0022-538X/89/051877-07$02.00/0
Copyright © 1989,AmericanSociety forMicrobiology
Short, Duplicated Sequence Indicative of
the
Recombinogenicity of
the Junction between
a
Unique
and
an
Inverted
Repeat
Sequence
in
the S
Component of the
Herpes
Simplex Virus
Type 1
Genome
KENICHI UMENE
Department of Virology, Faculty of Medicine, Kyushu University 60, Fukuoka 812, Japan Received16September 1988/Accepted29December1988
A herpessimplex virustype1(HSV-1) strain, B3,wasfoundtohaveashortduplicationonthe left junction
between theuniquesequence (Us)and the inverted repeatsequence
(Rs)
in theS componentof thegenomeDNA. A shortregion of
Rs
contiguoustotheleftUs-Rs
junctionwasduplicatedinB3. Basedonthe nucleotide sequencesin and around theUs-Rs
junctions of B3and otherHSV-1 strains,aconceptofjunctionstretchwasproposed. Theorganization of junctionstretch is
Rs
side5'-(GorAstretch)AGC-3'Us
side.Introductionof theconceptofjunctionstretch ledtoadefinition ofthestructurein and around theUs-Rs
junction,in theformcommontoHSV-1 strains. The rightendof
Us
inthe HSV-1 genome wasthe A of theATG initiationcodonofgeneUS12, and thus theATGtriplet may actas a bufferto preventexpansion of
Rs,
asisthecasewithHSV-2. The duplication inB3wasgeneratedbya crossovereventbetweenapointon
Rs
andtheUs
sideendofthe left junctionstretch.These observationssuggestthat the
Us
side endofthejunctionstretchpossessestheproperty of recombinogenicity, responsible for generation ofthe duplication in strain B3 and also for the formation of the
Us-Rs
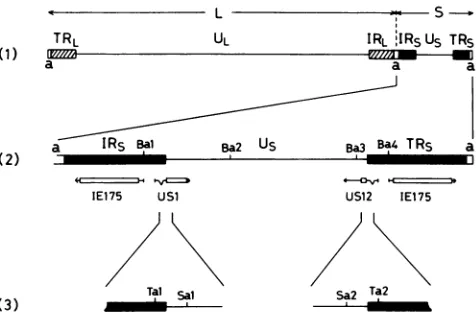
junction of HSV.Three alphaherpesviruses ofherpes simplexvirus types 1 and2(HSV-1 andHSV-2)andvaricella-zoster virus (VZV) arecommonhumanpathogens (7, 8, 19). They have aDNA genomecontaining twouniquesegments(ULand Us), each of which is bounded by its own inverted repeats (TRL and IRL, and
IRS
andTRs)
to form the long (L) and short (S) components(4, 18)(Fig. 1). HSV-1 and HSV-2aresimilar in both genome structure and gene layout. The other alpha-herpesvirus, VZV, differssignificantlyfrom the herpes sim-plex viruses in genome size and base composition. Exami-nations ofthepolypeptide sequencehomology showed that the S components of HSV-1 and VZV are related to asignificant degree, though they do differ in genelayout (3). Itishypothesizedthatarcombinationbetweentwounique sequencesaligned in an opposite orientationcaused expan-sion of the inverted repeat and contraction of the unique sequence in herpesvirus genomes (7, 32). This proposal seemed to account for three alphaherpesviruses HSV-1, HSV-2, and VZV as having descended from a common ancestor. Based on this same idea, Davison and McGeoch presenteda scheme toshow the derivation of the S compo-nents of HSV-1 and VZV from the S component of an ancestral herpesvirus by expansion and contraction of
Rs
(3). The scheme satisfactorily explained the differences between HSV-1 and VZV in gene layout. HSV variants, which were assumed to be generated by recombination between uniquesequences arranged in anopposite orienta-tion withexpanded repeat sequence and contracted unique sequence, were actually isolated (1, 11, 23). The
Us-Rs
junction corresponds to the crossover point. As a factor involved in the determination of theUs-Rs
junction, Whitton and Clements noted the location of the first translational initiationcodon (32). Thetranslational initiation codon of the IEmRNA-5gene(corresponds to gene US12) of HSV-2 had the Aof the ATGasthefirst base inUs,
andthisdefined the rightUs-Rs
junction. They assumed that the ATG of the HSV-2 IEmRNA-5 gene acted as a buffer to prevent the expansionofTRs.
However, the first ATG of theIEmRNA-5 geneof HSV-1 strain 17was8 basepairs (bp)into
Us
and didnotdirectlydefine therightUs-Rs
junction (32).I have now obtained evidence fora novel HSV-1 strain, termedB3, which hasashortduplicatedregionderived from
Rs
on the leftUs-Rs
junction. The nucleotide sequencesaround
Us-Rs
junctionsof B3 and other HSV-1 strainswere compared. A concept ofjunction stretch was introducedto define thestructurearoundtheUs-Rs
junctionsin the form common toanyHSV-1 strain. The rightend ofUs
of HSV-1 defined in this formwasthe A ofthefirstinitiator ATG ofthe US12 (IEmRNA-5) gene. The ATG of IEmRNA-5 gene seemed to act as abufferto preventthe expansionofRs
in the HSV-1 genome, as was the case with HSV-2. The duplicationin B3 was assumed to begenerated by a cross-over eventbetweenapointinR.
and theUs
side end of the leftjunction stretch. Thus, theUs
side end of thejunction stretch apparently possesses the property of recombinoge-nicity, responsible forgeneration oftheduplicationin strain B3 andalsofor the formation of theUs-Rs
junctionof HSV.MATERIALS ANDMETHODS
Cellsand viruses. Vero cellsweregrown inEagleminimal essential medium(MEM) supplementedwith 5% calfserum.
