0022-538X/88/072219-09$02.00/0
Copyright C 1988,American Society for Microbiology
Translational Efficiency of Poliovirus mRNA: Mapping Inhibitory
cis-Acting
Elements within the 5' Noncoding Region
JERRYPELLETIER,' GERARDO KAPLAN,2 VINCENT R. RACANIELLO,2 AND NAHUM SONENBERGl.3* DepartmentofBiochemistry' and McGillCancerCenter,3 McGill University, Montreal, Quebec, Canada H3G I Y6,
and
Department of
Microbiology,
ColumbiaUniversity,
NewYork,
New York 100322 Received17December1987/Accepted 13 March1988Poliovirus mRNA contains a long 5' noncoding region of about 750 nucleotides (theexact numbervaries
among thethree virusserotypes), whichcontainsseveralAUGcodonsupstream ofthemajorinitiator AUG. Unlike most
eucaryotic mRNAs, poliovirus
does not contain am7GpppX
(where X is any nucleotide) capstructure at its 5' end and is translated by a cap-independent mechanism. To study the manner by which poliovirusmRNAisexpressed,weexamined the translational efficiencies ofaseries of deletionmutantswithin the5' noncoding region ofthe mRNA. In thispaper wereportstrikingtranslation system-specificdifferences intheability of thealtered mRNAstobetranslated.The resultssuggesttheexistence ofaninhibitorycis-acting element(s)withinthe5'noncoding region of poliovirus(between nucleotides70 and 381) which restrictsmRNA translationinreticulocyte lysate, wheatgermextract, andXenopusoocytes, butnotinHeLacellextracts. In addition, weshow that HeLa cellextractscontainatrans-actingfactor(s) thatovercomesthisrestriction.
Thetranslation of poliovirusRNAyields a large precursor
polypeptide (NCVPOO) which is cleaved at specific sites to
generatepoliovirus proteins(for areview, seereference22). The manner by which poliovirus RNA is translated is
perplexing in several respects. First, poliovirus polysomal
mRNA, unlike the vast majority of eucaryotic mRNAs, is
not capped at its 5' end, but terminates in pUp (11, 24).
Thus, its translationmustoccurby acap-independent
mech-anism.Second, poliovirusmRNA(Lansing strain) containsa
long 5' noncoding region (745 nucleotides) which harbors
seven AUGs shortly followed by in-frame termination
co-dons (other strains ofpoliovirus contain similar cistrons in
the 5' leadersegment),whereas in most eucaryoticmRNAs the
5'-proximal
AUGistheinitiatorAUG (15).Theabilityofthese upstream AUGs to serve as translation initiation
codonshasnotbeendemonstrated. It is still unclear how the
correct initiator AUG ofpoliovirus is selected.
Translation studies in reticulocyte lysates demonstrated
that poliovirion RNA is a weak mRNA compared with
encephalomyocarditis virion (another member of the
picor-navirus family)RNA,which isalsouncapped (33). The poor
translational efficiency of poliovirion RNA and its
conse-quentinabilitytocompeteeffectivelywithothermRNAsfor components of the translational machinery were suggested (13) to be reasons why poliovirus, and not
encephalomyo-carditis virus, evolved a specific mechanism to inactivate
eIF-4F(atranslation initiation factorrequiredfor translation
ofcappedmRNAs[8, 34]). This inactivationispostulatedto
allow poliovirus mRNA to usurp the cellular translational
machinery without the need to compete
with
cellularmRNAsfor components ofthe translational machinery (for
reviews,
see references 8 and 34).Invitro studies of thetranslationalpropertiesofpoliovirus by Dorner et al. (7) revealed that in reticulocyte
lysates,
significant initiationof translation occurred in the P3 region
located in the 3' one-third of the RNA. This apparently aberrantinitiationcould besuppressedbyfactors present in HeLacellextract(7). Asimilareffectwasobserved in other studies(3, 30).Thus, reticulocyte
lysates
mightbe deficient* Correspondingauthor.
in a component(s) necessary for 5'-proximal initiation of
translation onpoliovirusmRNA (3, 7, 30).
To gain a better understanding of the mechanism of translationinitiation of poliovirusmRNA, we haveanalyzed theeffects ofdeletions in the 5' noncoding regionof polio-virus mRNA on itstranslationin severalinvitro systems,as well as inmicroinjected Xenopus oocytes. We have mapped
amajor domain(between nucleotides 70 and 381) within the
mRNA 5' noncoding region of poliovirus Lansing strain, which restricts its translation in reticulocyte lysate, wheat germ extract, and Xenopus oocytes, but not in HeLa cell extracts.Thisregioncanact inciswhenfusedto a heterol-ogous mRNA. A model is presented to explain the differ-ences in translational efficiencies observed with poliovirus mRNA in differentcell extracts.
MATERIALS ANDMETHODS
Constructionof deletiontemplates. Theconstructionof the
deletion templates and poliovirus-chloramphenicol
acetyl-transferase (Polio/CAT) fusion constructs has been
de-scribed in detail (27). The plasmid used to construct the deletionmutants(pP2-5';seeFig. 1A)wasmadebyinserting
aHindIII-SmaIfragment of poliovirustype2LansingcDNA
(17), representing bases 1 to 1872ofthe viral genome, into pSP64.
Invitrotranscriptions.Plasmidswerelinearizedwith SmaI
(for pP2-5' deletion constructs) or BamHI (for pP2/CAT
fusion constructs). Transcriptions were primed with the
dinucleotide
m7GpppG
orGpppG
(500 ,uM) toprovide
5'capped methylated(m7GpppG. .) orcapped unmethylated
(GpppG...) terminated mRNAs, respectively, and
per-formedas
previously
described (27).In vitro translations. Mahoney poliovirion and tobacco mosaic virus (TMV)RNAs were akindgift ofK. A. W.Lee. Brome mosaic virus RNA was purchased from Promega Biotec. Poliovirus mRNAs generated by in vitro transcrip-tion were 5' terminal capped methylated unless otherwise indicated. In vitro translations in micrococcal nuclease-treated rabbit reticulocyte
lysates
were performed by the method of Pelham and Jackson (26). The translationswereperformed with 12.5-,lI reaction mixtures
containing
0.2 ,ug2219
on November 10, 2019 by guest
http://jvi.asm.org/
2220 PELLETIER ET AL.
ofpP2-5' deletion mRNAs or0.5 ,ug of poliovirionorTMV RNA. Incubations took place for 60minat 30°C. Following translation, RNase T1 (200 U) and RNase A (1 ,ug) in 3 M urea-25 mM EDTA-0.5 mM phenylmethylsulfonyl fluoride (final concentration) were added, and the incubation was continued at 30°C for 1 h. A 4-,u portion of the incubation mixture was withdrawn and processed for electrophoresis. Translations in HeLa cell extracts were as described by Rose et al. (32) with modifications by Lee and Sonenberg (18). Following translations, proteins were analyzed on gradient sodium dodecyl sulfate-10%to15%polyacrylamide gels (16). Gelswerefixedin40% methanol-7.5%aceticacid, treated with En3Hance (New England Nuclear Corp.), and exposed against X-ray film at -70°C. Translation experi-mentswererepeated twice, withavariation in theresultsof less than 10%.
