Copyright © 1987,American Society forMicrobiology
Structure
and
Expression
of the
Herpes Simplex
Virus
Type 2
Glycoprotein gB Gene
LAURAL. STUVE, SHERYL BROWN-SHIMER, CAROLPACHL, RICHARDNAJARIAN, DINODINA,
ANDRAE LYN BURKE*
Chiron Corporation, Emeryville, California 94608
26June 1986/Accepted 23 October 1986
Thegeneforglycoprotein gB2ofherpes simplexvirustype 2 strain 333wascloned, sequenced,andexpressed in mammalian cells. The gB2 proteinhadanoverallnucleotide and amino acidsequencehomologyof 86%with
the cognate gBl protein. However, of the 125 amino acid substitutions or deletions, only 12.5% were
conservativereplacements. Thesedifferenceswereclustered withinanNH2-terminal region, acentralregion,
and a COOH-terminal region, resulting in domains ofnear identity broken by small regions of marked divergence. Regionsof greatesthomologyincludeda90-amino-acidstretchstartingat residue 484 and 39amino acids spanning residues 835 to 873, which cover a rate-of-entry locus mapped to Ala-552 and a syn locus mappedtoArg-857, respectively, ingBlbyBziketal. (D. J. Bzik,B. A.Fox,N. A. DeLuca,andS.Person, Virology 133:301-314, 1984). Pellett etal. (P. E. Pellett, K. G. Kousoulas, L. Pereira, and B. Roizman, J. Virol.53:243-253, 1985) mappedthe mutations in three monoclonalantibody-resistant gBlmutantsbetween aminoacids 273 and 443. Theseepitopesareincluded inaregionof 98 residues identical betweengBlandgB2. Theidentityof thisproteinwasverifiedby placingatruncatedgenelackingthe 303carboxyl-terminalamino
acidsofgB2into mammalian COS and CHO cells. Expressionwasdemonstratedbyimmunofluorescenceand
radioimmunoprecipitation.Thisproteinwillbepurifiedfrom the stable CHO cell lines andcomparedwithgBl
forimmunogenicityand protectiveefficacyin animalchallenge models.
Herpes simplex virustype 2 (HSV-2) specifiesfiveto six major glycoproteins which areinserted into the viral enve-lopeaswell asintotheplasma membrane oftheinfectedcell (36). Theglycoprotein gB2 isoneofthemoreabundant viral glycoproteinsin theinfected cell. Byanalogywithstudiesof the cognate HSV-1 gBl gene, this glycoprotein plays a
centralrolein theadsorption of virus and inthepenetration of virus into the host cell (reviewed in reference 36). In addition, the viral glycoproteins and, in particular, gB are major elicitors of the host immune response (reviewed in reference 9). Thus, it is of interest to characterize and producethegB proteintoaidinunderstandingtheinitiation of viralinfectionaswellasthespecificityandprotectiverole ofthehostimmuneresponse.
The nucleotide sequence of the genefor HSV-1gBl has been determined (6, 26). The locations of several different gBl mutations which reduce the sensitivity of the virus to neutralization by monoclonal antibodies, alter the rate of virus penetration into the cell, and alter the morphology of viral plaques have been reported (7, 26). Based on serolog-ical analyses (11, 27), the gBl and gB2 glycoproteins are quite similar. However, there is some evidence of a type
specificity
in the viralattachment process (39, 40).Inthis report, we present the nucleotide sequence of the gB2 gene and report the establishment of permanent cell linessecretingatruncated gB2protein. In the accompanying paper (24) we describe the high-level expression, purifica-tion, and partial amino acid sequence analysis of the gBl protein.
MATERIALS AND METHODS
Cells, viruses, and reagents. The HSV-1 strain Patton and the HSV-2 strain 333 viruses were obtained from Richard
*
Corresponding
author.Hyman, Hershey Medical Center, Hershey, Pa., and prop-agated in Vero cells obtained from Evelyn Lennette, ViroLabs, Berkeley, Calif. The HindIII-H restriction
frag-mentof HSV-2 strain 333 (32) cloned into plasmid pBR322 (17) was kindly provided by Dr. Hyman. COS 7 cells (13) wereobtained from Robert Tjian, University of California, Berkeley, and grown in Dulbecco modified Eagle medium (Gibco Laboratories, Grand Island, N.Y.) supplemented with10% fetal calf serum (Hyclone, Logan, Utah), 100 U of penicillin per ml, and 100
pLg
ofstreptomycin per ml. Chinese hamster ovary (CHO) cells deficient in dihydrofolate reductase (DHFR-) were obtained and grownas described elsewhere (24).Biological reagents were purchased from Bethesda Re-search Laboratories,Gaithersburg, Md., and usedas speci-fied by the manufacturer. Standard DNA manipulations followedprocedures outlined byManiatis etal. (19).
Isolation ofRNA and Northern blot analysis. Total RNA was prepared from HSV-1-infected and HSV-2-infected Vero cells 6 h after infection with a multiplicity of 10 PFU/cell. Monolayers containing aproximately 2.4 x 108 infected cells were washed, incubated with 50 ml of extrac-tion buffer, and processed as described previously (25). Polyadenylated [poly(A)+] RNA was prepared byloading 2 mg of total RNA onto a 3-ml column of oligo(dT)-cellulose equilibrated in 500 mM NaCl-10 mMTris hydrochloride (pH 7.5)-l mM trisodium EDTA-0.1% sodium dodecyl sulfate (SDS). The column was washed with 10 ml of 100 mM NaCl-10 mMTris hydrochloride (pH 7.5)-i mM trisodium EDTA-0.1% SDS, and then the poly (A)' mRNA fraction was eluted with 10 ml of 10 mM Tris hydrochloride (pH
7.5)-i
mMtrisodium EDTA-0.1% SDS. For Northern blotanalysis, poly (A)+RNA was denatured with glyoxal (21), fractionated by electrophoresis on 1% agarose gels, trans-ferredto nitrocellulose paper (37), and hybridized with the 32P-labeled DNAprobes noted in Results.
326
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF HSV-2 gB GENE 327
Sequencing andinvitromutagenesis.DNAwassequenced
by thedideoxy chain termination method (34) after cloning restrictionfragments into the M13 phage derivative vectors
mpl8 and mpl9 (Pharmacia, Piscataway, N.J.). Extensive
use was made of synthetic deoxyoligonucleotide primers
complementary to specific HSV DNA sequences for se-quence extension on M13 templates and for verification of
ambiguous regions. These primers were synthesized by the
phosphoramidite method (20) with a DNA synthesizer (model 380A; Applied Biosystems, Foster City, Calif.). In addition, alternate precursors, eitherdeoxyinosine triphos-phate (dITP) (Pharmacia) (22) or deoxy-7-deazaguanosine
triphosphate (c7dGTP) (obtained from F. Seela, University
of Paderborn; now available from Boehringer Mannheim,
Indianapolis, Ind.) was substituted into the dideoxy-deoxynucleoside triphosphate mixinplace of dGTP, bothat
fivefold-higher concentrations than that of dGTP. In addi-tion, gels containing 7 M urea, 25% formamide, and 7%
acrylamide were usedto help eliminate band compressions.
The PstI site(5'-CTGCAG-3')atposition -61 (relativeto the initial ATG codon) was converted to a unique HindIII site (5'-AAGCTT-3') by in vitro mutagenesis to facilitate precise insertion into the expression vector. The 34-base oligonucleotide 5'-TCCTCTACTCGGTCAAGCTTCA-CCTCGCCCCCAG-3', complementary (exceptasindicated
by the four underlinedbases) tonucleotides -75 to -42of
thenoncoding strand of the HSV-2 gBsequence(see Fig. 3),
wasusedas aprimer for synthesisontheM13template (44).
The 3.46-kilobase (kb) NruI-BamHI fragment of pHS208 (see Fig. 1) was subcloned into SmaI-BamHI-digested M13mpllreplicative form (RF) and used totransform com-petent Escherichia coliJM101. Recombinantphage contain-ing the HSV-2 insert were identified by hybridization to nitrocellulose filterreplicasof the plaques at67°C with the
32P-labeled 34-mer as the probe (3). After the recombinant
phage DNAs wereused as templates for mutagenesis, mu-tant phage were detected by hybridization with the same
oligomeras aprobein6x SSC(1x SSC is 0.15 M NaClplus
0.015 Msodiumcitrate)-10x Denhardt solution(19)at73°C.
The filterswere then washed in 6x SSC at73°C.
DNAtransfections. All DNA transfectionswereperformed as described elsewhere (24), with the following modifica-tions. The selective medium used for CHO DHFR- cells contained 10% fetal calfseruminplaceofdialyzedfetalcalf serum. For COS 7 celltransfections,thecellswereplatedin two-chamber glass slide wells (Lab-Tek; Miles Scientific, Naperville, Ill.) and grown to 75% confluency. The glass
slides had been precoated for 1 h at 20°C with 0.5 ml of filter-sterilized1%gelatininphosphate-bufferedsaline(PBS;
0.15 M NaCl, 2.7 mM KCl, 15.3 mM Na2HPO4, 1.5 mM
KH2PO4) per well, followed by three rinses with PBS. A
250-,ul portion of the calcium phosphate-DNA suspension containing4 p.gofplasmidwas addedto2 ml of mediumper
slide well. After a 7.5-h inclubation at 37°C, the DNA solution was removed and the cell monolayer was rinsed once with fresh medium. The brief incubation in 15%
glyc-erol was omitted. At 33 h after transfectionthe cells were fixed with 100% methanol for analysis ofgB2 synthesis by
immunofluorescence as described in the accompanying manuscript (24), except that the primary antibody was an anti-HSV-2 rabbit antiserum (DAKO, Santa Barbara, Calif.).
ELISA analysis. Portions (0.1 ml) of conditioned medium harvested from confluent 24-well cultures of CHO DHFR+
cells which had been incubatedfor 24 h in atotal volume of 0.25 mlof mediumwere assayed by capture enzyme-linked
immunosorbent assay (ELISA) as outlined for gBl (24), except that the anti-HSV-2 rabbitantiserum wasused as the secondary antibody at a 1:100 dilution.