SP23, used as the standard HSV-1 in this work, was a
single-plaque isolatefrom HSV-1 strainPatton (27). HSV-1 strain B3, the duplication of which was analyzed in this work, and HSV-1 strains B4 and B9 wereisolatedatKyushu University Hospital (25).
Working stocks of HSV-1 were made on Vero cells in EagleMEMwith2% fetal bovineserum at alowmultiplicity of infection (0.01 PFU per cell). HSV-1 DNAs were pre-pared from viral particles obtained after glycerol gradient centrifugation (23).
Restriction endonuclease digestion, acrylamide gel electro-phoresis, and Southern hybridization. Restriction endonu-cleaseswere purchased from Takara ShuzoCompany (Ky-oto, Japan) and Toyobo Company (Osaka, Japan), and conditions for digestion were those recommended
by
the 1877on November 10, 2019 by guest
http://jvi.asm.org/
(1)
(2)
TRL UL IRL IRSUs TRs
a a a
aIR5Bal Ba2 U5 Ba3 4TR5
IE175 USi US12 IE175
Tal sal Sa Ta2
FIG. 1. Maps of HSV-1 genome. (1) Structure of the HSV-1 genome arranged in prototype orientation (18). HSV-1 DNA is a linear, double-stranded molecule of about 155 kb, consisting of two covalentlylinked components, L and S, that constitute 82 and 18% ofthe genome, respectively. Each component consists of unique sequences (UL and Us) bracketed by inverted repeat sequences (TRL, IRL,IRs,andTRS).Ashortsequence, a, is repeated directly atthe terminiofthe HSV-1 genome and is also present in the inverse orientationat the L-Sjunction. (2) Expansion of the S component. The locations and 5' to 3' orientations of mRNA species are indicatedbyhorizontal arrows. Protein-coding regions are shown as open boxes (13, 14, 17).BarmHI sites(Bi toB4)areindicated (26). (3) Expansions of the regions spanning the Us-Rsjunctions. TaqI sites (Tal and Ta2) and Sau3AI (isoschizomer of Mbol) sites (Sal andSa2) are indicated (13, 14).
manufacturers. The DNAs digested with restriction endonu-cleaseswereseparated in a5% acrylamide gel(22).Southern hybridization was carried out on a Biodyne A transfer membrane (Pall Ultrafine Corp.) asdescribed before (22).
DNAsequencing. An appropriaterestriction fragment was subcloned into both M13mplO and M13mpll and sequenced by usingthedideoxynucleotidechain termination procedure (15, 20).
RESULTS
Detection of an HSV-1 strain with a variation around the left
Us-Rs
junction. Restriction fragment length polymorphism of HSV-1 strains wasanalyzed by using restriction endonu-cleasesrecognizing4-bp motifs (25). It became clear that the HSV-1 strain termed B3 differed from other HSV-1 strains with respect to the length ofrestriction fragments deriving from the region spanning theUs-Rs
junction. The DNAs of strain B3 and strain SP23 (asastandard)weredigested with TaqI andMboI (isoschizomer ofSau3AI), electrophoresed in a 5% acrylamide gel, and transferred to a nylon mem-brane. The DNAs on the membrane werehybridizedwitha32P-labeled 0.14-kilobase (kb) TaqI-Sau3AI DNA fragment containingtheright
Us-Rs
junction, which corresponded to the Ta2-Sa2fragment inFig. 1(lanes 1 and 2 in Fig. 2). The fragment of about 0.14 kb, corresponding to the region containingtherightUs-Rs
junction (Ta2-Sa2 in Fig. 1), was detected in thedigested products of both B3 and SP23. The 0.15-kbfragment, correspondingto theregion containing the leftUS-Rs
junction [Tal-Sal in Fig. 1 (3)], was detected in the digests of SP23 but was not present in those of B3. A fragmentof 0.20 kbwasdetected in thedigested products of B3. Therefore, a structural variation around the leftUs-Rs
junction is present inthe B3genome.