Translations in Xenopus oocytes. Translations inXenopus oocytes were as follows. Ten oocytes were microinjected with [3H]CTP-labeled mRNAs (50 nlperoocyte;0.2
jig/ml)
andincubatedat20°C for 15h. Theoocyteswereprocessedand theCAT activitywasdeterminedasdetailedbyColman (5). Briefly, oocytes were homogenized in 0.1 M NaCl-1% Triton X-100-1 mM phenylmethylsulfonyl fluoride-20 mM Trishydrochloride (pH 7.6) and centrifugedinanEppendorf centrifuge for 5 min, and the supernatant was used to
measureCATactivity. Before theassays, portionsof homo-genateswerecounted inascintillationcounter tonormalize for variations inmicroinjection and sample preparation.
RESULTS
Tostudythetranslational propertiesofpoliovirus mRNA, we used RNA transcribed from a type II (Lansing strain)
polioviruscDNA clone withSP6 RNApolymerase. A trun-catedcDNAclone (pP2-5')wasusedtoavoid theanalysisof many translation products due to protein processing. This cloneextendstonucleotide 1872 in the VP3regionandgives risetoasingle 1.8-kilobaseRNAspecies following transcrip-tion in the SP6 system and analysis on formaldehyde gels (datanotshown). Deletionsextendinginto thepoliovirus5' end were constructed (Fig. 1A), and translation of the
mRNAswasexamined in several translationsystems: rabbit reticulocyte lysate; extracts from HeLacells, wheat germ, and mouse L cells; and Xenopus oocytes (by microinjec-tion). Becausepoliovirus genomicandpolysomalRNAsare uncapped (9, 11, 19, 24) and translate byacap-independent
mechanism (reviewedin references 8 and34),weused both 5' terminal capped unmethylated (GpppG... .) and capped methylated (m7GpppG. . .)mRNAs forourtranslation
stud-ies. The capped unmethylated mRNAs are functionally analogous touncapped mRNAs,since it is themethylgroup
oncapped methylated mRNAs whichplaysapivotalrolein
the function of the cap structure (2). We used capped unmethylated mRNAs rather than uncapped mRNAs
be-cause theformer are considerably more stable (and are as
stableas theirmethylated counterparts) in cell-free transla-tion systems (10).
Translation of pP2-5' deletion mRNAs in reticulocyte ly-sates.Translationswereinitially performedinareticulocyte lysate toassess the effect ofa methylatedversus
unmethy-lated cap structure on the expression of pP2-5' mRNA. S-Adenosyl homocysteine was included in the translation
incubations to preventmethylation of unmethylated capped transcripts by endogenous methyltransferases (2). Transla-tion products were resolved by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and are shown in Fig.
2A.Translation of
poliovirion
RNAwasabout five times lessefficientthan thatof TMV RNA
(compare
lane 5with lane3;determined
by [35S]methionine
incorporation,
since theme-thioninecontentsofboth
proteins
weresimilar)
andwasnotinhibited
by
the capanalog
m7GDP(compare
lane 6 with lane5),
in contrast to the translation ofTMV RNA(85%
inhibition;
compare lane 4 with lane3).
Translation ofSP6-derived
methylated
andunmethylated
herpessimplex
virus type 1
thymidine
kinase mRNA(pX1)
wasperformed
as acontrolto assessthe cap
dependency
ofthetranslation system, since thetranslation of this mRNA haspreviously
been shown to be cap stimulated
(28).
Translation ofun-methylated
thymidine
kinase mRNAwasnotaffectedby
theadditionofm7GDP
(compare
lane 8 with lane7),
whereas thepresence ofa
methylated
cap structure stimulatedtransla-tion ofthe mRNAabout sixfold
(compare
lane 9 with lane7).
Addition of m7GDP
(lane 10)
reduced thetranslationof themRNA tothe levelobserved with the
unmethylated
thymi-dine kinase mRNA (lanes 7 and
8). Thus,
asexpected,
the translation ofcap-dependent
mRNAs in ourreticulocyte
lysate
is stimulatedby
the presence of amethylated
cap structure. Translation of thepP2-5'
transcripts
was veryinefficient and cap
independent
in thereticulocyte
lysate
(lanes
11 to14),
as was observed with the translation ofpoliovirion
RNA in thereticulocyte lysate (lane 5) (33).
Since the
protein product
is not visible atthis exposure, alonger
exposure is shown(lanes
19 to22).
Translation of bothunmethylated
andmethylated
mRNAswas similarandslightly
stimulated(1.5-fold) by
theaddition ofm7GDP(lanes
20 and
22).
Suchastimulation hasbeenpreviously
observed withnaturally
uncapped
mRNAs(35)
and can also be noticed withpoliovirion
RNA(compare
lane6with lane5).
Taken
together,
these resultsindicatethatpoliovirus
mRNA translationisindifferenttothenatureof thestructurepresent at its 5' terminus. Theprotein product
synthesized
from pP2-5' mRNA was confirmed to be a truncatedpoliovirus
product
containing
VP4 since it could beimmunoprecipi-tated
by using
an anti-VP4antibody (data
notshown).
Wenote that translation of
poliovirion
RNA(lanes
5 and6)
ismuch better than that of
pP2-5'
mRNA(lanes
11 to14).
Aplausible explanation
forthis difference isthat most ofthetranslation
products
frompoliovirion
RNAresultfrominter-nalinitiationin the P3
region
(compare
with reference7);
thisregion
islacking
inpP2-5' mRNA,
which can thereforetranslate
only by
5'-proximal
initiation.We wished to determine whether the low translational
efficiency
ofpoliovirus
mRNA inreticulocyte lysates
wascaused
by
sequences in the 5'noncoding region.
Wethere-fore
assayed
a series ofmutantpP2-5'
mRNAscontaining
deletions in the 5'
noncoding
region
andconcomitantly
assessed the effect of
methylated
versusunmethylated
capped
mRNAsonexpression.
TheresultsareshowninFig.
2B:
A5'-33
(lanes
6 and7),
A5'-80
(lanes
8 and9),
A5'-96
(lanes
10 and11),
and A5'-140(lanes
12 and13)
translatedpoorly
in thereticulocyte lysate, and,
likepP2-5'
mRNA(lanes
4 and5),
translated in anon-cap-stimulated
fashion(densitometry
scanning
ofthese lanes wasperformed
on alonger
exposureofFig.
2).