In vivo labeling of cells and immunoprecipitation. Cells were labeled with [35S]methionine and immunoprecipitated asdescribed in the accompanying manuscript (24) with the following modifications. Duplicate 3.5-mm dishes of confluent cellswere labeled with [35S]methionine for 1 h at 37°C. At the end of the labeling period, the medium was removed and the cell monolayers were washed with PBS. The cells from one dish were lysed in lysis buffer lacking 0.1% SDS. The remaining dish was incubated for an addi-tional 5 h at 37°C with Dulbecco modified Eagle medium containing 2.5 mM methionine, and then the cells were processed as above. The anti-HSV-2 rabbit antiserum (5
RI1)
wasused to precipitate gB2 from 50Rl
of cell lysate, and 15,ul
was used for 200 ,ul of cellmedium.Immunoprecipitates
were analyzed on 10% SDS-polyacrylamide gels (18), 1.5mmthick.
RESULTS
Identification and subcloning of a DNA fragment encoding gBof HSV-2. The gene encoding glycoprotein B of HSV-2 has been shown to be colinear with the corresponding HSV-1 gB gene by analysis of HSV-1 x HSV-2 intertypic recombinants and to lieapproximately between map coordi-nates0.30and 0.42 in the prototype arrangement (33). Thus, theHindlIl H fragment of HSV-2, which spans map coordi-nates 0.28 to 0.40 (10, 32) (Fig. 1), should include the gB2 coding region. In addition to their colinear map location, serological (11, 27) and heteroduplex (17) analyses indicated the closesimilarityof gBland gB2. Therefore, to locate the gB2coding region, fragmentsof the gBl gene (Fig. 1B, line I)wereusedtoprobeSouthern blots of restriction digests of theHSV-2 HindlIl Hfragment. The 0.55-kbPstI-SalI
frag-mentthat encodesamino acids323 to 506 of gBl hybridized
to a 2.6-kbXhoI fragmentand a 2.0-kb SstIfragment. The adjacent 0.49-kb NcoI fragment of gBl hybridized to the flanking3.2-kb XhoI bandaswellas a4.2-kbSphI band(data
notshown). Twooftheseoverlapping restriction fragments were subcloned into apBR322plasmid derivative to
gener-ate theplasmids pHS203 (containingthe 5' endof the gene
on a2.6-kbXhoI fragment) and pHS207 (containing the 3' end of the geneon a4.2-kbSphI fragment)(Fig. 1B, line III). Theexactlocation and theidentity ofthe gB2 gene were verified by probing a Northern blot of poly (A)' mRNA isolated from HSV-2-infectedVerocells with the restriction fragments of pHS203 shown in Fig. 1B, line II. Both a 0.89-kb XhoI-SmaI and a 1.29-kb SmaI-NruI fragment hy-bridized to anabundant 3.0-kb mRNA, anappropriate size andrepresentational frequencyforgB2by analogywith the analysis of gBl transcripts (6, 14, 30). However, the same
mRNAdidnothybridizetotherightmost0.47-kb NruI-XhoI fragment (data not shown). As expected, a similar hybrid-ization patternwasobserved whenpoly
(A)'
RNAprepared from HSV-1-infected Vero cells was probed with these fragments (data not shown). Since this analysis indicated boththelimit ofthe right-handendofthegB2 geneand its size, it was apparent that the gB2coding
sequences mustextend an additional 1 kb to the left of those sequences contained within
pHS203
intotheoverlapping pHS207
plas-mid. Therefore,thegB2genewasclonedasonecontinuous fragment by ligation of the 1.98-kbNruI-SphI
fragrnent
of pHS203tothe1.48-kbSphI-BamHI
fragment
ofpHS207
and insertion intoNruI-BamHI-digested pBR322
to generate pHS208.VOL.61,1987
on November 10, 2019 by guest
http://jvi.asm.org/
0.2
Map Position
0.4 0.6
B IN, H
0.28 ,"0.30 0.32
, l
B. r7IL
I
IL
H B XE Bs K BBgE KSp Hp X
11
III F
E
R
A_
0_~~~~~~~~~~~~~~~~~~~~~~~~~
,L0.34 0.36-,_ 0.38
3 5 I
,s
Bs S KI
Nc Nc PS
pHS207
il X Sm
Sm N NX pHS203
II
pHS208
FIG. 1. (A)Physicalmapof HSV-2 DNA. The conventionalmapfor theprototypeconfigurationis shown inthe firsttwolines(31).The
hatched boxes correspond tothe terminalrepeatelements. Therestrictionmapfor HindlIl is shown in the third line (10, 32). (B)Partial
restrictionmapof HindIII-H(17.2 kb). Black bar correspondstogB2 coding region.Restriction sites:B,BamHI; Bg, BglII; Bs, BstE2; E, EcoRI; H, HindIll; Hp, HpaI; K, KpnI; Sp, SphI; X, XhoI. LineI: Location ofHSV-2-homologous DNA sequencefor two HSV-1 DNA
fragments usedashybridization probestoSouthernblots of restrictiondigestsof the HSV-2 HindIII-H DNAsegment.Restriction sites:Nc, NcoI; P, PstI; S, Sall. For PstI andSall, thehomologous sitesareconserved in the HSV-2 DNA. LineII: HSV-2 restrictionfragments obtained from plasmid pHS203 and used as hybridization probes to Northern blots of poly (A)' RNA prepared from HSV-1- and
HSV-2-infected Vero cells. Restriction sites: N, NruI; Sm, SmaI; X, XhoI. LineIII:Maplocations of the HSV-2 DNA inserts withinplasmids pHS203,pHS207, and pHS208.
A.
0
BasePairs
500 1000 1500 2000 2500 3000 3500
NScP P Sc SSm S P Sp PvX Pv Pv Sc B
I;
1
1
1
11 11
1i
1
-<-1
-.|<
(
I I I I II I *
1H~ ~ <_ F-i- I_
-~~
X
~
~
||-
0f
B.
l
f
,,,
,
,
,,
,,
3< It 'i' ti??Nil? tflffY tli ll l It f,,
5 8ii
IT,
I II I II II II I I II I III II ll Iii ?l IFIG. 2. (A)DNA sequencing strategy and partial restriction map for the 3.46-kbNruI-BamHI(map position0.344to0.367)DNAfragment ofHSV-2encodingglycoprotein gB. The orientation of this map is reversed from that shown in Fig. 1, so that transcription proceeds from lefttoright,5' to3', throughthecoding region. Restriction map (abbreviations: B,BamHI;N, NruI;P,PstI;Pv, PvuII; S,SaIl;Sc, Sacl;
Sp, SphI;X,XhoI)is shown on the second line. The specific restriction fragments used are indicated by the narrow horizontal lines with vertical ends;arrowsindicate the extent anddirection of each sequence analysis. Template-specific synthetic oligonucleotides were used for priming internalstretches that did not initiate from a restriction site. (B) Open reading frame analysis. The locations of potential translational initiationcodonsare indicated by the balloons, and stop codons are indicated by the vertical lines. The top three frames (1 to 3) read left to right, 5' to3',whilethe bottom three frames (4 to 6) read right to left, 5' to 3'. Reading frame 2 encodes gB2.
0 0.8 1.0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.139.489.70.282.2] [image:3.612.112.535.398.651.2]STRUCTURE OF HSV-2 gB GENE 329
Analysis of the gB2 DNA sequence. An outline of the sequencing strategy for the 3.46-kbNruI-BamHI (map coor-dinates 0.344 to 0.367) HSV-2 fragmentencoding glycopro-tein B is shown in Fig. 2A. This figure is inverted with respect to the HSV prototype orientation shown inFig. 1,so
that the gB transcript reads from left toright, 5' to3'. Both strands were sequenced in their entirety. In addition, the 2.6-kb XhoI insert in pHS203 was sequenced from the leftmost XhoI site through theSphI site which had been used to ligate the twogBl gene fragments. This sequence analysis assured us that there were no small, internal fragments deleted from thefinal NruI-BamHI gene clone.
To facilitate the sequencing of very largerestriction frag-ments cloned into the M13 derivative phages mpl8 and mpl9,we used synthetic oligonucleotidescomplementary to portions of the HSV sequence that we had determined primed a second round of synthesis on the same phage template. Thus, the DNA region accessible to sequence analysis was extended. To helpresolve band compressions in thesequencing gel, which are commonfor DNAs of high G+C base composition (22), we used two separate dGTP analogs, dITP and c7dGTP, as substitutes for dGTP in the polymerization reaction (2, 22).The dITP analogdisruptsthe normal G-C base pairing, making only two, rather than three, hydrogenbonds withcytosine, whereas Watson-Crick base pairing is unaltered for c7dGTP (35). However, Hoogsteen bond formation is completely precluded for c7dGTP by the removal of the nitrogen at the 7 position, whereas for dITP, two hydrogen bonds are formed in the Hoogsteenconformation (35). Thus,each analog has unique utility in destabilizing thesecondarystructure that generates the sequencingartifact ofband compressions.
The six possible translational reading frames for the gB sequence are shown in Fig. 2B, with the locations of methionine and stop condons indicated. In frame 2, a large open readingframe of2,748 nucleotides encodes a polypep-tide of the appropriate molecular weight for gB2. This protein is homologous to the deduced amino acid sequence for HSV-1 gB strain Patton as shown here and as reported for strain KOS (6) and strain F (26). The predicted protein sequence includes the syncytial and rate-of-entry loci (7) as well as the monoclonal antibody resistanceloci (26) located in gBl. Frame 6 actually contains the largestopen reading frame of 2,889 nucleotides, but this frame is unlikely to encode gB2because thepredicted protein sequencebears no homology to gBl and this strand of the DNA did not hybridize to the gBl mRNA (14; C. Pachl, unpublished results). No other open reading frame is large enough to encode gB2 (in theabsence ofa splicedmRNA, whichisnot expected for this viralprotein).
Thenucleotide sequenceforgB2 is shown in Fig. 3, line 2. The predicted amino acid sequence ofgB2 is shown above theDNA sequence.Forcomparison,the DNA sequenceand the amino acid sequence ofgB1 from HSV-1 strain Patton are shown below. Spaces have been inserted into the se-quences to permit maximal alignment of the two proteins. All numbers in Fig. 3 refer to the gB2 sequence. The sequence ATATAT which abuts 5' to the gB2 NruI site shown in Fig. 3 is most likely the TATA transcriptional regulatory sequence, since it is completely homologous to the corresponding region in gBl (6, 26). Our sequence analysis did not extend further upstream to the putative location of the CAT box and othertranscriptionalregulatory elements. In the 3' noncoding region, a polyadenylation signal, AATAAAAA (29), at residues 2744 to 2751 is the probable termination site of the gB2 mRNA. (Allnucleotides
and amino acids are numbered withreferencetothe trans-lational startcodonATG at +1 to +3.)