Structuresof theregions spanning
Us-Rs
junction of HSV-1 genomes. TheEcoRIHfragment of strain B3, which contains1 2 3 4 M
-0.271 -0.234
- MO -0.194
-0.118
-0.072
FIG. 2. Southernhybridization analysesof DNAfragments con-taining Us-Rs junctions of HSV-1 genomes. Genome DNAs of HSV-1 strains SP23 (lane 1) and B3 (lane 2), 1- to 3-kb BamHI fragmentsof strainB3(containingrightUs-Rsjunction) (lane 3),and 4- to 7-kb BamHI fragments of strain B3 (containing left Us-Rs junction)(lane 4) weredigested withTaqI and MboI.Thedigested DNAswere electrophoresed in an5%acrylamide gel, transferred to a Biodyne A transfer membrane, and hybridized with 32P-labeled 0.14-kb Taq1-Sau3AI DNA fragment containing right Us-Rs junc-tionof HSV-1 strain Patton[Ta2-Sa2 in Fig. 1 (3)]. Lane M, Marker mixture ofHaellI fragments of (X174 DNA. Sizes of the fragments areshown in kilobases.
allof
Us
and a part ofRs,
wascloned into XgtWES (26).Therecombinant phage was termed XEH3. The two TaqI-Sau3AI fragments of 0.14 kb and 0.20 kb, which were assumed to correspond to the regions containing the right
and left
Us-Rs
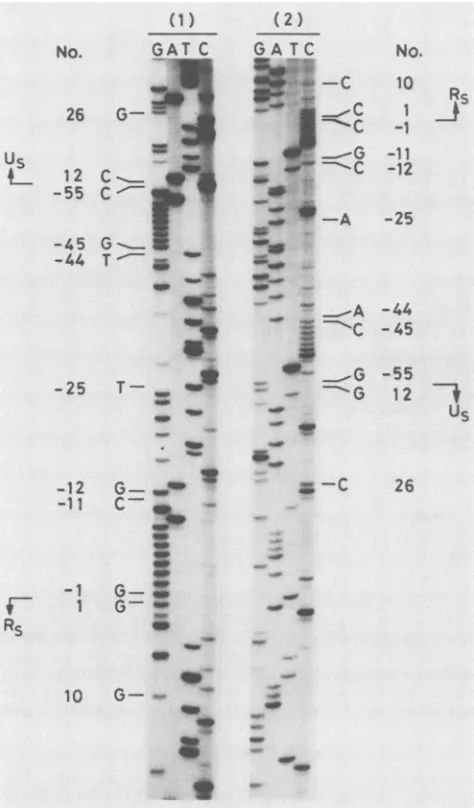
junctions, respectively, were prepared fromXEH3 DNAs and cloned into both M13mplO and M13mpll. Thenucleotidesequencesof the fragments weredetermined by usingthedideoxynucleotide chain termination procedure (Fig. 3 and 4). In the 0.20-kb fragment containing the left
Us-Rs
junction of B3, a duplication of about 40 bp was identified. The presence of the duplication was assumed to be the cause ofelongation of the region spanning the leftUs-Rs
junctionin B3(lane 2 in Fig. 2).The known nucleotide sequences around the
Us-Rs
junc-tions of strains 17 and Patton werecompared [Fig. 5 (1 and 2)] (13, 14, 16, 31). With respecttothenucleotide sequence on theRs
side, strain 17 was similar to strain Patton. However, there weredifferences in the nucleotide sequence ontheUs
side. The first threeorfour residueson theright end ofUs
differed between the two strains [Fig. 5(1R and 2R)]. The A of initiation codon ATG of gene US12 is the eighthresiduefromtherightUs-Rs
junctionin strain 17 and theseventh residue in strain Patton[Fig.5(1R and2R)]. TheUs-Rs
junctionin thesestrainswasdeterminedsimplyasthe point over which the homology between the twoRs
se-quenceswas nolonger maintained. To search forregulation in thestructureofUs-Rs
junctions,thenucleotide sequences around theUS-Rs
junctions oftwoother HSV-1 strains, B4 and B9, were determined [Fig. 5 (3 and 4)] and compared with those of strains 17and Patton. If theUs-Rs
junctionsare definedas the pointover which the homology between thetwo
Rs
sequenceswas nolongermaintained,thelocation of thejunction would not be in a form common to HSV-1 strains.Thus,Iintroduced the concept ofjunctionstretchto define theregions spanning theUs-Rs
junctions in the form commonto anyHSV-1 strain. Construction of thejunctionon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.65.303.73.229.2] [image:2.612.379.505.76.241.2](1) (2)
No. GAT C G A T C No.
*ws
;
W10
26 G-- 1
G -11
U
12
C_C..-*
-12-55 C- _
* _ -~~A-25
-45 G_.
-44
T---_ _A -44
-- -C -45
-25T--~ump* G -5 5
-25
T-_
--
-G
12_~
-12 G-_ S-C 26
-11 C _ _
-1 G-
-r
1 G--_Rs
10
G-Rs
us
FIG. 3. Autoradiographs showing nucleotide sequences of the
novelduplicationonthe leftUs-Rsjunction of HSV-1 strain B3. The
0.20-kb TaqI-Sau3AI fragmentofXEH3, containingthe left Us-Rs junction of strain B3, was cloned in M13mplO (panel 1) and
M13mpll(panel 2) andsequenced bythedideoxynucleotide chain termination method. The products were separated in a thin 6%
polyacrylamide-urea gel. The nucleotide numberswerederivedby comparisonwith thesequencereported byMcGeochetal.(13, 14) with the referencepoint atthe left Us-Rsjunction. The regions of
novelduplication and junction stretchwerenumberedwithaminus (-1to -55).The concept ofjunction stretchwasproposedin this
work, and the detailsaredescribed in thetext.