ThetranslationofA5'-320,
albeitpoor,
revealed that themethylated
mRNA wasexpressed
slightly
better(1.5-fold)
than itsunmethylated
counterpart(compare
lane 15 with14).
FormutantsA5'-632
andA5'-733,
translation of
unmethylated
mRNAs wasremarkably
moreefficient
(ca.
20-to35-fold)
than that ofpP2-5'
mRNA(lanes
16 to19).
Translation ofmethylated
A5'-632 andA5'-733
mRNAs was ca. threefold more efficientthan that oftheir
unmethylated
counterparts(compare
lane 17 with lane 16 J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
cis-ACTING ELEMENTS IN POLIOVIRUS mRNA
A 1:
FRAME 2: C . mD
FRAME 3:
F4- HHindm
pP2-5 As 7
66
A5-33 ,(-<
33
,&5'-80 l=4
80
&5-96 r ) 96
A5'140 Cr (
_--
BalI ATG745HaeEBsm I EcoFBamVH Smna
380 457 70 8568 1872
RETIC LYSATE
GQDDoG Mmpa
1.0 1.1
1.5 1.6
1.6 1.4
2.3 2.2
2.8 3.5
320
632
733
Y Y
631 733
m8m mThTThn
38 733
CATFUSION MIITANTS
ATG BunHI
X ;..-.¢.
1
l2e
,r-w*v,&w SQ67i;57:sf
pCAT Cz
RETIC L CELL W&TGERM
256 07 44 300
10 1D ID 1.D
1.0
7 07.
320
46
63 631
47 1.0
0L7
105 2.0 12.2
25 12
320
461..
, 32 46FIG. 1. Maps of poliovirus deletionmutants.(A) Structure of deletionconstructsofpoliovirusRNA. Potentialopenreadingframes in the 5'noncoding regionareshownatthetopof thediagram. V,AUGpositions. The cleavage site of restrictionenzymesusedtoproducethe
deletionmutantsareshownonthepP2-5' template. The numbers shownrepresentdistances,innucleotides,from the firstnucleotide of the 5'end of the viraltemplate. Symbols: oi,SP6polymerasepromoter;-,24nonviral nucleotides betweentheSP6promoterand the first
nucleotide of the viralgenome; _, P2/Lansing viralsequences;deletedsequences arebracketed. RNAstranscribedfrom thesetemplates
containthe initiator ATG (shown at nucleotide 745) of the viral polyprotein encoding anopen readingframe foratruncated poliovirus
polyprotein consisting of VP4 (69 amino acids), VP2 (271 amino acids), and 37 amino acids of VP3 (predicted total molecular weight, 42,000 [17]). Deletions thatcausealterations withinupstreamopenreading frames, resulting in termination within the major polioviruspolyprotein openreading frame,areindicated abovetheconstructsA3'-631 and A3'-381. Therelativetranslational efficiencies of the deletionconstructs
obtained inreticulocyte lysatesareindicatedtotheright. Radioactivebandscorrespondingtopoliovirus protein productswerequantified by
soft-laserdensitometry (LKB Instruments, Inc.), andthe value obtained forcapped unmethylated pP2-5'mRNAwassetas1.0.(B) Diagram
of fusionconstructsbetweenpoliovirusdeletionmutantsand CATcodingsequence.Symbols: =z,CATgene; _,sequencesderived from
pP2-5'. Alterationsinopenreadingframescausedbydeletionmutagenesisaredrawnabove theappropriateconstruct.Theasterisksabove
A5'-465/CAT denote the positions of synthetic HindIll linkers. The relative translational efficiencies obtained with the pP2/CAT fusion
constructsareindicated totheright. Thevalue obtained forpP2/CATwassetas1.0.
VOL. 62, 1988 2221
&5 320 EJ(
&5'632 =(
5-'733 l K(
A3-631 = 4
A3 381 C3-4
B
3.6 5.3
34 110
20 66
1.2 1.1
1.1 1.5
pP2/CAT OZ3-4
AS533/CAT D(
A5140tAT D(
O5'-320tICAT ,(
&5:465/CAT =D(
a-631/CAT OH
£3- 461/CAT
3'?70/CAT
70 a-320/&3r461/CAT [{
.,At F.=-l 11
=4
_
v
.,C,gxzm
140
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.130.493.66.558.2]2222 PELLETIER ET AL.
-RNA TMV POLIO -X2_5' A5-733
r_ 1--'' .m 2E2'
1 2 3 4 576 8 9 1011 12 13 14 15 1617 18 kOal
93-
n--
46-
a-
30-
20-GpppGO. + * + +
m7upppG... + + + + +
mr7GDP - - + - + - + - + - * -+ - + - +
pXlP2-5 4!5 -80
5_r_-kDa I r23 4 5 6 7 8 9 10 11
93-46- WO
30-12 13 14 15 16 17 18 19 20 21 22 23
_e
_ I_
_
--
20-GpppG.:..
[image:4.612.159.467.68.390.2]m7GpppG. .+ * 4
FIG. 2. Translation in rabbitreticulocyte lysates programmed with pP2-5' poliovirusdeletion mRNAs. Valuesobtainedafterscanningthe
autoradiograph relativetotheamountofprotein synthesized from pP2-5' mRNAareindicatedinFig. 1A. Kinetic analysisanddose-response
curvesindicated thattherates of translation werein the linearrange. (A) Effect ofcapanalogontranslation inreticulocyte lysates.The
followingamountsof mRNAswereadded: TMVandpoliovirionRNA,0.5,ug; thymidinekinase 0.2p.g;pP2-5',0.2Rg;A5'-733,0.2,ug.The
protein products in lanes 11to14weredifficulttovisualize, andtheautoradiographafterbeing exposedfive timeslongeris shown(lanes 19
to22). m7GDP (0.6 mM)wasadded where indicated. Lanes containing unmethylated (GpppG....) andmethylated(m7GpppG. .)capped mRNAsareindicated.(B) Translation of pP2-5' poliovirus deletionmRNAs.Translations(in12.5RIl)wereperformedwith0.25 p.gof mRNA.
Molecularmassesinkilodaltons (kDal)areshownontheleft of eachpanel. and lane 19 with lane 18). This translation was cap stimu-lated, since translation of methylated A5'-733 mRNA was inhibited by m7GDP, whereas itsunmethylated counterpart was not (Fig. 2A, lanes 15 to 18). The translation oftwo mRNAs, A3'-631 and A3'-381, in which distal sequences
within the 5' noncoding region were removed, was very inefficient(Fig. 2B, lanes 20to23). Thereduced translation obtainedwith these mRNAs isnotduetothepresence ofa new upstream cistron which now overlaps the majoropen reading frame of VP4-VP2-VP3 (diagrammed in Fig. 1A). Polypeptides ofca. 17 and ca. 12 kilodaltons areexpected
from the new cistrons in A3'-631 andA3'-381, respectively. Theseproteins, whichareexpectedtocontain six (A3'-631) andfive(A3'-381) methionines, werenotobservedfollowing translation of these mRNAs (Fig. 2B), indicating that the newcistronsarenotused.These data indicate thatsequence
elements upstream ofnucleotide 381 formpartofadomain which contributes tothe reduced translational efficiency of poliovirus mRNA in areticulocyte system.