Primary structure of the gB2 protein. The predicted gB2 protein is 904 amino acids long and contains elements characteristic ofamembraneglycoprotein. After cleavageof the predicted 22-amino-acid signal sequence, the mature, nonglycosylated protein would have amolecular weight of 98,221. The amino-terminal 22residues contained acore of hydrophobic residues (Leu-6 to Ala-20) preceded by a charged basic residue (Arg-2) and an alanine-rich signal peptidase recognition sequence, Ala-20-Ser-21-Ala-22, con-forming to rules identified for preferred signal peptidase cleavage sites and the general characteristics ofeucaryotic signal peptides(28, 42, 43). Protein sequence analysis ofthe N terminus of recombinant HSV-1 glycoprotein B (24) identifiedthefirst amino acid ofthe mature type 1 protein as Ala-31, followed by Pro-Ser-Ser-Pro. Due to the conserva-tion of the six amino acids centered around the signal cleavage recognition sequence, Ala-23 is probably the first amino acid of the mature gB2 glycoprotein.
The external hydrophilic region of the protein from Ala-23 to Asp-723 contained eight possible sites for N-linked glycosylation identified by the sequence Asn-X-Thr/Ser, where Xis any of the 20 amino acidsexcept perhaps aspartic acid (16). There was a potential transmembrane anchor region of 54 amino acids from Ala-745 to Leu-798. Analysis by the Chou and Fasman algorithm (8) indicated a mixed
1-sheet
anda-helixpotential
for this entireregion.
However,
toavoid orientationof polarsidechains toward thelipophilic environment ofthe membrane bilayer, it is likely that this region adopts an a-helical conformation (12). An ahelix of this length (8.1 nm) would be more than sufficient to spana biological membrane 3 nm thick two times, placing the C-terminal domain ofthe proteinonthe exterior of the cell. Alternatively, asproposed for gBl (26), the transmembrane domain may traverse the membrane threetimes and include the amphipathic domain beginning at Asp-723 that contains four additional charged residues,localizingtheCterminus of the protein within the cytoplasm. The C-terminal region of gB2 extended from the end of the membrane anchor region atLeu-798 tothe end of theprotein at Leu-904 and contained a high density of charged residues. No potential N-linked glycosylation sites werepresentin thisportionofthe HSV-2 protein.
Comparison of gB2 with gBl.Acomparison of theprimary sequencesof the HSV-1 andHSV-2gB isalso shown inFig. 3. Amino acid differences between the HSV-2 and HSV-1 proteins are enclosed by boxes. Overall, the two proteins have anucleotide and amino acid homology of 86%. How-ever, the differences appear to be significant, since only 12.5% of the amino acid substitutions between gBl and gB2 were conservative changes. These differences in primary sequence were clustered, resulting in long peptide domains which were identical, as well as small regions of marked divergence.
One region ofdissimilarity was the signal sequence. For gB2, the predicted signal sequence wasonly 22 amino acids long, compared with 30 for gBl strain Patton (the signal sequence for gBl strain Patton is 30 amino acids long, comparedwith 29 amino acids forgBl strain F)and hadonly 55% amino acid homology with the gBl protein. It is of interest that while the length of the entire coding sequence for gBl andgB2 was the same(904amino acids),the mature gB2 would be 7 amino acids longer than gBl due to its shorter signal peptide.
The homologyofgBland gB2wasrevealedbya compar-VOL. 61,1987
on November 10, 2019 by guest
http://jvi.asm.org/
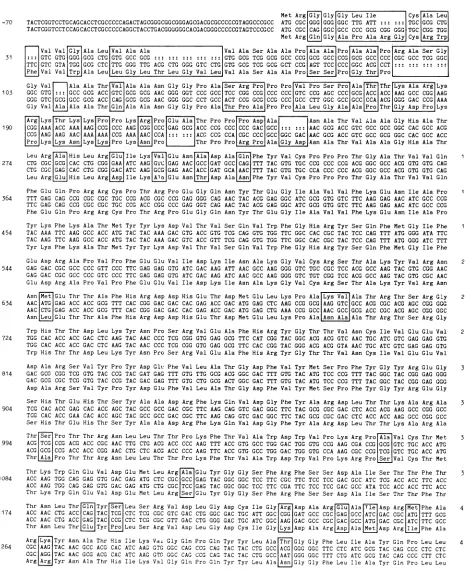
-308 TCGCGAGCTCATTATCGCCACC ACACTCTTTGCGTCGGTCTACCGGTGCGGGGAGCTTGAGTTGCGCCGCCCCGACTGCAGCCGCCCGACCTCCGAAGGTCTGTACCGCTACCCGCCGG
TCGCGAGCTGATTATCGCCACCACACTCTTTGCCTCGGTCTACCGGTGCGGGGAGCTCGAGTTGCGCCGCCCGGACTGCAGCCGCCCGACCTCCGAAGGTCGTTACCGTTACCCGCCCG
-189 GCGTGTACCTCACGTACAACTCCGACTGTCCGCTGGTGGCCATCGTCGAGAGCGGCCCCGACGGCTGCATCGGACCCCGCTCGGTCGTGGTTTACGACCGAGACGTTTTTTCCATCCTC
GCGTATATCTCACGTACGACTCCGACTGTCCGCTGGTGGCCATCGTCGAG AGCGCCCCCG ACGG CTGTATCGGCCCC CGGTCGGTCGTGGTCTACG ACCGAGACGTTTTCTCGATCCOTC
Met Arg GlyGly Gly Leu Ile Cys Ala Leu -70 TACTCGGTCCTGCAGCACCTCGCCCCCAGACTAGCGGGCGGCGGGAGCGACGCGCCCCCGTAGGCCCGCC ATG CGC GGG GGG GGC TTG ATT TGC GCG CTG
TACTCGGTCCTCCAGCACCTCGCCCCCAGGCTACCTGACGGGGGGCACGACGGGCCCCCGTAGTCCCGCC ATG CGC CAG GGC GCC CCC GCG CGG GGG TGC CGG TGG Met Arg Gln Gly Ala Pro Ala Arg Gly Cys Arg Trp
[ Val ValGly Ala Leu Val Ala Ala Val Ala Ser Ala Ala Pro Ala Ala Pro Ala Ala Pro Arg Ala Ser Gly
31 :::GTC GTG GGG GCG CTG GTG GCC GCG GTG GCG TCG GCG GCC CCG GCG GCC CCG GCG GCC CCC CGC GCC TCG GGC TTC GTC GTA TGG GCG CTC TTG GGG TTG ACG CTG GGG GTC CTG GTG GCG TCG GCG GCT CCG AGT TCC CCC GGC ACG CCT
Phe Val Val[rp Ala Leu Leu Gly Leu Thr Leu Gly Val Leu Val Ala Ser Ala Ala Pro Pro Gly Thr Pro
Gly Val Ala Ala Thr Val Ala Ala Asn Gly Gly Pro Ala Ser Arg Pro Pro Pro Val Pro Ser Pro AlaThr Thr Lys Ala Arg Lys
105 GGC GTG ::: GCC GCG ACC GTC GCG GCG AAC GGG GGT CCC GCC TCC CGG CCG CCC CCC GTC CCG AGC CCC GCG ACC ACC AAG GCC CGG AAG GGG GTC GCG GCC GCG ACC CAG GCG GCG AAC GGG GGC CCT GCC ACT CCG GCG CCG CCC GCC CTT GGC GCC GCC CCA ACG GGG GAC CCG AAA
Gly Val Ala Ala Ala ThrGln Ala Ala Asn Gly Gly Pro Ala Thr Pro Ala Pro Pro Ala Leu Gly Ala Ala Pro Thr Gly AspPro Lys Arg Lys|Thr Lys Lys Pro Pro LysaAg Pro Glu Ala Thr ProPro Pro Asp Ala Asn Ala Thr Val Ala Ala Gly HisAla Thr 190 CGG AAA ACC AAA AAG CCG CCC AAG CGG CCC GAG GCG ACC CCG CCC CCC GAC GCC AAC GCG ACC GTC GCC GCC GGC CAC GCC ACG CCG AAG AAG AAC AAA AAA CCG AAA AACICCA A::::::IACG CCG CCA CGC CCC GCC GGC GAC AAC GCG ACC GTC GCC GCG GGC CAC GCC ACC jProLysjLys AsnJLys[LysPro Lys Prol |Thr Pro Pro|Arg Pro Ala Gly AspAsn Ala Thr Val Ala Ala Gly HisAla Thr Leu Arg Ala His Leu Arg G IleLys[Va Glu AsnAla Asp Ala Gln Phe Tyr Val Cys Pro Pro Pro Thr Gly Ala Thr Val Val Gln 1
274 CTG CGCIGCGICAC CTG CGG GAA ATC AAG GTC GAG AACIGCCGGAT GCC CAG TTT TAC GTG TGC CCG CCC CCG ACGGGC GCC ACG GTG GTG CAG