stretch is
Rs
side5'-(GorAstretch)AGC-3'Us
side[Fig. 5 (5)].Thelengthof the GorAstretch variedfrom 3bp (rightUs-Rs
junctionof strainB9)to 10bp (leftUs-Rs
junctionof strainPatton) in the four HSV-1 strains examined (Fig. 5). ThejunctionstretchwasconsideredapartofRs
withregard to derivation. The nucleotides in thejunction stretch werenumbered and extended from the residuecontiguous to
Rs
[numbered -1 inFig. 5(5Land5R)]. Divergenecefrom theconsensus sequence of thejunction stretch was found at residue 1 in the left
Us
side of strain 17[Fig. 5(1L)] andatresidue 4 in the left
Us
side of strain B4 [Fig. 5 (3L)]. The nucleotide sequences encompassing the junction stretchwere conserved among the HSV-1 strains and could be numbered
uniformly.
TheRs
side sequenceadjoining
thejunction
stretchstarted from G[numbered
1 inFig.
5(5L
and SR)]. TheUs
side sequenceadjoining
the leftjunction
stretch started from C
[numbered
12 inFig.
5(5L)]
and ended atA[numbered
12972 inFig.
5(5R)]
ontheopposite
side of
Us
adjoining
theright junction
stretch. The number-ingsystemforRs
andUs
wasthe same asthat of McGeoch etal.(13)and McGeochetal.(14),respectively.
Theregions
which had been regarded as terminal parts of
Us
in theprevious
definitionwere converted into thejunction
stretch in this work(Fig.
5). This alterationplaced
the A of translationalinitiation codon ATG of gene US12at the first baseinUs
from theright junction
stretch.Fixation of theduplicated regionontheleft
Us-Rs
junction
in strainB3.Nucleotidesequenceanalyses
ofarecombinant phage clonecarrying
the EcoRI Hfragment
of strain B3 indicated the presence oftheduplicated
region
on the leftUs-Rs
junction
ofstrain B3(Fig.
3 and4).
To confirm thefixation ofthe
duplication
on the leftUs-Rs
junction,
B3 genome DNAs wereanalyzed directly.
DNA of strain B3 was digested with BamHI and electrophoresed in a 0.8% agarosegel.
Two agarosegel
blocks, containing
the BamHIfragments
of1to3kb[including
the2.0-kbBamHIfragment
spanning
theright
Us-Rs
junction (Ba3-Ba4)
inFig.
1(2)]
and the BamHI
fragments
of4to7 kb[including
the5.5-kb BamHIfragment spanning
theleftUs-Rs
junction (Bal-Ba2)
in
Fig.
1 (2)],respectively,
wereprepared.
DNAsextracted from each agarosegel
block weredigested
withTaqI
andMboI,
electrophoresed
ina5%acrylamide
gel,
transferredto a nylonmembrane,
andhybridized
with the 32P-labeled 0.14-kbTaqI-MboI
fragment
containing
theright
Us-Rs
junction (lanes
3 and 4 inFig. 2).
The 0.14-kbfragment,
which was assumed to spanthe
right
Us-Rs
junction,
was detectedonly
indigests
ofDNAsfromthe agarosegel
blockcontaining
the BamHIfragments
of 1 to 3 kb(including
Ba3-Ba4
fragment
withright
Us-Rs
junction).
The 0.20-kbfragment having
theduplicated region
wasdetectedonly
in thedigests
ofDNAs from the agarosegel
blockcontaining
the BamHI
fragments
of 4 to 7 kb(including
Bal-Ba2fragment
withleftUS-Rs
junction).
Itwasconcluded that theduplicated region
ofstrain B3 was fixed at the leftUs-Rs
junction
andwas nottransferredtotheright
Us-Rs
junction.
Effect ofthepresenceoftheduplicated
region
on transcrip-tion and growth rate of strain B3 in culture cells. Theduplicated
region
ofB3existed within the transcribedregion
of gene US1(IEmRNA-4)
and was outside thepredicted
protein-coding
regions (14, 17).
Toexamine the effectofthe presence of theduplicated
region
on thetranscription
of geneUS1,
immediate-early
mRNAs(IEmRNAs)
ofB3 and SP23wereprepared
andanalyzed by
Northernblotting (Fig.
6)
(12,
21, 30).TheIEmRNA-3(gene
IE175)
and IEmRNA-4 (geneUS1) produced
by
infection with B3werecomparable
tothose with SP23. The resultsindicated that the presence of theduplicated region
on gene US1 hadnosignificant
effect onthetranscription
of gene USL.Toexamine theeffect ofthe presenceofthe
duplication
on the growth ofHSV-1,
one-stepgrowth
curves of B3 and SP23 onVero cellswere constructed(2).