Translation of pP2/CAT deletion mRNAs in reticulocyte lysate.To determine whethertheinhibitory effecton trans-lation observed with the poliovirus mRNA 5' noncoding region could beimpartedto an mRNA whose translation is
very efficient inreticulocyte lysates, wejoined the 5'
non-coding region of the deletion mutantsto the bacterial CAT coding region (Fig. 1B). Translations of capped unmethyl-atedversuscapped methylatedmRNAswereperformedfor
all the experiments described below, but only the results with capped methylated mRNAs are presented, since the
data obtained with thetwotypes of mRNAsweresimilarto those obtained in Fig. 2. The results of translating the chimeric deletion mRNAs inareticulocyte lysateareshown
inFig. 3,and the dataaresummarized inFig. 1B.Asshown above (Fig. 2), the translation of poliovirion RNA was inefficient(lane 2)ascomparedwiththetranslation of TMV (Fig. 3,lane3)and brome mosaic virus(lane4)RNAs.pCAT mRNAwastranslatedveryefficiently (lane 5).Fusion ofthe 5' noncoding region of poliovirusmRNA (nucleotides 1 to 733)to theCAT coding sequence drastically reduced CAT
protein synthesis (ca.250-foldreduction;lane6). Adeletion of 33 nucleotides from the 5' end did not relieve the inhibitory effect of the poliovirus 5' noncoding sequences
(lane 7). However, further deletions gradually led to
in-creases in the translational efficiency ofpP2/CAT mRNA. Translation was stimulated 3.7-fold for deletion mutant A5'-140/CAT (compare lane 8 with lane 6), 6.8-fold for A5'-320/CAT (compare lane 9 with lane 6), and 47-fold for A5'-465/CAT (compare lane 10 with lane 6) relative to
1P2-0 1920C21 22
J. VIROL.
wF *
_ * _ F
4 +
on November 10, 2019 by guest
http://jvi.asm.org/
<_CO
68-~~~~~~~~~!-~
C)
<.
460
~-
> ><~
-kDal
46-93- Ij .1 ..
F-~~~~~~~~~~~~C
68- C,,DC
46-
30-0
a0 VW -O - .m-'--CAT _-woo*~ -CAT
20-X 2 3 4 5 6 7 8 9 10 11 12 13 1.4
FIG. 3. Translation in rabbit reticulocyte lysates of pP2/CAT deletion mRNAs. Translations were performed as described in Materials andMethods. Values obtained by scanning of the
autora-diograph relative to the amount of protein synthesized from pP2/CAT mRNA (setas 1.0)areshown in Fig. 1B. Theamountsof
mRNA used was asfollows: poliovirion, 0.25 ,ug; TMV, 0.25 ,ug;
bromemosaic virus(BMV), 0.25 ,ug;pP2/CAT, 0.25 ,g. Molecular
massesin kilodaltons (kDal)areshownontheleft.
pP2/CAT. The inhibition obtained with A5'-465/CAT was only 5-fold, compared withthe 250-fold inhibition obtained with thefull-length poliovirus 5' noncoding region. Transla-tion ofboth A3'-631/CAT and A3'-461/CAT mRNAs was as inefficientastranslation of mRNA containing the full-length
5'noncodingsequence(lanes 11and12), consistent with the previous conclusion that the major inhibitory sequence is upstream of nucleotide 465. When alarger deletion mutant
(A3'-70) was tested, translation was vastly increased and
approached the efficiency exhibited by pCAT mRNA
(com-pare lane 13 withlane 5). These results areconsistent with
thoseofFig. 2andmapthetranslational inhibitory element to sequences between nucleotides 70 and 461. A fragment containing nucleotides 320 to 461 was fused to the CAT coding sequence and tested for its ability to translate in a reticulocytelysate. Translation of thismRNAwasinhibited 10-foldmorethan that of CAT mRNA(comparelane 14 with
lane5). Thisinhibition, althoughsignificant, cannot account for the largedecrease in translation(250-fold) caused bythe full-length 5' noncoding region ofpoliovirus mRNA when fused to CAT mRNA. Therefore, sequences upstream of nucleotide 320arerequiredfor theinhibition.Since deletion
ofsequencesdownstream ofnucleotide 381 didnotmitigate the inhibition (Fig. 2), we conclude that a domain exists
between nucleotides 70 and 381 which prevents efficient translation ofpoliovirus mRNA inareticulocyte lysate.
Translation ofpP2/CAT deletion mRNAs in extractsfrom HeLa cells. Translations in HeLa cell extracts were per-formed withcapped methylated deletionmRNAs ofpP2-5' andpP2/CAT.' The results obtained were similar with both setsofconstructs, andonly thoseforpP2/CAT mRNAsare
shown (Fig. 4; summarized in Fig. 1B). Translation of poliovirion RNA (Fig. 4, lane 2) was as efficient asthat of TMVRNA(lane 3) in HeLaextracts. Theefficient transla-tion ofpoliovirionRNA in HeLacellextracts isincontrast toits inefficienttranslation inreticulocyte lysate (Fig. 2and 3). Juxtaposition of the poliovirus 5' noncoding region in
1 2 3 4 5 6 7 8 9 10 11
FIG. 4. Translation of pP2/CAT deletion mRNAs in mock-in-fected HeLa cell extracts. Translations were performed, as de-scribed inMaterialsandMethods,with 0.4 ,ugof cappedmethylated pP2/CAT deletion mRNAs and 0.6
jig
of poliovirion and TMV RNAs, and the products wereresolved ongels and processed as described in Materials andMethods. Molecularmasses in kilodal-tons(kDal) areshown on theleft.front oftheCATmRNAslightly increased translation
com-pared with that ofpCAT mRNA (1.5-fold; compare lane 5
with lane 4), in sharp contrast to the inhibitory effect observed in the reticulocyte lysate (Fig. 3). Stepwise dele-tions originating from the 5' end of the mRNA had no
significant effect on the translational efficiency of the
mRNAs(lanes6to8). Similarly, removal of102nucleotides fromthe 3' end ofthe noncoding region (nucleotides 631to 733) had nosignificant effect ontranslation (1.3-fold higher than thatofpP2/CAT mRNA; lane 9). When mostofthe 5'
noncoding region was removed by a 3' deletion up to
nucleotide70,translationwasagainstimulatedtwofoldover
that of pP2/CAT mRNA (compare lane 10 with lane 4).
