol acetyltransferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
22. Hall, C.,E.Jacob, G.Ringold,and F. Lee. 1983. Expression and regulation ofEscherichia coli lacZ gene fusions in mammalian cells. J. Mol.Appl.Genet. 2:101-109.
23. Hansen,L.J.,D. L.Chalker,and S. B.Sandmeyer. 1988. Ty3, a
yeast retrotransposonassociated with tRNAgenes,hashomology
toanimal retroviruses. Mol. Cell. Biol. 8:5245-5256.
24. Hatfield, D., Y.-X. Feng, B.J. Lee, A. Rein, J. G. Levin, andS. Oroszlan. 1989. Chromatographic analysis of the aminoacyl-tRNAswhicharerequired for translation of codonsatand around the ribosomal frameshift sites ofHIV, HTLV-1 andBLV. Virol-ogy173:736-742.
25. Jacks, T.,H. D.Madhani,H. D.Masiarz,and H.E. Varmus. 1988. Signals for ribosomal frameshifting in the Rous sarcoma virus
gag-pol region. Cell 55:447-458.
26. Jacks, T.,M. D.Power,F. R.Masiarz,P. A.Luciw,P.J. Barr,and H. E.Varmus. 1988. Characterization of ribosomal frameshifting in HIV-1 gag-pol expression. Nature (London) 331:280-283. 27. Jacks, T., and H. E. Varmus. 1985. Expression of the Rous
sarcomavirus pol gene by ribosomal frameshifting. Science 230: 1237-1242.
28. Kuchino,Y.,H. Beier,N.Akita,and S.Nishimura. 1987. Natural UAG suppressor glutamine tRNA is elevated in mouse cells infected with Moloney murine leukemia virus. Proc. Natl. Acad. Sci. USA 84:2668-2672.
29. Le, S. Y., J. H. Chen, and J. V. Maizel. 1989. Thermodynamic stability and statistical significance ofpotential stem-loop
struc-turessituatedat the frameshift sitesinretroviruses. NucleicAcids Res. 17:6143-6152.
30. Miller, J.H. 1972. Experiments inmoleculargenetics,p. 352-355. ColdSpringHarbor Laboratory, Cold Spring Harbor, N.Y. 31. Neumann,J.R.,C.A. Morency, and K. 0. Russian. 1987. A novel
rapidassayforchloramphenicol acetyltransferase geneexpression. BioTechniques5:444-447.
32. Nguyen, V. T., M. Morange, and 0. Bensaude. 1988. Firefly luciferase luminescence assays using scintillation counters for
quantitation in transfected mammalian cells. Ann. Biochem. 171: 404-408.
33. Panganiban,A. T.1988.Retroviralgagamber codonsuppression is caused byanintrinsiccis-acting componentofthe viralmRNA.J. Virol. 62:3574-3580.
34. Parkin, N.T., M. Chamorro, and H. E. Varmus. 1992. Human immunodeficiency virus type 1 gag-pol frameshifting isdependent
on downstream mRNA secondary structure: demonstration by expression invivo.J.Virol. 66:5147-5151.
35. Pelham, H. R. B. 1978.Leaky UAG terminationcodon in tobacco mosaic virus RNA. Nature (London) 272:469-471.
36. Polard, P., M. F. Prere, M. Chandler, and 0. Fayet. 1991. Programmed translational frameshiftingand initiationat an AUU codonin gene expression ofbacterial insertion sequenceIS911.J. Mol. Biol. 222:465-477.
37. Qatsha, K.A.,C. Rudolph, D. Marme, C. Schachtele,and W. S. May. 1993. Go 6976,a selective inhibitorofprotein kinaseC, is a potent antagonist of human immunodeficiency virus 1 induction fromlatent/low-level-producing reservoir cells in vitro. Proc. Natl. Acad. Sci. USA 90:4674-4678.
37a.Reil, H., et al.Submitted for publication.
38. Reil, H., and H. Hauser. 1990. Test system for determination of HIV-1 frameshifting efficiency in animal cells. Biochim. Biophys. Acta 1050:288-292.
39. Rieckmann, P., G. Poli, J. H. Kehrl, and A. S. Fauci. 1991. Activated B lymphocytes from human immunodeficiency virus-infected individuals inducevirusexpression in infected T cellsand a promonocyticcell line,UI.J. Exp. Med. 173:1-5.
39a.Rousset, J.-P. Unpublished data.
on November 9, 2019 by guest
http://jvi.asm.org/
40. Serpente, N., A. Hemar, D.Cefal, A. Dautry-Varsat, R. Fagard, S. Fisher,and C. Vaquero.1993. Transcriptionaland post-transcrip-tional mechanisms are involved in the absence of CD4 surface
expression in two HIV-1 chronically infected T cell lines. Int. Immunol. 5:939-947.
41. Serpente, N., M. Sitbon, and C. Vaquero. 1992. Suboptimaland optimal activation signals modulate differently theexpression of HIV-1 and cytokine genes. Biochem. Biophys. Res. Commun. 182:1172-1179.
42. Tsuchihashi, Z., and A. Kornberg. 1990.Translational frameshift
generatestheysubunitof DNApolymerase IIIholoenzyme. Proc.
Natl. Acad. Sci. USA 87:2516-2520.
43. Vickers,T.A., and D. J. Ecker.1992. Enhancement of ribosomal frameshifting by oligonucleotides targeted to the HIV gag-pol region. Nucleic AcidsRes.20:3945-3953.
44. Vlach, J., and P. M. Pitha. 1992. Herpes simplex virus type
1-mediated induction of human immunodeficiency virus type 1 provirus correlates with binding of nuclear proteinstotheNF-KB enhancer and leadersequence.J. Virol. 66:3616-3623.
45. Weiss, R. B., D. M. Dunn, M. Shuh, J. F. Atkins, and R. F. Gesteland. 1989. E. coli ribosomesre-phase on retroviral frame-shift signals at rates ranging from 2 to 50 percent. New Biol. 1:159-169.
46. Wilson, W., M. Braddock, S. E. Adams, P. D. Rathjen, S. M. Kingsman,and A.J. Kingsman. 1988. HIVexpressionstrategies: ribosomal frameshifting is directed bya short sequencein both
mammalian andyeast systems. Cell 55:1159-1169.
47. Yoshinaka, Y.,I.Katoh,T. D.Copeland, and S. Oroszlan. 1985.
Murine leukemia virus protease is encodedby the gag-pol gene
and is synthesized through suppression ofanamber termination codon. Proc. Natl. Acad. Sci. USA82:1618-1622.