CTG CGC GAG CAC CTG CGG GAC ATC AAG GCG GAG AAC ACC GAT GCA|AAC TTT TAC GTG TGC CCA CCC CCC ACG GGC GCC ACG GTG GTG CAG Leu ArgLGJuHis Leu Arg[spJIle Lys[Ala Glu Asn ThrAsp Ala|Asn|Phe Tyr Val CysPro Pro Pro Thr Gly Ala Thr Val ValGln
Phe Glu Gln Pro Arg Arg Cys Pro Thr Arg Pro Glu Gly Gln Asn Tyr Thr Glu Gly Ile Ala Val Val Phe Lys Glu Asn Ile Ala Pro 1
364 TTT GAG CAG GCG CGC CGC TGC CCG ACG CGC CCG GAG GGG CAG AACTAC ACG GAG GGC ATC GCGGTG GTC TTC AAG GAG AAC ATC GCC CCG TTC GAG CAG CCG CGC CGC TGC CCG ACC CGG CCC GAG GGT CAG AAC TAC ACG GAG GGC ATC GCG GTG GTC TTCAAG GAG AAC ATC GCC CCG Phe Glu Gln Pro Arg Arg Cys Pro Thr Arg Pro Glu Gly Gln AsnTyr Thr GluGly Ile AlaVal Val Phe Lys Glu Asn Ile Ala Pro Tyr Lys PheLys Ala Thr MetTyr Tyr Lys AspVal ThrVal SerGln Val Trp Phe Gly His Arg TyrSer Gln Phe Met Gly Ile Phe 1 454 TAC AAA TTC AAG GCC ACC ATG TACTAC AAA GAC GTG ACC GTG TCG CAG GTG TGG TTC GGC CAC CGC TAC TCC CAG TTT ATG GGG ATA TTC
TAC AAG TTC AAG GCC ACC ATG TAC TAC AAA GAC GTC ACC GTT TCG CAG GTG TGG TTC GGC CAC CGC TAC TCC CAG TTT ATG GGG ATC TTT Tyr Lys Phe Lys Ala Thr Met Tyr Tyr Lys Asp Val Thr Val Ser Gln Val Trp Phe GlyHis Arg TyrSer GlnPhe Met Gly Ile Phe Glu Asp Arg Ala Pro Val Pro Phe Glu Glu Val Ile Asp Lys Ile Asn Ala Lys Gly Val Cys Arg Ser Thr Ala Lys Tyr Val Arg Asn 2 544 GAG GAC CGC GCC CCC GTT CCC TTCGAG GAG GTG ATC GAC AAG ATT AAC GCC AAG GGG GTC TGC CGC TCC ACG GCCAAG TACGTG CGG AAC
GAG GAC CGC GCC CCC GTC CCC TTC GAG GAG GTG ATC GAC AAG ATC AAC GCC AAG GGG GTC TGT CGG TCC ACG GCC AAG TAC GTG CGC AAC Glu Asp Arg Ala Pro Val Pro PheGlu Glu Val Ile Asp Lys lie Asn Ala Lys Gly Val Cys ArgSer Thr AlaLys Tyr Val ArgAsn
Asn Met Glu Thr Thr Ala Phe His Arg Asp Asp His Glu Thr Asp MetGlu Leu Lys Pro Ala|Lys ValAla Thr Arg Thr SerArg Gly 2
634 AAC|ATG GAG ACC ACC GCG TTT CAC CGG GAC GAC CAC GAG ACC GAC ATG GAG CTC AAG CCG GCGIAAG GTC|GCCACG CGC ACG AGC CGG GGG AACICTGIGAG ACC ACC GCG TTT CAC CGGGAC GAC CAC GAG ACC GAC ATG GAG CTG AAA CCG GCCIAACGCC GCG ACC CGC ACG AGC CGG GGC Asn Leu Glu Thr Thr Ala Phe His Arg Asp Asp His Glu Thr AspMet Glu Leu Lys Pro Ala|Asn Ala Ala ThrArg Thr Ser Arg Gly
Trp His Thr Thr Asp LeuLys Tyr Asn Pro Ser Arg Val Glu Ala Phe His Arg Tyr Gly Thr Thr Val Asn Cys Ile Val Glu GluVal 2 724 TGG CAC ACC ACC GAC CTC AAG TAC AAC CCC TCG CGG GTG GAG GCG TTC CAT CGG TAC GGC ACG ACG GTC AAC TGC ATC GTC GAG GAG GTG
TGG CAC ACC ACC GAC CTC AAG TAC AAC CCC TCG CGG GTG GAG GCG TTC CAC CGG TAC GGG ACG ACG GTA AAC TGC ATC GTC GAG GAG GTG Trp His Thr Thr Asp Leu Lys TyrAsn Pro Ser ArgVal Glu Ala Phe His Arg TyrGly Thr Thr Val Asn Cys Ile Val Glu Glu Val Asp Ala Arg Ser Val Tyr Pro Tyr Asp Glt Phe Val Leu Ala Thr Gly Asp Phe Val Tyr MetSer Pro Phe Tyr Gly Tyr Arg Glu Gly 3 814 GAC GCG CGG TCG GTG TAC CCG TAC GAT GAG TTT GTG TTG GCG ACG GGC GAC TTT GTG TAC ATG TCC CCG TTT TACGGC TAC CGG GAG GGG
GAC GCG CGC TCG GTG TAC CCG TAC GAC GAG TTT GTG CTG GCG ACT GGC GAC TTT GTG TAC ATG TCC CCG TTT TAC GGC TAC CGG GAG GGG Asp Ala Arg Ser Val Tyr Pro Tyr AspGlu Phe Val Leu Ala Thr Gly Asp Phe Val TyrMet Ser Pro Phe TyrGly Tyr Arg Glu Gly
Ser His ThrGlu His Thr Ser Tyr Ala Ala Asp Arg Phe Lys Gln Val Asp Gly Phe Tyr Ala Arg Asp Leu Thr Thr Lys Ala Arg Ala 3
904 TCG CAC ACC GAG CAC ACC AGC TAC GCC GCC GAC CGC TTC AAG CAG GTC GAC GGC TTC TAC GCG CGC GAC CTC ACC ACG AAG GCC CGG GCC TCG CAC ACC GAA CAC ACC AGC TAC GCC GCC GAC CGC TTC AAG CAG GTC GAC GGC TTC TAC GCG CGC GAC CTC ACC ACC AAG GCC CGG GCC Ser His Thr Glu His Thr Ser Tyr Ala Ala Asp Arg Phe Lys Gln Val Asp Gly Phe Tyr Ala Arg Asp Leu Thr Thr Lys Ala Arg Ala Thr[er Pro Thr Thr Arg Asn Leu Leu Thr Thr Pro Lys Phe Thr Val Ala Trp Asp Trp Val Pro Lys Arg Pro Ala Val Cys Thr Met 3
994 ACGITCG CCG ACG ACC CGC AAC TTG CTG ACG ACC CCC AAG TTT ACC GTG GCC TGG GAC TGG GTG CCG AAG CGA CCGIGCG GTC TGC ACC ATG ACG|GCG CCG ACC ACC CGG AAC CTG CTC ACG ACC CCC AAG TTC ACC GTG GCC TGG GAC TGG GTG CCA AAG CGC CCG|TCGIGTC TGC ACC ATG Thr[AlaPro Thr Thr Arg Asn Leu Leu Thr Thr Pro Lys Phe Thr Val Ala Trp Asp Trp Val Pro Lys ArgProe Val Cys Thr Met Thr Lys Trp Gln Glu Val Asp Glu Met Leu Arg[Aa Glu Tyr Gly Gly Ser Phe Arg Phe Ser Ser Asp Ala Ile Ser Thr Thr Phe Thr 3
1084 ACC AAG TGG CAG GAG GTGGAC GAG ATG CTC CGCIGCC GAG TAC GGC GGC TCC TTC CGC TTC TCC TCC GAC GCC ATC TCG ACC ACC TTC ACC
ACC AAG TGG CAG GAG GTG GAC GAG ATG CTG CGCITCC GAG TAC GGC GGC TCC TTC CGA TTC TCC TCC GAC GCC ATA TCC ACC ACC TTC ACC Thr Lys Trp Gln Glu Val Asp Glu Met Leu Arg e Glu Tyr Gly Gly Ser Phe Arg Phe Ser Ser Asp Ala Ile Ser Thr Thr Phe Thr Thr Asn Leu Thr Gin Tyr Ser Leu Ser Arg Val Asp Leu Gly Asp Cys Ile Gly Arg Asp Ala Arg Ala Tie Asp Arg Met Phe Ala 4 1174 ACC AAC CTG ACC CAG TAC TCG CTC TCG CGC GTC GAC CTG GGCGAC TGC ATT GGCICGGGGAT GCC CGCGAG GGCCATC GAC CGCIATG TTT GCG
ACC AAC CTG ACC GAG TAC CCG CTC TCG CGC GTT GAC CTG GGG GAC TGC ATCGGC AAG|GAC GCC CGC GAC GCC ATGGAC CGC ATC TTC GCC Thr Asn Leu ThrjGluTyr[PoLeuSer ArgVal Asp Leu Gly Asp Cys Ile Gly(LsAspAla
ArgjApAlaMetAsp
Arg[lePhe Ala Arg[ys Tyr Asn Ala Thr His Ile Lys Vao Gly Gln Pro Gln Tyr Tyr Leu Ala Thr Gly Gly Phe Leu Ile Ala Tyr Gln Pro Leu Leu 41264 CGC|AAG|TAC AAC GCC ACG CAC ATC AAG GTGGGC CAG CCG CAG TAC TAC CTG GCCIACGGGGG GGC TTC CTC ATC GCG TAC CAG CCC CTC CTC CGC AGG| TAC AAC GCG ACG CAC ATC AAG GTC GGC CAG CCG CAG TAC TAC CTG GCCIAATIGGG GGC TTT CTG ATC GCG TAC CAG CCC CTT CTC ArgjrJTyr Asn Ala Thr His Ile Lys Val Gly Gln Pro Gln Tyr Tyr Leu Ala[snGly Gly Phe Leu Ile Ala Tyr Gln Pro Leu Leu
FIG. 3. DNA sequence of HSV-2
glycoprotein
gB gene.The top twolines snowtheDNA and thepredictedamino acidsequence for HSV-2 strain 333 gB2.Thebottomtwolines show thecorresponding
gBl
gene andderivedamino acidsequence forHSV-1 strainPatton.Throughout
thecoding region,
spaces have been insertedasrequired
tomaximize homologybetweenthe two proteins. The amino aciddifferences betweenthetype1 andtype2
proteins
arehighlighted by
boxes.Allnucleotides andaminoacids arenumbered withreferencetothetranslationalstartcodon ATG
assigned
as1,andonly
thetype2sequenceis numberedinthisfigure.