Fusion of thefragmentcontaining nucleotides 320to 461 to
CAT (A5'-320/A&3'-461/CAT) did not affect CAT translation
(lane 11), in contrast to theinhibition obtainedin the rabbit
reticulocyte lysate (Fig. 3). These results demonstrate that
the cis-inhibitory elements present in the 5' noncoding
region of poliovirus mRNA do not function in HeLa cell
extracts andare consistent withthe findingthatpoliovirion RNAtranslatesefficientlyin HeLa cellextracts(Fig.4, lane 2).
TranslationofpP2/CAT deletion mRNAs in other transla-tion systems. The results presented so far point to striking translational differences among the various 5' deletion mu-tants in reticulocyte
lysates
and HeLa cell extracts.Conse-quently, it was of interest to determine whether these
differences occurbecause the reticulocyte lysate ispeculiar with respect to poliovirus RNA translation. We used two additional invitrotranslation systems: extractsfrommouse Lcells(afibroblastictumorcellline) andplantwheat germ.
The translation results obtained with these systems are
summarized in Fig. 1B. Translation of pCAT in L-cell
extracts was reduced
only
4.4-fold when the 5'noncoding
region ofpoliovirus mRNA was
joined
to the CATcoding
region. Somewhat higher
(12-fold)
translationalefficiency
than thatofpP2/CAT mRNAwas obtained with theA3'-70
deletion construct mRNA. Deletion ofthe
5'-proximal
320on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.296.70.267.2] [image:5.612.331.546.71.263.2]2224 PELLETIER ET AL.
H)
C'I) C\j
< 11-. (O rl-~~~~N C')
Z t4 e\ 1 .1
cE 0 O- C'I) ~ c' Lo
T- C C '1 41
U.
3-AcC- _
1-AcC-A'm.4E
c-o-
e
e
1 2 3 4 5 6
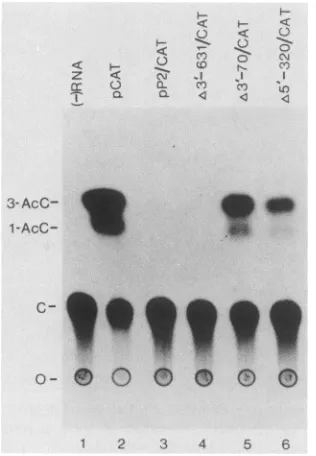
FIG. 5. Translation of pP2/CAT deletion mRNAs in Xenopus
oocytes. Kinetic analysis and concentration curves indicated that
the observed CAT activity for all constructs was in the linear
response range. The positions ofchloramphenicol(C) and
acetyla-tedforms ofchloramphenicol(AcC)areindicated. 0,Origin.
nucleotides of poliovirus mRNA 5' noncoding region (A5'-320) resulted in increased translational efficiency (8.4-fold higher than that of pCAT). Thus, the behavior of the 5' noncoding region of poliovirus mRNA in L-cell extracts resembles the situation in HeLa cell extracts more closely, since it exerts a relatively weak (as compared with the
reticulocyte lysate) cis-inhibitory effecton CAT translation
in L-cellextracts.
A strikingly differentpattern from thatobserved in L-cell
extracts emerged when translation was conducted in wheat
germextract(results summarized in Fig. 1B). Fusion of the poliovirus mRNA 5' noncoding region to the CAT mRNA causedadramatic decrease in translation relativetopCAT (a
ca. 300-fold difference). Similarly, the deletion mutant A3'-631/CAT translated inefficiently. Thus, the results from the wheat germ translation system indicate that this system resembles the reticulocyte lysate with respect to poliovirus translation and is considerably different from HeLa and L-cellextracts.
Inadditiontotheinvitro studies, wewishedtoassess the translational properties ofthe mutant mRNAs in vivo. To this end, we used the Xenopus oocyte system, in which translation of pCAT mRNA was determined by analysis of
CAT activity after injection of mRNA. Extracts from
oocytes not injected with mRNA showed no CAT activity
(Fig. 5, lane 1), whereas those injected with pCAT mRNA exhibited CAT activity (77% ofchloramphenicol converted
to the acetylated form; lane 2). Fusion of the poliovirus mRNA 5' noncoding region to the CAT coding sequence
completely abolished CAT activity (lanes 3 and 4). A 3' deletion extendingto nucleotide70 (/&3'-70/CAT)resultedin appreciable CAT activity relative to that obtained with pCAT mRNA (twofold less than that of pCAT mRNA; compare lane 5 with lane 2), whereas injection of mRNA witha5' deletionextendingtoposition 320 resulted in some
CAT activity (16-fold less than that ofpCAT mRNA;
com-parelane6withlane2). These resultsdemonstratethat the
translational
machinery
in Xenopus oocytes moreclosely
resembles thereticulocyte lysate translation systemthan the HeLacellextractwithrespecttotheeffectsof thepoliovirus
mRNA5' noncodingregion ontranslation.
Possible translational trans-activator in HeLa cells. The
differential effects of the poliovirus mRNA 5' noncoding
elements on translation in various cell extracts and in Xenopus oocytescould becausedbytrans-acting repressors
or activators present in various amounts in the different
translation systems. As an initial step in analyzing these
possibilities, we examined the effects of adding ribosomal
high-salt washfractions (referredto asinitiationfactor [IF]
preparations, since these fractions are enriched in IF) from
reticulocytes and mock- andpoliovirus-infected HeLa cells
on the translation of pP2/CAT fusion mRNAs in rabbit
reticulocyte lysates. When the fractions were added to micrococcalnuclease-treatedreticulocyte lysate without ex-ogenous mRNA, no new protein products were observed (Fig. 6,lanes 2 to 4), indicating that no translationally active RNAcopurifiedwith the IF. The addition of reticulocyte IF had no effect on the translation of pCAT mRNA (compare lane 6with lane 5), whereas addition of IFs from mock- and poliovirus-infected HeLa cells caused a considerable de-crease in pCAT mRNA translation (ca. 10-fold; compare lanes 7 and 8 with lane 5). The reasons for inhibition of pCAT mRNA translation by IFs from mock-infected HeLa cells are not clear, although a similar effect has been previ-ously observed (4). Translation of pP2/CAT mRNA was very inefficient (lane 9), as shown in Fig. 3, and IFs from the reticulocyte lysate did not stimulate translation of this mRNA (Fig. 6, lane 10). However, addition of IFs either from mock-infected (lane 11) or from poliovirus-infected (lane 12) HeLa cells stimulated translation of pP2/CAT mRNA to thelevel observed when HeLa IFs were added to pCAT mRNA translation (lane 7). We also tested the dele-tion mutant A3'-70/CAT and found that it translated with
similar characteristics to pCAT mRNA: expression of this
mRNA was not stimulated by IF from reticulocytes (com-pare lane 14 with lane 13), but was inhibited by IF from mock-infected HeLa extracts (lane 15) and further inhibited by IF from poliovirus-infected HeLa extracts (lane 16). These results suggest that HeLa cells contain a trans-acting factor that facilitates efficient translation of poliovirus mRNA.