Thegrowth
curve of B3wascomparable
tothatofSP23; hence,
the presenceof theduplicated
region probably
had nosignificant
effect onthe
growth
of B3 in culture cells. DISCUSSIONHSV-1strain B3wasfoundtohavea
duplicated
region
onits left
Us-Rs
junction,
and the nucleotide sequenceon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.55.292.76.480.2]TaqI
(1) (L) TCGAGGCGACCGGCGGCGACCGTTGCGTGGACCGCTTCC T
50 IRs+ Us 11
GC TCG TCGGGGCGGGGG GA AGCC AC TG TG GTC C TC CGGG AC
10 1 1 12 30
TRs , Us
(R) GC TCGTCGGG:GGGGAGC ATGTCGTGGGCCC TGGAAATGGC
10 1(1) (8) (30)
12979 12972 12950
TaqI o.uR-seq
(2) (L) TCGACGCGACCGGCGGCGACCGTTGCGTGCQACCGC T TCCT
50 lRs5 3534R-seq U-seq 20 11
GC TC GTCGGGGGGGGGGGAGCGACCGTTGCG TGG TC CGCT
' _12g
10 1 - 11u-seq-.Us -25 -30
TC C TGCrC
GTCGGT|GGGGGGGAG~CC
AC TG TGGTCC TC CG-31 -44-45 -5512 26
TRs . Us
(R) GCTCGTCGGG[G13GGGGGAGCATGTCG TGGGC CCTGGAAAT
10 1-1 -10 12953
12972
FIG. 4. (1) Nucleotide sequences of the regions spanning left (L) and right (R)Us-Rsjunctions ofHSV-1strain17,published by McGeoch etal.(13, 14). The locations of U -Rsjunctions and the numbering system with thereferencepoint at the left U -Rsjunction were as described by McGeoch etal. (13, 14). Thenucleotidenumbers ofUsextending from right Us-Rsjunction are given in parentheses. Thetranslational initiationcodonATGof gene US12 is underlined. The Taql sitecorresponds toTal in Fig. 1 (3). (2) Nucleotide sequences of the regions spanningleft(L) andright(R) Us-Rsjunctions ofHSV-1 strain B3. The nucleotide numbers are the same as those used byMcGeoch et al. (13, 14), exceptforthe regions numbered with a minus, as described for Fig. 3. Two homologous sequences of R-seq (residue 33 onRsside toresidue -11) and U-seq(residue -12 toresidue -55) are indicated. The junction stretches are boxed. Theinitiation codon ATG of gene US12 is underlined.
ningtheduplicated regionwasdetermined [Fig. 4(2L)]. Two homologous sequences were present around thejunction. One isfrom residue33toresidue -11(R-seq),and the other isfrom residue -12 to residue -55 (U-seq). Both sequences were 44 bp in length and differed at two residues. One difference is between residue 20 on R-seq (A) and residue -25 onU (T). The otheris between residue 1 on R-seq(G) andresidue -44 onU-seq (T). Thebasesatresidues 20 and 1 on R-seq are the same as those at the corresponding residues in
TRs
of strain B3 andRs
of other HSV-1 strains. Theduplicationwasassumedtobegenerated byaninter-or intramolecular recombination between a point onRs
of a hypotheticalancestorofstrain B3 [I in Fig. 7 (1)] and theUs
side end of the junction stretch ofthe ancestor [I' in Fig. 7 (1)]. Ahomology of two bases of GC, atresidues 35 and 34 in
Rs
[I of Fig. 7 (1)] and at residues -10 and -11injunction stretch [I' of Fig. 7 (1)] was present on the recombined region. After formation of the duplication, the bases of A at residue -25 and G at residue -44 in the U-seq copy were assumedtohave been substitutedbyT[IIin Fig. 7(2)]. The proposed pathway for generation of the molecule with the duplication from a molecule without the duplication is shown in Fig. 7. However, there remains the possibility that strain B3 might be derived from an HSV-1 ,molecule having dupli-cationsonboth sides ofUs
iftheduplication on the right side is removed.The duplication in strain B3 was fixed on the left
Us-Rs
junctionandwas nottransferredtotheright side of
Us
(Fig. 2). I had already isolated two novel HSV-1 derivatives, SP22-4 and SP26-3, having duplications of DNA sequences containinggeneUS12 and theorigin ofDNAreplication (27). The duplicated sequences were amplified and transferred fromoneside to theother inUs
asaresultofrecombinations between the region deriving fromRs
on the duplicated sequencesand thecorresponding homologous region ofRs.
Two structural features of theduplicatedsequenceof strain B3 are perhaps involved in fixation of the duplication, in comparison with those of SP22-4 and SP26-3. One is the shortnessof theduplicatedsequenceof strain B3,i.e.,44bp. Recombinationbetween thetwosequenceswith suchashort homology would berare.The other is the absence of origin ofDNAreplicationintheduplicated sequenceof strain B3. The regions around the
origin
are assumed to be unstable and recombinogenic. A DNA fragment with the origin, replicating separatelyfrom viral genomes, may beamplifiedor transferred, as was interpreated for the VZV genome rearrangements by VlaznyandHyman (29).