Differences betweenthegBl
strain Patton sequenceand thosestrains KOSand Fareconsideredin Table 1.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.77.547.116.689.2]61, 1987 OF gB
Ser Asn Thr Leu Ala Glu Leu Tyr Val Arg Glu Tyr Met ArgGlu Gln Asp Arg Lys Pro Arg Asn Ala Thr Pro Ala Pro Leu Arg Glu 481 1354 AGC AAC ACG CTC GCC GAG CTG TAC GTG CGG GAG TAC ATG CGG GAG CAG GAC CGC AAG CCC CGG AAT GCCACG CCC GCG CCA CTG CGG GAG
AGC AAC ACG CTC GCG GAG CTG TAC GTG CGG GAA CAC CTC CGA GAG CAG AGC CGC AAG CCC CCA AAC CCC ACG CCC CCG CCG CCC Ser Asn Thr Leu Ala Glu Leu Tyr Val Arg Glu His Leu Arg Glu Gln Ser Arg Lys Pro Pro Asn Pro Thr Pro Pro Pro Pro
Ala Pro Ser Ala Asn Ala Ser Val Glu Arg Ile Lys Thr Thr Ser Ser Ile Glu Phe Ala Arg Leu Gln Phe Thr Tyr Asn His Ile Gln 511 1444 GCG CCC AGC GCC AAC GCG TCC GTG GAG CGC ATC AAG ACC ACC TCG TCG ATC GAG TTC GCC CGG CTG CAG TTT ACG TAT AAC CAC ATA CAG
GGG GCC AGC GCC AAC GCG TCC GTG GAG CGC ATC AAG ACC ACC TCC TCC ATC GAG TTC GCC CGG CTG CAG TTT ACG TAC AAC CAC ATA CAG
Gly Ala Ser Ala Asn Ala Ser Val Glu ArgIle Lys Thr Thr Ser Ser Ile Glu Phe Ala ArgLeu Gln Phe Thr Tyr Asn His Ile Gln Arg His Val Asn Asp Met Leu Gly Arg Ile Ala Val Ala Trp Cys Glu Leu Gln Asn His Glu Leu Thr Leu Trp Asn Glu Ala Arg Lys 541
1534 CGC CAC GTG AAC GAC ATG CTG GGG CGC ATC GCC GTC GCG TGG TGC GAG CTG CAG AAC CAC GAG CTG ACT CTC TGG AACGAG GCC CGC AAG CGC CAT GTC AAC GAT ATG TTG GGC CGC GTT GCC ATC GCG TGG TGC GAG CTG CAG AAT CAC GAG CTG ACC CTG TGG AAC GAG GCC CGC AAG
ArgHis Val Asn Asp Met Leu Gly ArgVal Ala Ile Ala Trp Cys Glu Leu Gln Asn His Glu Leu Thr LeuTrp Asn Glu Ala ArgLys
Leu Asn Pro Asn Ala Ile Ala Ser Ala Thr Val Gly Arg Arg Val Ser Ala Arg Met Leu Gly Asp Val Met Ala Val Ser Thr Cys Val 571
1624 CTC AAC CCC AAC GCC ATC GCC TCC GCC ACC GTC GGC CGG CGG GTG AGC GCG CGC ATG CTC GGA GAC GTC ATG GCC GTC TCC ACG TGC GTG CTG AAC CCC AAC GCC ATC GCC TCG GCC ACC GTG GGC CGG CGG GTG AGC GCG CGG ATG CTC GGC GAC GTG ATG GCC GTC TCC ACG TGC G3TG Leu Asn Pro Asn Ala Ile Ala Ser Ala Thr Val Gly Arg Arg Val Ser Ala Arg Met Leu Gly Asp Val Met Ala Val Ser Thr Cys Val Pro Val Ala Pro Asp Asn Val Ile Val Gln Asn Ser Met Arg Val Ser Ser Arg ProGly Thr Cys Tyr Ser Arg Pro Leu Val Ser Phe 601
1714 CCC GTC GCC CCG GAC AAC GTG ATC GTG CAG AAC TCG ATG CGC GTC AGC TCG CGG CCG GGG ACG TGC TAC AGC CGC CCC CTG GTC AGC TTT CCG GTC GCC GCG GAC AAC GTG ATC GTC CGA AAC TCG ATG CGC ATC AGC TCG CGG CCC GGG GCC TGC TAC AGC CGC CCC CTG GTC AGC TTT Pro Val Ala Ala Asp Asn Val Ile Val Gln Asn Ser Met Arg Ile Ser Ser Arg Pro Gly Ala Cys Tyr Ser Arg Pro Leu Val Ser Phe Arg Tyr Glu Asp Gln Gly Pro Leu Ile Glu Gly Gln Leu Gly Glu Asn Asn Glu Leu Arg Leu Thr Arg Asp Ala Leu Glu Pro Cys Thr 631 1804 CGG TAC GAA GAC CAG GGC CCG CTG ATC GAG GGG CAG CTG GGC GAG AAC AAC GAG CTG CGC CTC ACC CGC GAC GCG CTC GAG CCG TGC ACC
CGG TAC GAA GAC CAG GGC CCG TTG GTC GAG GGG CAG CTG GGG GAG AAC AAC GAG CTG CGG CTG ACG CGC GAT GCG ATC GAG CCG TGC ACC ArgTyr Glu Asp Gln Gly Pro Leu Val Glu Gly Gln Leu Gly Glu Asn Asn Glu Leu Arg Leu ThrArg Asp Ala Ile Glu Pro Cys Thr Val Gly His Arg Arg Tyr Phe Ile Phe Gly Gly Gly Tyr Val Tyr Phe Glu Glu Tyr Ala Tyr Ser His Gln Leu Ser Arg Ala AspVal 661 1894 GTG GGC CAC CGG CGC TAC TTC ATC TTC GGC GGG GGC TAC GTG TAC TTC GAG GAG TAC GCG TAC TCT CAC CAG CTG AGT CGC GCC GACIGTCG
GTG GGA CAC CGG CGC TAC TTC ACC TTC GGT GGG GGC TAC GTG TAC TTC GAG GAG TCA GCG TAC TCC CAC CAG CTG AGC CGC GCC GACIATCG
Val Gly His Arg Arg Tyr Phe Thr Phe Gly Gly Gly Tyr Val Tyr Phe Glu Glu Ser Ala Tyr Ser His Gln Leu Ser Arg Ala Asp[le Thr Thr Val Ser Thr Phe Ile Asp Leu Asn Ile Thr Met Leu Glu Asp His Glu Phe Val Pro Leu Glu Val Tyr Thr Arg His Glu Ile 691 1984 ACC ACC GTC AGC ACC TTC ATC GAC CTG AAC ATC ACC ATG CTG GAG GAC CAC GAG TTT GTG CCC CTG GAG GTC TAC ACG CGC CAC GAG ATC
ACC ACC GTC AGC ACC TTC ATC GAC CTC AAC ATC ACC ATG CTG GAG GAT CAC GAG TTT GTC CCC CTG GAG GTG TAC ACC CGC CAC GAG ATC Thr Thr Val Ser Thr Phe Ile Asp Leu Asn Ile Thr Met Leu Glu Asp His Glu Phe Val Pro Leu Glu Val Tyr Thr Arg His Glu Ile Lys Asp Ser Gly Leu Leu Asp Tyr Thr Glu Val Gln Arg Arg Asn Gln Leu His Asp Leu Arg Phe Ala Asp Ile Asp Thr Val IleArgf 721
2074 AAG GAC AGC GGC CTG CTG GAC TAC ACG GAG GTC CAG CGC CGC AAC CAG CTG CAC GAC CTG CGC TTT GCC GAC ATC GAC ACG GTC ATCICGCG
AAG GAC AGC GGC CTG CTG GAC TAC ACG GAG GTC CAG CGC CGC AAC CAG CTG CAC GAC CTG CGC TTC GCC GAC ATC GAC ACG GTC ATC CAC
Lys Asp Ser Gly Leu LeuAsp Tyr Thr Glu Val Gln Arg Arg Asn Gln Leu His Asp Leu Arg Phe Ala Asp Ile Asp Thr Val Ile.His Ala Asp Ala Asn Ala Ala Met Phe Ala Gly Leu C Ala PhePhe Glu Gly Met Gly Asp Leu Gly Arg Ala Val Gly Lys Val Val Met 751
2164 GCC GAC GCC AACGCC GCC ATG TTC GCG GGG CTGITGCG GCG TTCTTC GAG GGG ATG GGG GAC TTG GGG CGC GCG GTC GGC AAG GTA GTC ATG
GCCGAC GCC AAC GCC GCC ATG TTC GCG GGC CTGIGGTIGCG TTT TTC GAG GGG ATG GGC GAC CTG GGG CGC GCG GTC GGC AAG GTG GTG ATG Ala Asp Ala Asn Ala Ala Met Phe Ala Gly LeuGyAla Phe Phe Glu Gly Met Gly Asp Leu Gly Arg Ala Val Gly Lys Val Val Met
Gly VTa Val GlyGly Val Val Ser Ala Val Ser Gly Val Ser Ser Phe Met Ser Asn Pro Phe Gly Ala Leu Ala Val Gly Leu Leu Val 781
2254 GGA|GTA GTG GGG GGC GTG GTG TCG GCC GIL TCG GGC GTG TCC TCC TTT ATG TCC AAC CCC TTC GGG GCG CTT GCC GTG GGG CTG CTG GTC
GGCIATCG GTG GGC GGC GTG GTA TCG GCC GTG TCG GGC GTG TCC TCC TTC ATG TCC AAC CCC TTT GGG GCG CTG GCC GTG GGT CTG TTG GTC
Gly Ile Val Gly Gly Val Val Ser Ala Val Ser Gly Val Ser Ser Phe Met Ser Asn Pro Phe Gly Ala Leu Ala Val Gly Leu Leu Val Leu Ala Gly Leu[Va Ala Ala Phe Phe Ala Phe Arg Tyr Val Leu Gln Leu Gln ArgAsn Pro Met Lys Ala Leu Tyr Pro Leu Thr Thr 811
2344 CTG GCC GGC CTGIGTCG GCG GCC TTC TTC GCC TTC CGC TACGTCICTG CAAICTG CAAICGCG AAT CCC ATG AAG GCC CTG TAT CCG CTC ACC ACC CTG GCC GGC CTGIGCGG GCG GCC TTC TTC GCC TTT CGC TAC GTC |ATG CGGG|CTG CAG AGCG AAC CCC ATG AAG GCC CTG TAC CCG CTA ACC ACC Leu AlaGly Leu Ala Ala Ala Phe Phe Ala Phe ArgTyr Val|Met ArgLeu Gln Ser Asn Pro Met Lys Ala LeuTyrPro Leu Thr Thr