DISCUSSION
We have mapped an element(s) in the 5' noncodingregion of poliovirus mRNA that exhibit a strongcis-inhibitoryeffect on translation inreticulocytelysate, wheat germ extract, and Xenopus oocytes. Thiselement(s), as deducedfrom transla-tion of a series of deletransla-tion mRNAs in a reticulocyte lysate, appears to reside between nucleotides 70 and 381(Fig. 2and 3). A fragment containing nucleotides 320 to 461 inhibited translation 10-fold whenfused to CAT mRNA (Fig. 3). This inhibition is 25-fold lower than that causedbythefull-length 5' noncoding region. However, deletion of nucleotides 1 to 320resulted in a35-fold-higher inhibition of CATtranslation (Fig. 3). A further deletion extending to nucleotide 465 (A&5'-465/CAT; Fig. 3) inhibited CAT expression only five-fold. These results suggest that there may be two or more inhibitory elements between nucleotides 70 and 381 which operate synergistically and which have been bisected by some of the 5' deletions reported here. Alternatively, it is possible thatnucleotides 70 to 381 contain onecis-inhibitory
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.105.263.68.296.2]cis-ACTING ELEMENTS IN POLIOVIRUS mRNA 2225
r--RNA
r
---pCAT
- l pP2/CATir--3-70/CAT---1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
kDal
93-
68-
46-
30-
20-Retic IF:- + _- _ + _ _- +
Mock-infected IF: - - + + _ _ _ + _ _ _ +
Infected IF: - - - + + _ + - +
FIG. 6. Effect of IF addition on translation of pP2/CAT mRNAs in areticulocytelysate.Reactionmixtures (13 ,ul) were programmed with 0.2 ,ugofpCAT, pP2/CAT, orA3'-70/CAT mRNA. Reaction mixtures contained 3 ,ulofIF(ca. 31 mg/ml)from reticulocytes or mock- or poliovirus-infected HeLa cells whereindicated. After incubation at 30°C for 60 min, samples of4,ul were processed for gel electrophoresis. Thegelswereprocessed for autoradiography as described in Materials and Methods. Molecular masses inkilodaltons(kDal)are shown on the left.
element.Thesepossibilities are also supported by the oocyte
microinjectiondata(Fig. 5),which show thatdeletionof the
5'-proximal 320 nucleotides partially relieved the
transla-tional inhibitory effectexerted by the 5' noncoding region.
Additional work is necessary to distinguish between these
twopossibilities.
Itis unlikelythat thetranslationalcis-inhibitory effects of
the poliovirus mRNA 5' noncoding region are caused by
upstream AUGs, which would act to impede the 5'-to-3' movement ofribosomes toward the initiator AUG745 and
thus reducetranslationalinitiation(15).If thiswere thecase,
then removal of upstream AUGs would be expected to
stimulate translation of poliovirus mRNA. We tested this
hypothesis by systematically mutating the 5' upstream
AUGs to UUGs and did not find such an effect in the
reticulocyte
lysate
(J. Pelletier, V. R. Racaniello, and N.Sonenberg, unpublished observations). A similar case has
been reported by Katz et al. (14), in which deletion of an
upstream AUG had no effect on the translation of avian
retrovirusmRNA. Atpresent, a simpleinverserelationship
betweenthe existence ofupstreamAUGs in the 5'
noncod-ing region of poliovirus mRNA and its translational
effi-ciency does not exist.
The restrictions imposed by the 5' noncoding region of
poliovirus mRNA on translation in a reticulocyte lysate
might be relevanttotheunusual
pattern
ofproteinssynthe-sized inthe reticulocyte lysateas described by Brown and
Ehrenfeld(3)andDorner et al.(7). Dorner et al. showed that
inareticulocyte lysateasignificant proportion of translation
initiation originates in the P3 region located in the 3'
one-third of the RNA (7). Evidence from electron microscopy
studies showing ribosome bindingto the mRNA P3 region support thisconclusion (20).Itispossiblethatinreticulocyte
lysates, in which initiation of translation at the 5' end of
poliovirus
mRNA isweak,aninternalinitiation siteneartheP3 region becomes available for ribosome binding. Under
conditions when 5' end initiation is efficient
(in
HeLaextracts), one might not expect to see initiation at the P3
region owing tocompetition. Inagreement with these
obser-vations, Phillips and Emmert (30) presented evidence that
reticulocyte lysates are limiting in a factor required for
utilizationof the 5'major initiator AUG.Supplementationof
areticulocyte lysatewith HeLacytoplasmicextract resulted
in aqualitative improvementinthe selection ofthe correct
initiationsite (3, 7, 30). Brown and Ehrenfeld also reported
astimulatory effect ofaribosomal saltwashfromHeLa cells
ontranslation of poliovirusRNAinareticulocyte lysate (3).
However, a similar stimulation was also observed when a
ribosomal salt washfrom reticulocytes was added (3). The
activitywe described here is specific for HeLacells, since
addition of a ribosomalhigh-saltwashfrom thereticulocyte had noeffectontranslation ofpP2/CATmRNA(Fig.6, lane 10), whereas HeLa fractions from mock- and poliovirus-infected cells had a significant stimulatory effect (Fig. 6, lanes 11 and 12).Thus,theactivity in HeLa cells described
here is apparently different from that describedby Brown
and Ehrenfeld(3).
Whatmight be the nature of the translational restrictive
elements? We believe that it is likely to be secondary structure, since such elements have been shown to inhibit translation(28). Itis of interest thatcomputer-aided
model-ing ofthesecondarystructureofpoliovirusmRNArevealed
extensive secondary structure in the 5' noncoding region
that is conservedin the threepoliovirusserotypes(J. Maizel,
personal communication). It is possible that the secondary
structure interdicts the translation of
poliovirus
mRNA inreticulocyte lysates, wheat germ extracts, and Xenopus
oocytes. We haverecently shown(27)thatthepoliovirus5'
noncoding region has a sequence element necessary for
cap-independent translation. Our datasuggest that
reticulo-cyte lysates are limiting in a
factor(s)
necessary forcap-independent translation. This would explain the low
trans-lationalefficiency of
pP2/CAT
inreticulocyte
lysates. Inthisscenerio, ribosomes would be inhibited from
scanning
thepoliovirus
leaderregion by
secondary
structure betweennucleotides 70 and 380. In HeLa extracts,
however,
ribo-VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.142.474.68.284.2]2226 PELLETIER ET AL.
somes would presumably bypass this barrier by binding internally to the 5' noncoding region (29). Consistent with this idea, we have recently shown both in vivo and in vitro that ribosomes can initiate internally on the 5' leader of poliovirus mRNA (J. Pelletier and N.