The two copies of the inverted repeats enclosing one
unique sequencerecombine with each other in HSV-1 rep-lication (5, 9, 24, 28). Exchanges ofgeneticmaterialsby the recombinations function to maintain homogeneity between the two copies of the inverted repeats (13). Of the two homologous sequences presentonthe
Us-Rs
junctionof B3 (R-seq and U-seq), the nucleotide sequence of the R-seqon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.159.473.70.367.2]IRs+ j.s
(i4 (L)
GTCGGGGGGGGGAAGCCAC
I
6 1 1 12
TRS4H
Us
(R) GTC
GGG
GGAGCATG
6 11 8
IRS |US
(2) (L) GTCGGG
WGGGGGGGGGAGC CAC
6 1 1 14
TRS
4-US
(R) GTCGGG
IAAAAG:CATG
6 11 7
IR5
US
(3) (L)
GTCGGGGGGGGGAGACAC
65 11 4 5
TRs+ US
(R)
G TCGGG|GG
G AGC|A
T GII r -I I
11 6 5 1 1 5
IRS
.j
Us(4) (L) G
TCGGGGGGGGAGCC
AC6 11 9
TRs.j*
US
(R)
GTCGGGAAAAGCATG
6 1 1 7
IRs- Junction stretch *-U5
(5)
(L)
GTCGGG(GorA)
AGCk AC6 1-1 12
TRS
oJunction stretch *-U5
(R)
GTCGGG
(G
orA)
AGC ATGI I I
6 1-1 12972
FIG. 5. (1 to 4)Nucleotidesequencesof the regions spanning left (L) andright (R) Us-Rsjunctions of HSV-1 strains 17 (panel 1), Patton(panel2), B4 (panel3), and B9(panel 4). The sequences of strain 17 (panel 1) are from McGeoch et al. (13, 14), and the sequence aroundright Us-Rsjunction of strain Patton was from WatsonandVandeWoude (31). Other sequences were determined in the present work. The Us-Rsjunctions are placed at the point overwhich the homology between the two Rs sequences was no longer maintained. The nucleotidesare numberedextending inboth directions fromtheUs-Rsjunction. Junction stretches are boxed. (5) Consensus structurein andaroundthe Us-Rsjunction. The num-beringsystems arethe same as those used by McGeoch et al. (13, 14),exceptforthe region ofjunction stretch. The nucleotides within thejunction stretch are numbered with a minus symbol from the residue contiguousto R . Inthis definition- the A of initiationcodon ATG ofgeneUS12is located at the right end ofUs.
copywasthesame asthecorrespondingregion in
TR5
[Fig. 4(2L) and7(2)]. However, the U-seq copydjfferedfrom the corresponding region ofTRS
at tworesidues [Fig. 4 (2L) and 7 (2)]. These results suggest that there were no effective recombinational events to maintain homogeneity between U-seq copy andTRs.
Thus, the U-seq copy functionally resembledUs,
though the sequence was derived fromRs.
A 1 2
_
-B 1 2
__
-28S
-
18S
FIG. 6. Northern blotting analyses of IEmRNAs of HSV-1 strains SP23(lanes 1) and B3(lanes2). IEmRNAswereprepared by infecting Verocell monolayerswith HSV-1 in medium containing cycloheximide,asdescribed before (30). Polyadenylated RNAwas
selected on an oligo(dT)-cellulose column,denatured with glyoxal anddimethylsulfoxide, electrophoresed on an 1%agarosegel,and transferredtoa Biodyne A transfer membrane(12, 21).The mem-brane washybridized witha32P-labeled0.48-kbSacI-Xholfragment fromahybridphage Dec36carryingtheEcoRIHfragment ofHSV-1 (26) (A). The bands detected with the0.48-kb fragment represent mRNAs ofgene US1 (IEmRNA-4). The membrane was then hy-bridized with 32P-labeled 0.35-kb SmaI fragment of hybrid phage XNDE54 carryingEcoRlfragment ofHSV-1class Idefective DNA (22) (B). The bands detected with the 0.35-kb fragment represent mRNAsofgene IE175 (IEmRNA-3).Thepositions of28S and18S calfliver rRNAs(Pharmacia,catalogno.27-2506-01)areindicated.
The
Us-like
character of the U-seq copy is relevant to thefixation of theduplicated sequence onthe left side ofUs. There-is thehypothesis that
Rs
regionsofHSVandVZV wereformed by expansion ofRs
of an ancestral herpesvirus through a nonhomologous recombination between twoUs
sequences arranged in an opposite orientation (7, 32). This approach related the structures of the S components of HSV-1 and VZV by a discrete number of steps involving expansion orcontraction ofRs
(3). The discoveryof HSV variants with expanded repeat sequences and contracted uniquesequences,aspredictedby the hypothesis,supported the presence of a mechanism which mediates the process suggested by the hypothesis (1, 11, 23). In the hypothesis, theUS-Rs
junctions correspond to recombinedpoints. Un-der thedefinition ofUs-Rs
junctioninthiswork,theUs
side end of the junction stretch forms one side of theUs-Rs
junction, i.e., therecombinedpoint.TheR.