Lys Glu Leu Lys Thr Ser Asp Pro GlyGly Val GlyGly Glu Gly Glu Glu Gly Ala Glu Gly Gly[ly Phe Asp Glu Ala Lys Leu Ala 841
2434 AAG GAA CTC AAG ACTTCC GAC CCC GGG GGC GTG GGC GGG GAG GGG GAG GAA GGC GCG GAG GGG GGCIOGGG TTT GAC GAG GCC AAG TTG GCC AAG GAG CTC AAG AAC CCC ACC AAC CCG GAC GCG TCC GGG GAG GGC GAG GAG GGC GGCGAC TTT GAC GAG GCC AAG CTA GCC
Lys GluLeu Lys|Asn Pro Thr Asn Pro Asp Ala SerGly GluGly Glu Glu Gly Gly[AsJPhe Asp Glu AlaLys Leu Ala
Glu Ala ArgGlu Met Ile Arg Tyr Met Ala Leu Val Ser Ala Met GluArg Thr Glu His Lys Ala|Arg Lys Lys Gly ThrSer Ala Leu 871 2524 GAG GCC CGA GAA ATG ATC CGA TAT ATG GCT TTG GTG TCG GCC ATGGAG CGC ACG GAA CAC AAG GCC|AGA AAG AAG GGC ACG AGC GCC CTG
GAG GCC CGG GAG ATG ATA CGG TAC ATG GCC CTG GTG TCT GCC ATG GAG CGC ACGGAA CAC AAG GCCIAAG AAG AAG GGC ACG AGC GCG CTG Glu Ala Arg Glu Met Ile Arg Tyr Met Ala Leu Val Ser Ala Met GluArg Thr Glu His Lys Ala Lys Lys Lys Gly Thr Ser Ala Leu Leu Ser SerLys Val Thr Asn Met Val Leu Arg Lys Arg Asn Lys Ala Arg Tyr Ser Pro Leu His Asn Glu Asp Glu Ala Gly Asp Glu 901
2614 CTC AGCITCCG AAG GTC ACC AAC ATG GTTICTG CGC AAG CGC AAC AAA GCC AGG TAC TCT CCG CTC CAC AACOAOjGAG GAG GCCGGA GAC GAA CTC AGCGCC AAG GTC ACC GAC ATG GTC ATG CGC AAG CGCOCGC AAC ACC AAC TAC ACC CAA GTT CCC AAC AAA GACGGT GAC GCC GAC GAG Leu SerAlaLys Val Thr Asp Met Val Met Arg Lys Arg|ArgAsn Thr AsnTyr Thr Gln Val Pro Asn[yjAspGly Asp AlaAsp Glu
AspGlu Leu OC 904
2704 GAC GAG GCTC TAA GGGAGGGGAGGGGAGCTGGGCTTGTGTATAAATAAAAAGACACCGATGTTCAAAAATACACATGACTTCTGGTATTGTTTTGCCTTGGTTTlsTATTTGGGGG
GAC GAC CTG TGA CGGGGGGTTTGTTGTAAATAAAAACCACGGGTGTTAAACCGCATGTGCATCTTTTGGTGTGTTTGTTTGGTACGCCTTTTGTGTGTGTGGGAAGAAAGAAAAG Asp Asp Leu OP
2819 GGGGCGTGTGACTAGAAAAACAAATGCAGACATGTGCTAACGGGAAAACCAACCCCAAACCAACCCCAAACCAACCCCGTCTCCCTGCGACCGGTCG CTTTCCAC ACCCCCTCCCCGTG GGAACACATAAACTCCCCCGGGTGTCCGCGGCCTGTTTCCTCTTTCCTTTCCCGTGACAAAACGGACCCCCTTGGTCAGTGCCGATTCCCCCCCACGCCTTCCTCCACGTCGAAGGCTT 2938 GTAGTCTTCCGGGCCTTCCGTCGCGTGTGGGGGCCATCGGTT CGGCTCCTAG CCCCCCCCCCCTCACCCCT CCG ACCT AATTTTTGTGTC ATTCGG CCCA CTTTC CCC CCC ACTCC AC C TTGCATTGTAAAGCTACCCGCCTACCCGCGCCTCCCAATAAAAAAAAAGAACATACACCAATGGGTCTTATTTGGTATTACCTGGTTTATTTAAAAAGATATACAGTAAGACATCCCAT 3057 CCCCCCCTCTCAAACAAAAACACAAGCACACGAAGTGGGTATACTTTTGTCCGGTTGTTTGTTTATTTAAAATATATGAAAACACACACCCCCCCCAAGTCCGGArcc
[image:6.612.62.547.84.720.2]GGTACCAAAGACCGGGGCGAATCAGCGGGCCCCCATCATCTG AGAGACCAACAAATCGGCGG CGCGGGCCGTGTCAACGTrCCACGTCTGCTGCGCTGCTGG CGTTG AC
FIG. 3.-Continued.
on November 10, 2019 by guest
http://jvi.asm.org/
A.
o o cx ~~~MMMmM n R c c Ss
1
11
CCI
I
II
I
Cl
III
1
cI
I
I
T
I
I
~~~II
II
I
0 100 200 300 400 500 600 700 800 900
Amino Acids
B.
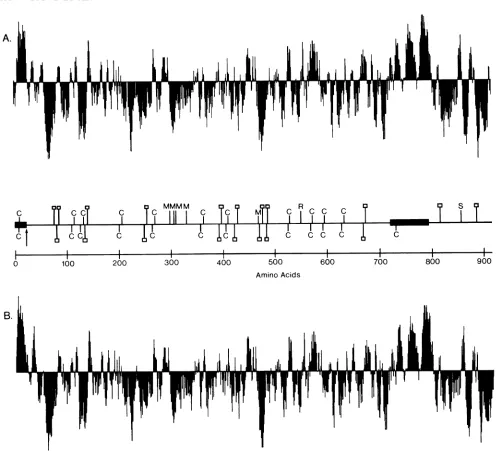
FIG. 4. Hydropathicanalysis ofthegBl (A) and gB2(B)coding regions. Relative hydropathyateachposition is computed by usinga
moving window ofsevenamino acids (15). Symbolsabove the lineindicate increasedhydrophobicity, while those below indicate increasing hydrophilicity. The locations of the predicted signalsequenceandmembraneanchor sequence(solid boxes), potential N-linkedglycosylation sites(O),cysteines (C), rate-of-entry mutation (R)inHSV-1 strainKOS (6), syncytial mutation (S) in HSV-1 (6), and monoclonalantibody resistance mutations(M) inHSV-1(26) areshownonthe middleaxis, withtheHSV-1 locationshown abovetheaxisand theHSV-2location shown below the axis.
ison of the hydropathic structural curves (15) for each protein (Fig. 4). In this analysis, the signal peptide and transmembrane regions wereeasily identified as large hydro-phobicpeaks which rose above the x axis. The similarity of the hydropathic plot for the transmembrane anchor, in particular, reflected the strong homology in this region
(93%).
The conservation of cysteine residues and potentialN-linked glycosylation sites are also indicated in Fig. 4 on the axis between the two curves. With the exception of Cys-733ingB2, 11 of 12 of the cysteines were shared, and all of the predicted glycosylation sites in the external domain
wereconserved.
Additionally, severalregions of the gB protein that appar-entlyhadfunctionalsignificance were also regions where the
moststriking amino acid similaritieswereobserved between gBl and gB2. Bziketal. (7) identifiedamutation in HSV-1 strain tsB5 that is probably responsible for its rapid-entry phenotype relativetoHSV-1strain KOS as a Val (GTC)-to-Ala(GCC) substitutionataminoacid553of gBl (Fig. 4). The gB proteins from both HSV-2 strain 333 and HSV-1 strains Patton and F have an Ala at the homologous position (residue 550 in gB2, 553 in gBl, and 552 in gBl strain F, correspondingtoAla-552of tsB5). As might be expected for
adomainaffectingtherate of virus entryinto the host cell, the51-amino-acid region extendingfrom Ala-524 to Ala-574 of gB2, which is centered around the rate-of-entry locus mapped for gBl, was identified between the type 1 and 2 proteins.
I .
i
.on November 10, 2019 by guest
http://jvi.asm.org/
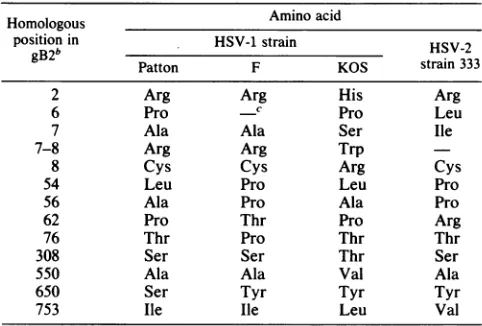
[image:7.612.67.568.56.507.2]STRUCTURE OF HSV-2 gB GENE 333 TABLE 1. Intratypic sequence variation in gBla
Homologous Amino acid
position in HSV-1 strain HSV-2 gB2b Patton F KOS strain333
2 Arg Arg His Arg
6 Pro C Pro Leu
7 Ala Ala Ser Ile
7-8 Arg Arg Trp
8 Cys Cys Arg Cys
54 Leu Pro Leu Pro
56 Ala Pro Ala Pro
62 Pro Thr Pro Arg
76 Thr Pro Thr Thr
308 Ser Ser Thr Ser
550 Ala Ala Val Ala
650 Ser Tyr Tyr Tyr
753 Ile Ile Leu Val
aSummary of the amino acid differences betweengBl strains Patton, F (26), and KOS (6) relative to thehomologous position in gB2 strain 333. For comparison, the amino acid present in gB2 strain 333 is listed in thelast column.
b Forpositions 37 to 52, the sequence for strains Patton andFis
Ala-Ala-Thr-Gln-Ala-Ala-Asn-Gly-Gly-Pro-Ala-Thr-Pro-Ala-Pro-Pro. For strainKOS,
the frame isshifted to read
Arg-Asp-Pro-Gly-Gly-Glu-Arg-Gly-Pro-Cys-His-Ser-Gly-Ala-Ala.The HSV-2 strain 333gB2 hashomology but reads Ala-Ala-Thr-Val-Ala-Ala-Asn-Gly-Gly-Pro-Ala-Ser-Arg-Pro-Pro-Pro.
c-,Space inserted for maximalalignment.