Sonenberg,
submitted for publication).The possible existence of trans-actingfactors that affect the translation of poliovirus mRNA in a cell type-specific manner is novel and of interest in several respects. The differential effects on translation exhibited bythepoliovirus mRNA 5' noncoding sequences can be explained either by the existence of negative regulatory trans-acting factors in reticulocytes, wheat germ extracts, and Xenopus oocytes or, alternatively, by the presence ofpositive regulatory trans-actingfactors in HeLa and L cells. Our data(Fig. 6) support the second possibility, since ribosomal high-salt wash frac-tions from HeLa cells (both mock andpoliovirus infected) relieve thecis-inhibition imposed by thepoliovirus mRNA 5' noncoding region in reticulocyte lysates. It is important to emphasize that we have excluded the possibility that the cell-specific translational effects areattributed todifferences in the in vitro translation conditions, since similar results were obtained under different salt concentrations and tem-peratures (J. Pelletier, unpublished observations).
What is the nature of the translational trans-activator? One possibility is that an initiation factor thatisrequired in larger amounts for the translation of poliovirus mRNA is moreabundant in HeLa and L cells than in the specialized cells we used here. Infact, such a property was attributed to eIF-4A by Daniels-McQueen et al. (6), who concluded that higher levels ofeIF-4A are required for poliovirus mRNA thanforencephalomyocarditis virus mRNAtranslation. This is aninteresting possibility, because eIF-4A was suggested to possessRNA-unwinding activity (31), and the restriction ofpoliovirus mRNAtranslation couldbeattributedto ahigh degree of secondary structure. There are several reasons, however, why we do not believe that the low levels of
translation of poliovirus mRNA in
reticulocyte
lysates,wheat germextracts, or Xenopus oocytes can be attributed tolimiting levels of eIF-4A per se. The addition of purified eIF-4A had no effect on the translation ofpP2/CAT mRNA in reticulocyte lysate (Pelletier, unpublished observations). Addition of IF preparations from reticulocytes (which are a
very rich source of eIF-4A) could not relieve the
cis-inhibition imposed by thepoliovirus 5' noncoding region on pCAT mRNA translation, whereas IF preparations from HeLacells partiallyrelieved this inhibition(Fig.6). Further-more, it was shown by Blair et al. (1) that eIF-4A had no
stimulatory effect onthetranslation ofpoliovirus mRNA in
an ascites-reconstituted translation system, whereas the translation of encephalomyocarditis virus mRNA was stim-ulated. Because of these reservations, an alternative possi-bility is that HeLa cells contain a factor that facilitates cap-independent and internal binding ofribosomes to the 5' noncodingregion of poliovirus mRNA and thatthisfactor is limiting in reticulocyte lysates, wheat germ extracts, and Xenopus oocytes. As mentioned above, translational initia-tion in these extracts probably occurs by a 5'-end-mediated eventand isrestricted by a domain found between nucleo-tides 70 and 381 (presumably in the form of secondary structure).
Thepossibility that poliovirus mRNA translatesefficiently only in certain cell types is consistent with recent results showing that translation of poliovirus RNA in U-937 (a hystiocytic cell line that contains an equal number of polio-virus receptors to that in HeLa cells [23]) is considerably
lower than that in HeLacells (L. Carrasco, personal
com-munication). In addition, poliovirus infection of several
established human blood cell lines showed that viral
repli-cation differeddependingonthedifferentiationstageand cell
lineage (25). Inthose studies, the stagein the viral
replica-tion cycle at which the restriction occurred was not deter-mined, but our results would suggest that a block at the
translational levelisaclearpossibility. Thus, thetranslation
restriction phenomenon may be related to the restricted
tissuetropismexhibited by poliovirus
(12,
25). Althoughit isbelievedthatpoliovirus tissuetropismisdeterminedlargely
bythe distribution ofcell-specific virus receptors (12), it is
possible that additional restrictions are imposed by the
translationalmachinery of agivencell. Our study raises the interesting possibility that cis-acting sequences similar to those ofpoliovirus also existin cellular mRNAs and deter-minetissue specific translation. Suchelements could play a key role in development anddifferentiation.
Recently,Nicklinetal.(21) reported that deletion of the 5' noncoding region ofpoliovirus mRNA resulted in a marked increase in translation efficiency in a rabbit reticulocyte
lysate,
in agreement with our results. Together, these datapoint to the existence of a cis-inhibitory element(s) within
the 5' noncodingregion of poliovirus RNA. Thiselement(s) restricts mRNA translation inreticulocyte lysates, Xenopus oocytes, and wheat germ extracts, but not in HeLa cell extracts.
ACKNOWLEDGMENTS
We thankAnthoula Lazaris for excellenttechnical assistance and Isaac Edery for critical comments on the manuscript. We are grateful to EckardWimmer for his gift of anti-VP4antibody.
This research was supported by a grant from the Medical Re-search Council ofCanada to N.S. and by Public Health Service grantAI-20017toV.R.R. fromthe National Institute ofAllergyand Infectious Diseases. N.S. is the recipient of a Medi,cal Research Council scientist award from the Medical Research Council of Canada. V.R.R. is the recipient ofa SearleScholar Award andan I. T. Hirschl Career Scientist Award. J.P. was a recipient of a
predoctoral studentship from the Medical Research Council of Canada.
LITERATURE CITED
1. Blair,G. E.,H. H. M. Dahl,E. Truelsen, and J. C. Lelong. 1977. Functionalidentity of a mouse ascites andarabbit reticulocyte initiationfactorrequired for naturalmRNAtranslation. Nature (London)265:651-653.
2. Both, G. W., A. K. Banerjee, and A. J. Shatkin. 1975. Methyl-ation-dependent translation ofviral messenger RNAs in vitro. Proc. Natl. Acad. Sci. USA72:1189-1193.
3. Brown, B. A., and E.Ehrenfeld. 1979. Translation of poliovirus RNAin vitro:changesincleavagepatternandinitiation sitesby ribosomal salt wash. Virology 97:396-405.
4. Brown, B. A., and E.Ehrenfeld. 1980. Initiation factor prepara-tions from poliovirus-infected cells restrict translation in retic-ulocytelysates. Virology 103:327-329.
5. Colman, A. 1984. Translation ofeukaryotic messengerRNAin Xenopus oocytes, p. 271-302.InB. D. Hames andS.J.Higgins (ed.),Transcription and translation: a practical approach. IRL Press Ltd.,Oxford.
6. Daniels-McQueen, S., B. M.Detjen, J. A. Grifo,W. C. Merrick, and R. E.Thach. 1983. Unusualrequirementsforoptimal trans-lationof polioviral RNA in vitro. J.Biol. Chem.258:7195-7199. 7. Dorner, A. J., B. L. Semler, R. J. Jackson, R. Hanecak, E. Duprey,andE. Wimmer. 1984. Invitro translationofpoliovirus RNA:utilization of internal initiation sites inreticulocytelysate. J.Virol. 50:507-514.