side ofthenew recombinedpoint in strain B3[residue -11inFig.4(2L)and 7 (2)], which forms the boundary between the R-seq copy andU-seq copy,wastheUs
side endofthejunction stretch. These results suggest that theUs
side end of thejunction stretch possesses the property of recombinogenicity in-volved in theformation ofUs-Rs
junctions of HSV-1 strains and also in generation ofa novel duplication of strain B3. Other HSV variants generated by such a mechanism may also exist. However, except for strain B3 described in this work, structuralalterations indicating therecombinogenicity
of theUs
side end ofthejunction stretch have not been reported. In this sense, probably, the recombinationattheUs
side end of thejunction stretchis not sofrequent.In
HSV-2,
the Aof the ATGasthetranslational initiation codonof the IEmRNA-5 gene (geneUS12)wasthefirst base inUs
and defined the rightUs-Rs
junction. Whitton and Clements assumed that the ATG acts asabufferto preventon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.374.498.76.228.2] [image:5.612.60.292.77.482.2]IRS5
Junction stretch * Us35 34 20 5 1 12
I -
---CGGCGA---A---TCGGIGGGGGGGGAGCICA---Recombination
I ----TC
GGG
GGGGGGGGAGCjC
A---5 1-1 -11 12
IR5s
Junction stretch * UsIRs - US
5 1 -11 -12 -25 -44 -55 12
( 2) E ----TCGG|GGGGGG GGAUC GA--
---TCGGGGGGGGGGAGC|CA---R-seq *
46-T
[image:6.612.100.521.75.329.2]U-seq *W
FIG. 7. Model for thegeneration of the duplicated region in HSV-1 strain B3. (1) Existence ofan ancestral HSV-1 strain withoutthe
duplicated regionwas assumed. Theoccurrence ofaninter- orintramolecularrecombination between the twoRs sequences on the2-bp
homology(residues 35 and 34 inmolecule I and residues -10 and -11 in molecule F') is shown. The numberingsystemisasdescribedfor
Fig. 5 (5). Junction stretchesareboxed. (2) Product of therecombinationalevent (molecule lI). The numberingsystemis the same asthat
forFig. 4(2L). ThetwohomologoussequencesofR-seq and U-seqareindicated.The base substitutionsatresidues-25 and-44onU-seq
wereassumedtohaveoccurredafter generation of the duplication. theexpansion of
TRs
(32). In theirdefinition,the AofATGastheinitiation codon ofgeneUS12 of HSV-1 strain 17was
at the eighth residue in
Us
from the rightUs-Rs
junction. They supposed thatthe Aof the ATG of HSV-1 geneUS12 would come to be positioned 1 bp from the rightUs-Rs
junction,likethatofHSV-2, if equal though nonhomologous
crossoverweretakeplaceand thefirst ATGwereto actas a
bufferto prevent the expansionof
TRs.
In thedefinition in thepresentwork,the Aof the ATGastheinitiator codonofgene US12 ofHSV-1 is the first base in
Us,
as is that of HSV-2 [Fig. 5 (5R)]. Presumably, theRs
of HSV-1 had alreadyexpandedtothepointat which the initiator ATG ofgeneUS12 actedas abuffertopreventtheexpansionof
TRs.
It isprobable that the presence ofgeneUS12may functionadvantageously forreplicationormaintenanceof HSV in the human body, although gene US12 is not required forvirus growth in cultured cells (1, 10, 23). With respect to the degree ofexpansion of
Rs,
it is assumed that HSV-1 and HSV-2 are located at the same step in the hypothetical descentfromanancestral herpesvirus (6).The conceptofjunctionstretchwas introduced todrawa
constant line ofdemarcation between
Us
andRs
(Fig. 5). Two aspects in nucleotide sequences around theUs-Rs
junction support the definition of
Us
side boundary of the junction stretch. Onewas the presence ofthree conservednucleotides, 5'-AGC-3', on the
Us
side boundary of thejunction stretch, and theotherwasthepresence of theATG initiationcodononright-side boundaryof
Us.
However,theRs
sideboundaryof thejunction stretch wasdefined solelyso as to seta common
Rs
sequence adjoining thejunctionstretch, by comparing the nucleotide sequences of four
HSV-1 strains. Thus, the
Rs
side boundary of thejunction stretch is not so certain as theUs
side boundary. Theconceptofjunctionstretchmadefeasible the definition of the regions spanningthe
Us-Rs
junctioninthe formcommontoanyHSV-1 strain [Fig. 5(5)]. For example,thediscrepancy
between strains 17 and Patton in the previously determined distancefrom the
Us-Rs
junction tothe translational initia-tion codonofgeneUS12could be eliminatedby makinguseof the concept ofjunction stretch [Fig. 5 (1Rand 2R)] (16, 31). Thelengthand basecompositionof thejunctionstretch
are variable among HSV-1 strains and also between two
Us-Rs
junctionsofanHSV-1strain(Fig. 5).Thepresenceofanadditional44-bpsequenceonthejunctionstretch of strain B3had no serious effects onviral replication. Thejunction stretchesseemtobetolerant of such variations. The loss of uniformitybetweenapairofjunctionstretches derivedfrom
one strain isregarded as a
Us-like
property of thejunctionstretch, althoughthejunction stretches wereassumedtobe derived from
Rs.