A second mutation responsible for the syncytial (syn) plaquemorphology observedinHELand Vero cells infected with HSV-1 strain tsB5 is most likely due to a transition at nucleotide 2573 that causes an Arg (CGC)-to-His (CAC) amino acid substitution at residue 858 ofgB1 (7). (The syn mutation mapped to amino acid 857 of gB1 from HSV-1 strain tsB5byBzik et al. (7)corresponds toamino acid 858 of gBl from HSV-1 strain Patton.) The location of the homologous region in gB2 is indicated in Fig. 6. The syn locuslay in a stretch of39amino acids, Phe-835 to Ser-873, ofnearly complete identity; only one conservative change was observed between the type 1 and 2 proteins. This was the only extensive region of homology found in the C-terminaldomain ofgBand istherefore likelytobeessential for the membrane fusion function ascribedtothis protein.
ThreeHSV-1 mutants that areresistant to neutralization byoneoftwo type-common monoclonalantibodies directed against gB have been isolated and sequenced (26). The amino acid alterations observed in the mutant gB proteins areindicatedinFig.4. Although multiple amino acid substi-tutions were observed in two of the three mutants se-quenced, it was possible tolocate theepitopic sitesforthe two antibodies on the N-terminal side ofSer-313 and the C-terminal side of Ala-315, respectively. Thus, these two type-common epitopes lay in the most highly conserved region between gBl and gB2, in a stretch of 98 identical amino acidsextending from Ala-235toThr-332,andwithina larger region containing only6changesin 288residuesfrom Phe-108 toThr-395
(98%
homology).Intratypic variation versusintertypic variation. One inter-esting possibilitywasthat theregions ofgreatestdivergence between the type 1 and type 2 gB
proteins
would also correspond to regions ofintratypic
sequence variation. A compilation ofthe amino acid differencesreported
forgBl
between strains Patton, KOS (6), and F (26) is shown in Table 1. Overall, as expected, there was much greater variationbetween the type 1 and 2proteins,with 125amino acid differences or substitutions, compared with
only
29amino acid differences among the three type 1 sequences (including one large frame shift in KOS relative to strains Patton and F which accounted for 15 of the 29differences). The distribution ofthesedifferences was veryassymmetric, with all but four located within the first 76 amino acids. There were five substitutions and the frameshift in the amino-terminal region of the protein, an area of very high divergence between the type 1 and type 2 proteins. Of the fourremaining substitutions in therestof the protein, all of which lay in regions quite conserved between the type 1 and type 2 sequences, three were conservative amino acid re-placements. No intratypic divergence occurred in thecentral
orcarboxyl-terminalintertypic variant regions. We note that it is possible that some of theintratypic variation may arise through DNA sequencing errors, since DNAs of very high G+C basecomposition are notoriously difficult to sequence. Thus,weobservedboth intra- and intertypic variation at the amino terminus of gB. However, the remainder of the protein was highly conserved between the type 1 isolates, even though twoadditional variable regions were apparent aftercomparison of type 1 and type 2 proteins.
Expression of gB2 in mammalian cells. Glycoprotein gB2 wasengineered for expression in cells to determine the type specificity of the host immune response against gB. No convenient restriction sites were available within the 5' untranslated mRNA leader of the gB2 gene. Therefore, a
uniqueHindIII site was introduced by in vitro mutagenesis,
asdescribed inMaterials andMethods,byconversion of the PstI site atnucleotide -61.
ThegB2 genewasexcisedfrom themutagenized M13 RF derivative phage as a 1.90-kb HindIII-PvuII fragment and inserted into the mammalian expression vector pSVldhfr (24) in place of the dhfr gene by ligation to a 4.85-kb HindIII-BglII fragmentof the vectorinwhich theBglII site had been repaired to a blunt end by using the Klenow fragment of DNA polymerase I. The resultant plasmid, pHS210 (Fig. 5), contained 55 base pairs of 5' untranslated HSV-2 sequence upstream of thegB2 coding region. Tran-scription of gB2 is driven by the simian virus 40(SV40) early promoter, andthe 3' spliceand polyadenylation signalsare
also provided by SV40 sequences. Additionally, pHS210 contains the origin ofreplication and the ampicillin
resist-ance gene from pBR328 required for the replication ofthe plasmid in E. coli. The gB2 coding regionis truncatedat a
PvuII site, 110 amino acids N-terminal to the proposed membrane anchor sequence. The truncated gene thus
en-codesaproteinthatlacks thetransmembrane and C-terminal sequences ofgB2, and the encoded protein should be
se-creted into the medium afterexpressionin mammalian cells. DNA sequence analysis predicted that the translation of gBlin thepHS114construction would terminateatthe small Tstopcodonwithin theSV40sequencesdownstreamofthe gBl gene (24). Dueto the similarity of truncation sites and
vectorsplicesites between thepHS114 gBl constructionand thepHS210 gB2 construction described here, translationof gB2 presumably terminates at the same position. After expression in mammalian cells, the truncated gB2 protein shouldcontain 637 aminoacids,beforecleavageof thesignal peptide, including24C-terminal amino
acids,
due tofusion withSV40 smallTantigen.Toverifythe
production
ofgB2protein
from this vector, pHS210 was introduced into COS 7 cells (13)by
calcium phosphate-mediatedtransfection(41). The transient expres-sionofgB2proteinwas detected in fixed cellsby
immuno-fluorescence withananti-HSV-2 rabbit antiserum (datanot shown). After evaluation in COS 7cells, pHS210
was61,
on November 10, 2019 by guest
http://jvi.asm.org/
E p
r SV4O B pBR328
AP
IPo,
11~~~~ S
Ps Ad-dhfr(64210SV4oo
pBR322 -2 Bg B 1SV40 5pHS 2 Poly B
4/ frAeP 4
3Ad-2 3
ML9 ~ H
(S)(X)
Pv-i
Bg
FIG. 5. Mamalian cell expression vectors for DHFR and gB2.
Plasmid Ad-dhfr(6.44 kb) consists of the originofreplication and the ampicillin resistance (Ampr) gene of pBR322 (as a 3.7-kb
EcoRI-Sallrestrictionfragment)followedbytheadenovirustype2
majorlatepromoter(Ad-2 MLP)asanXhol-Hindlll fragment(map
units 16to 17.3) (4)fused tothe 0.72-kbHindlll-BgllI mousedhfr
cDNA(VI ) (23). dhfr is followed by the DNA coding for theintron
for SV40 small Tantigen (map units 0.56to0.44)(5) and the SV40
sequences between map units 0.19 and 0.00 (EcoRl site) which
include the early region polyadenylation site (poly A) at position 0.155(5). pHS210 (6.79 kb) is based onthevectorpSVldhfr (24). The HindIII-BglII fragment encoding dhfr has been replaced witha
1.9-kb HindIlI-PvuII fragment encoding a truncated gB2 protein.
, Transcripition is driven by the simian virus 40 earlypromoter
(SV40 EP). Restriction sites: B, BamHI; Bg, BglII; E, EcoRl; H, HindlIl, P, PstI; Pv, PvuIl; S, Sall; X, XhoI. Sites listed in
parentheseswereusedin thecloning butwerenotregenerated.
cotransfected into CHOcells (38) with theAd-dhfr plasmid (Fig. 5), which includes the selectable and amplifiable marker DHFR (1, 23). Following transfection and propaga-tion in selective medium, 84 DHFR+ clones were isolated and screened for synthesis and secretion of gB2 with an ELISA. Of the DHFR+ clones, 19% secreted detectable amountsof gB2 (>1.5 ng/ml). Clone pHS210 3-1, which had the highest levels of gB secretion, was chosen for further characterization of the gB2 polypeptide. The gB2 protein
was detected by labeling with [35S]methionine followed by
1 3 4
Kd
200 -925-.)..
9-2_
184
FIG. 6. Radioimmunoprecipitation of intracellular and secreted
gB2protein. Lane 1, Size standards: myosin (200 kDa),
phosphor-ylase b(92.5 kDa), bovine serumalbumin(68kDa), ovalbumin (43
kDa), chymotrypsinogen(25.7kDa), and fI-lactoglobulin(18.4kDa). Lanes 2 and 3, Immunoprecipitation of cell lysates from a 1-h
pulse-labeling of pHS210-transfected cell line 3-1 (lane 2) and an Ad-dhfr-transfected control cell line (lane 3). Lanes 4 and 5,
Immunoprecipitation of conditioned medium from a 1-h pulse
fol-lowedbya5-h chase for pHS210-transfectedcell line 3-1 (lane 4) and anAd-dhfr-transfected control cell line (lane5).
radioimmunoprecipitation. Aftera1-hpulse,diffusedoublet bandscorrespondingtopolypeptides of79 and 84kilodaltons (kDa) were detected intracellulary (Fig. 6, lane 2). These proteins arelarger than the68,991-dalton size predicted for the 637-residue truncated gene product and presumably correspondtopartially glycosylated precursors.Aftera 5-h chase, nogB2wasdetectedintracellularly (datanotshown), and an89-kDapolypeptidewasdetectedin themedium(Fig. 6, lane 4). The size of the mature, fully glycosylated gB2 secreted into the medium by clone pHS210 3-1 was
some-what smaller than the 100-kDa gB1 secreted by pHS114-6 (24) due to theremovalfrom pHS210 ofthecodingsequence for 94 amino acids included in the gBl
plasmid.
Approxi-mately4% ofthe intracellular gB2 labeled in the 1-h pulse wasfoundsecreted intothemedium afterthe 5-h chase.This lowsecretionefficiencywassimilartothatobserved forgBl with clonepHS114-6 (24).
DISCUSSION
We havedetermined the sequenceoftheHSV-2 glycopro-teingB andcomparedthe deduced aminoacidsequencewith that of the corresponding HSV-1 gB
protein.
The two proteinsshowstriking homology,aswouldbeexpected foraviralglycoproteinwithacentral role in virusattachmentand entry into the infected cell. The DNA sequences are 86% homologous, and the predicted amino acid sequences are
also86% homologous. Severalregions oftheprotein, includ-ingacentral domainand themembrane anchordomain,are even morehighlyconserved. Mutations ingBlmutantsthat were selected for resistance to type-common, neutralizing monoclonal antibodies (26) lie within a highly conserved central domain. In addition, twofunctional gB1
mutations,
onethat alters the rateofvirusentryinto theinfected cells anda second thatcausestheformation of
syncytial
plaques in infected cell monolayers (7), also mapped to verycon-served regions ofthe protein.