8. Ehrenfeld, E. 1984. Picornavirus inhibition ofhost cell protein synthesis, p. 177-221. In H. Fraenkel-Conratand R. R.Wagner J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
(ed.), Comprehensive virology, vol. 19. Plenum Publishing Corp., New York.
9. Flanagan, J. B., R. F.Petterson,V. Ambros, M. J. Hewlett, and D. Baltimore. 1977. Covalent linkageof a protein to a defined nucleotidesequence atthe 5'-terminus of virion and replicative intermediate RNAs of poliovirus. Proc. Natl. Acad. Sci. USA 74:961-965.
10. Furuichi, Y., A. LaFiandra, and A.J. Shatkin. 1977. 5'-Ter-minal structure and mRNA stability. Nature (London) 266:235-239.
11. Hewlett,M.J., J. K. Rose, and D. Baltimore. 1976.5'-Terminal structureofpoliovirus polyribosomalRNAispUp. Proc.Natl. Acad. Sci. USA73:327-330.
12. Holland, J. J. 1961. Receptoraffinitiesasmajordeterminants of enterovirus tissue tropism inhumans. Virology15:312-326. 13. Jen, G., B. M. Detjen, and R. E. Thach. 1980. Shutoff of HeLa
cellprotein synthesis by ehcephalomyocarditis virus and polio-virus: a comparative study. J. Virol.35:150-156.
14. Katz, R. A., B. R. Cullen, R. Malavarca, and A. M. Skalka. 1986. Role of the avian retrovirusmRNAleader in expression: evidence for novel translational control. Mol. Cell. Biol. 6:372-379.
15. Kozak, M. 1983.Comparison of initiation of protein synthesis in procaryotes, eucaryotes, and organelles. Microbiol. Rev. 47: 1-45.
16. Laemmli,U.K.1970.Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
17. La Monica,N.,C. Meriam, and V. R.Racaniello.1986.Mapping ofsequences required formouseneurovirulence ofpoliovirus type 2Lansing.J. Virol.57:515-525.
18. Lee, K. A. W.; and N. Sonenberg. 1982. Inactivation of cap-bindingproteinsaccompanies theshut-off of host protein syn-thesis by poliovirus.Proc.Natl.Acad. Sci. USA79:3447-3451. 19. Lee, Y. F., A. Nomoto, B. M. Detjen, and E. Wiinier.1977. A protein covalently linked to poliovirus genome RNA. Proc. Natl. Acad. Sci. USA 74:59-63.
20. McClain, K., M. Stewart, M. Sullivan, and J. V. Maizel, Jr. 1981. Ribosomal binding sites on poliovirus RNA. Virology 113:150-167.
21. Nicklin,M.J. H., H. G. Krausslich, H. Toyoda,J.J. Dunn,and E. Wimmer. 1987. Poliovirus polypeptide precursors: expres-sionin vitro andprocessing byexogenous 3Cand 2A protein-ases. Proc. Natl.Acad. Sci. USA84:4002-4006.
22. Nicklin,M. J. H., H. Toyoda, M. G. Murray, and E. Wimmer.
1986. Proteolytic processing in the replication of polio and
relatedviruses. Bio/Technology 4:33-42.
23. Nobis, P., R. Zibire,G.Meyer, J. Kuhne, G. Warnecke, andG. Koch. 1985. Production ofa monoclonal antibody against an epitope of HeLa cells that is thefunctional poliovirusbinding site.J.Gen. Virol. 66:2563-2569.
24. Nomoto, A., Y. F. Lee,and E. Wimmer. 1976. The 5' end of poliovirusmRNAisnotcapped with m7G(5')ppp(5')Np. Proc. Natl. Acad.Sci. USA 73:375-380.
25. Okada, Y., G. Toda, H. Oka, A. Nomoto, and H. Yoshikura. 1987.Poliovirus infection of establishedhumanblood cell lines: relationship betweendifferentiation stageand susceptibilityor cellkilling. Virology 156:238-245.
26. Pelham, H. P. B., and R. J. Jackson. 1976.AnefficientmRNA dependenttranslationsystemfromreticulocyte lysates. Eur.J. Biochem. 67:247-256.
27. Pelletier, J.,G. Kaplan, V. R. Racaniello, andN. Sonenberg. 1988. Cap-independent translation of poliovirusmRNAis con-ferred by sequence elements within the 5' noncoding region. Mol.Cell. Biol. 8:1103-1112.
28. Pelletier,J., and N. Sonenberg. 1985. Insertionmutagenesisto increase secondary structurewithinthe 5'noncoding regionofa eucaryoticmRNAreduces translationalefficiency. Cell 40:515-526.
29. Perez-Bercoff, R. 1982. But is the 5' endofmessenger RNA always involvedin initiation? p. 242-252. In R. Perez-Bercoff (ed.), Proteinsynthesis in eukaryotes. PlenumPublishing Corp., NewYork.
30. Phillips,B.A., and A.Emmert.1986. Modulation ofexpression ofpoliovirusproteins in reticulocytelysates. Virology 148:255-267.
31. Ray, B. K., T. G. Lawson, J. C. Kramer, M. H. Cladaras, J. A. Grifo,R. D.Abramson,W.C. Merrick,and R. E. Thach.1985. ATP-dependent unwinding of messenger RNA structure by eukaryotic initiation factors. J. Biol. Chem. 260:7651-7658. 32. Rose, J. K., H. Trachsel, K. Leong, and D. Baltimore. 1978.
Inhibition of translationby poliovirus: inactivation ofaspecific initiation factor. Proc. Natl.Acad. Sci. USA 75:2732-2736. 33. Shih,D.S.,C. T.Shih,0. Kew, M. Pallansch, R.Rueckert,and
P. Kaesberg. 1978. Cell-free synthesis and processing ofthe proteins of poliovirus. Proc. Natl. Acad. Sci. USA 75:5807-5811.
34. Sonenberg, N. 1987. Regulation of translation by poliovirus. Adv. VirusRes.33:175-204.
35. Weber, L. A., E. R. Feman,E. D.Hickey,M.C.Williams,and C.Baglioni. 1976. Inhibition ofHeLacellmessage RNA
trans-lationby 7-methylguanosine 5'-monophosphate. J. Biol. Chem. 251:5657-5662.
![FIG.1.deletionobtained5'contain5'openpolyproteinofnucleotidepP2-5'.A5'-465/CATconstructssoft-laser[17])](https://thumb-us.123doks.com/thumbv2/123dok_us/1337972.87526/3.612.130.493.66.558/fig-deletionobtained-contain-openpolyproteinofnucleotidepp-a-catconstructssoft-laser.webp)