However, thetwoproteinscancertainlybedistinquished by specific antibodies (11, 27),andcorrespondinglythereare regions of marked divergence. The most striking of the clusteredaminoacid substitutions occurswithin the amino-terminal 85 amino acids of the mature protein, while a
second region is located within the interior of the
protein
including amino acids 451to495. These regions mayplaya
role in the immunological differentiation between the two proteins but are unlikely to correspond to functional do-mains due to their divergence. The amino-terminal variant region alsocorrespondsto aregionofintratypic divergence between strains oftype 1.
We have also produced a truncated form of the gB2 protein by transfection of mammalian cell lines with gB2 geneengineeredintoanappropriate expressionvector.This protein was recognized by a gB-specific, type-common monoclonal antibodyF3A/B (31), inacapture ELISA (data
not shown), provingthat the DNA sequencedescribed here encodes gB2. These stable cell lines, which secrete gB2, should enableus toproduce and purify sufficient amounts of the gB2 to aid in elucidating the immune response of the infectedhost.
ACKNOWLEDGMENTS
We thank Mary Alice Malkemus and Paul Laybourne for their technical assistance, Charles Stabenforhis helpful comments, and DanaTopping, and Pauline Montesfor careful preparation ofthis manuscript.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.73.308.54.190.2] [image:9.612.143.238.480.633.2]STRUCTURE OF HSV-2 gB GENE 335 LITERATURE CITED
1. Alt, F. W., R. E. Kellems, J. R. Bertino, and R. T. Schimke. 1978. Selectivemultiplication ofdihydrofolate reductasegenes in methotrexate-resistant variants ofcultured murine cells. J. Biol. Chem. 235:1357-1370.
2. Barr, P. J., R. M. Thayer, P. Laybourn, R. C. Najarian, F. Seela, and D. R. Tolan. 1986. 7-Deaza-2'-deoxyguanosine-5'-triphosphate: enhanced resolutionin M13 dideoxy sequencing. BioTechniques 4:428-432.
3. Benton, W. D., and R. W. Davis. 1977. ScreeningXgt recombi-nant clones byhybridization to singleplaques in situ. Science 196:180.
4. Broker, T. R. 1980. Nucleotide sequences, transcription and translation analyses, and restriction endonuclease cleavage maps ofgroup-C humanadenoviruses,p.937-1002. In J. Tooze (ed.),DNAtumorviruses, 2nd ed., part 2, revised. Cold Spring HarborLaboratory, Cold SpringHarbor, N.Y.
5. Buchman, A. R., L. Burnett, and P. Berg. 1980. The SV40 nucleotidesequence, p.779-841.In J.Tooze (ed.), DNA tumor viruses, 2nd ed., part 2, revised. Cold SpringHarbor Labora-tory, Cold Spring Harbor,N.Y.
6. Bzik, D. J., B. A. Fox, N. A. DeLuca, and S. Person. 1984. Nucleotide sequence specifying the glycoprotein gene, gB, of herpes simplex virus type 1.Virology133:301-314.
7. Bzik, D. J., B. A. Fox, N. A. DeLuca, and S. Person. 1984. Nucleotide sequenceofa region of herpes simplex virus type 1 gBglycoproteingene:mutations affectingrate ofvirus entry and cell fusion. Virology 137:185-190.
8. Chou, P. Y., and G. D. Fasman. 1978. Prediction of the secondarystructure ofproteinsfrom their amino acidsequence. Adv. Protein Chem. 47:45-148.
9. Corey,L., andP. G. Spear. 1986.Infectionswithherpes simplex virus. N. Engl. J. Med. 314:686-691.
10. Cortini, R., and N. M. Wilkie. 1978. Physical maps for HSV type 2 DNA with five restriction endonucleases. J. Gen. Virol. 39:259-280.
11. Eberle,R., and R. J.Courtney. 1981. Assay oftype-specificand type-common antibodiestoherpes simplex virus types 1 and 2 in human sera. Infect. Immun. 31:1062-1070.
12. Engelman, D. M., and T. A. Steitz. 1981. The spontaneous insertion of proteins into and across membranes: the helical hairpin hypothesis. Cell23:411-422.
13. Gluzman, Y. 1981. SV40-transformed simian cells support the replication of earlySV40mutants. Cell 23:175-182.
14. Holland, L. E., R. M. Sandri-Goldin, A. L. Goldin, J. C. Glorioso, and M. Levine. 1984. Transcriptional and genetic analysesof theherpessimplex virus type 1genome: coordinates 0.29 to 0.45. J. Virol.49:947-959.
15. Hopp, T. P., and K. R. Woods. 1981. Prediction of protein antigenic determinantsfrom amino acidsequences. Proc. Natl. Acad. Sci. USA78:3824-3828.
16. Hubbard,S. C., and R. J. Ivatt. 1981.Synthesisandprocessing of asparagine-linked oligosaccharides. Annu. Rev. Biochem. 50:555-583.
17. Kudler,L., T. R. Jones, R. J.Russell,and R. W.Hyman. 1983. Heteroduplex analysis ofcloned fragments of herpes simplex virus DNAs. Virology 124:86-99.
18. Laemmli,U. K. 1970.Cleavage ofstructuralproteins duringthe assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
19. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. ColdSpring HarborLaboratory,
Cold Spring Harbor, N.Y.
20. Matteucci, M. D., and M. H. Caruthers. 1981. The synthesisof deoxyoligonucleotides on a polymer support. J. Am. Chem. Soc. 103:3185-3191.
21. McMaster, G. K., and G. G. Carmichael. 1977. Analysis of singleanddouble-stranded nucleic acidsonpolyacrylamideand agarose gels byusing glyoxal andacridine orange. Proc. Natl. Acad. Sci. USA74:4835-4838.
22. Mills, D. R., and F. R. Kramer. 1979. Structure-independent
nucleotide sequence analysis. Proc. Natl. Acad. Sci. USA76:
2232-2235.
23. Nunberg, J. H.,R.J. Kaufman,A.C. Y. Chang,S. N. Cohen, and R. T.Schimke. 1980.Structure and genomic organization of themousedihydrofolatereductase gene. Cell 19:355-364. 24. Pachl, C., R. L. Burke, L. L. Stuve, L. Sanchez-Pescador, G.
VanNest, F. Masiarz, and D. Dina. 1987. Expression of cell-associated and secreted forms of herpes simplex virus type 1
glycoproteingB inmammaliancells. J.Virol. 61:315-325. 25. Pachl, C., W. Schubach, R. Eisenman, and M. Linial. 1983.
Expression of c-myc RNA in bursal lymphoma cell lines: identification of c-myc encoded proteins by hybrid-selected translation. Cell 33:335-344.
26. Pellett, P. E., K. G. Kousoulas, L. Pereira, and B. Roizman. 1985. Anatomy of the herpes simplex virus type 1 strain F glycoprotein B gene: primary sequence and predicted protein
structureof the wild type and of monoclonal antibody-resistant
mutants.J. Virol. 53:243-253.
27. Pereira, L., D. V. Dondero, D. Gallo, V. Devlin, and J. D. Woodie.1982. Serological analysisof herpes simplex virus types 1and2 withmonoclonal antibodies. Infect.Immun. 35:363-367. 28. Perlman, D., and H. 0. Halvorson. 1983. A putative signal peptidase recognition site and sequence in eukaryotic and prokaryotic signal peptides. J. Mol. Biol. 167:391-409. 29. Proudfoot, N. J., and G. G. Brownlee. 1976. 3' Non-coding
region sequences in eukaryotic messenger RNA. Nature
(Lon-don)263:211-214.
30. Ratfield, L. F., and D. M.Knipe. 1984. Characterization of the major mRNAs transcribed from the genes for glycoprotein B andDNA-binding protein ICP8 of herpes simplex virus type 1. J. Virol. 49:960-969.
31. Rector, J. T., R. N. Lausch, and J. E. Oakes. 1982. Use of monoclonal antibodies for analysis of antibody-dependent im-munity to ocular herpes simplexvirus type 1infection. Infect. Immun. 38:168-174.
32. Roizman, B. 1979. The structure and isomerization of herpes
simplexvirus genomes. Cell 16:481-494.
33. Ruyechan, W. T., L. S. Morse, D. M. Knipe, and B. Roizman. 1979.Moleculargeneticsofherpes simplex virus.II.Mappingof themajor viral glycoproteins and ofthe geneticloci specifying the social behavior ofinfected cells. J. Virol. 29:677-697. 34. Sanger, F., S.Nicklen, and A. R.Coulson. 1977. DNA
sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
35. Seela, F., Q. Tran-Thi. and D. Frazen. 1982. Poly(7-deazaguanylic acid), the homopolynucleotide of the parent nucleoside of queuosine. Biochemistry21:4338-4343.
36. Spear, P. G. 1985. Glycoproteins specified by herpes simplex viruses, p. 315-356. In B. Roizman (ed.), The herpesviruses, vol. 3. PlenumPublishing Corp., New York.
37. Thomas,P. S. 1980.HybridizationofdenaturedRNA andsmall DNAfragments transferredtonitrocellulose. Proc. Natl. Acad. Sci. USA77:5201-5205.
38. Urlaub, G., and L. A. Chasin. 1980. Isolation of Chinese hamstercell mutants deficient in dihydrofolate reductase activ-ity. Proc. Natl. Acad. Sci. USA 77:4216-4220.
39. Vahlne, A., B. Svennerholm, and E. Lycke. 1979. Evidence for herpes simplexvirustype-selectivereceptors oncellular plasma membranes.J. Gen. Virol. 44:217-225.
40. Vahlne, A., B. Svennerholm, M.Sandberg, A.Hamberger, and E. Lycke. 1980. Differences in attachment between herpes simplex type 1 and type 2 viruses to neurons and glial cells. Infect. Immun. 28:675-680.
41. van der Eb, A.J., and F. L. Graham. 1980. Assayof transform-ing activity oftumor virus DNA. Methods Enzymol. 65:826-839.
42. von Heijne, G. 1984. How signal sequences maintain cleavage specificity. J. Mol. Biol. 173:243-251.
43. Watson, M. E. E. 1984. Compilation of published signal se-quences. Nucleic Acids Res. 12:5145-5164.
44. Zolier, M. J., and M. Smith. 1983. Oligonucleotide-directed mutagenesis of DNAfragments cloned into M13vectors. Meth-odsEnzymol. 100:468-500.
61,