Thepolymorphisminnucleotidesequencesofjunction stretches may serve as physical markers for genetic analyses ofthe HSV-1 genome.
ACKNOWLEDGMENTS
I thank M. Ohara forhelpful comments.
A partofthis workwassupported bygrants from theMinistryof
Education, Science andCulture ofJapan. LITERATURE CITED
1. Brown, M., and J. Harland. 1987. Three mutants of herpes simplex virus type 2: one lacking the genes US10, US11 and
US1andtwoinwhichRshasbeenextendedby6kbto0.91map unit with loss ofUs sequences between0.94 and the Us/TRs junction.J. Gen. Virol. 68:1-18.
2. Dargan, D. J., and J. H. Subak-Sharpe. 1985. The effect of
triterpenoid compoundson uninfected and herpes simplex
vi-(1 )
on November 10, 2019 by guest
http://jvi.asm.org/
rus-infected cells in culture. 1. Effect on cell growth, virus particles and virus replication. J. Gen. Virol. 66:1771-1784. 3. Davison, A. J., and D. J. McGeoch. 1986. Evolutionary
compar-isons of the Ssegments in the genomes of herpes simplex virus type 1 andvaricella-zoster virus. J. Gen. Virol. 67:597-611. 4. Davison, A. J., and J. E. Scott. 1986. The complete DNA
sequenceof varicella-zoster virus. J. Gen. Virol. 67:1759-1816. 5. Davison, A. J., and N. M. Wilkie. 1983. Inversion of the two segments of the herpes simplex virus genome in intertypic recombinants. J. Gen. Virol. 64:1-18.
6. Gentry, G. A., M. Lowe, G. Alford, and R. Nevins. 1988. Sequence analyses of herpesviral enzymes suggest an ancient origin for human sexual behavior. Proc. Natl. Acad. Sci. USA 85:2658-2661.
7. Honess, R. W. 1984. Herpessimplex and ''the herpescomplex": diverse observations and aunifying hypothesis. J.Gen. Virol. 65:2077-2107.
8. Honess, R. W., and D. H. Watson. 1977. Unityanddiversity in the herpesviruses. J.Gen. Virol. 37:15-37.
9. Knipe, D. M., W. T. Ruyechan, B. Roizman, and I. W. Halli-burton. 1978. Moleculargenetics of herpes simplex virus: dem-onstration of regions of obligatory and nonobligatory identity within diploid regions of the genome by sequence replacement and insertion. Proc. NatI. Acad. Sci. USA 75:3896-3900. 10. Longnecker, R., and B. Roizman. 1986.Generation of an
invert-ing herpes simplex virus 1 mutant lackingthe L-S junction a
sequences, an origin of DNA synthesis, and several genes includingthose specifying glycoprotein E and the a47 gene. J. Virol. 58:583-591.
11. Maclean, A. R., and S. M. Brown. 1987. Deletionand duplica-tion variants around the long repeats ofherpes simplex virus type 1 strain 17. J. Gen. Virol. 68:3019-3031.
12. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual, p. 200-201. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
13. McGeoch, D. J., A. Dolan, S. Donald, and D. H. K. Brauer. 1986. CompleteDNAsequenceof the short repeat region in the genome of herpes simplex virus type 1. Nucleic Acids Res. 14:1727-1745.
14. McGeoch, D. J., A. Dolan, S. Donald, and F. J. Rixon. 1985. Sequencedetermination andgeneticcontentofthe short unique region in the genome of herpes simplex virus type 1. J. Mol. Biol. 181:1-13.
15. Messing, J. 1983. New M13 vectors for cloning. Methods Enzymol. 101:20-78.
16. Murchie, M.-J., and D. J. McGeoch. 1982. DNA sequence analysisofanimmediate-earlygeneregionof theherpes simplex virus type 1 genome (map coordinates 0.950 to 0.978). J. Gen. Virol.62:1-15.
17. Rixon, F. J., and D. J. McGeoch. 1985. Detailed analysis of the mRNAs mappingin the short unique region of herpes simplex virus type 1. Nucleic Acids Res. 13:953-973.
18. Roizman, B. 1979. The structure and isomerization of herpes
simplex virus genomes. Cell 16:481-494.
19. Roizman, B., L. E. Carmichael, F. Deinhart, G. de-The, A. J. Nahmias, W. Plowright, F. Rapp, P. Sheldrick, M. Takahashi, and K. Wolf. 1981. Herpesviridae: definition, provisional
no-menclature, and taxonomy. lntervirology 16:201-217.
20. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
21. Umemura, T., K. Umene, J. Nishimura, Y. Fukumaki, Y.
Sakaki, and H. Ibayashi. 1986. Expression of c-myc oncogene during differentiation of human burst-forming unit, erythroid (BFU-E). Biochem. Biophys. Res. Commun. 135:521-526. 22. Umene, K. 1985. Variabilityof theregionof theherpessimplex
virus type 1 genome yielding defective DNA: Sinml fragment polymorphism. lntervirology23:131-139.
23. Umene, K. 1986. Conversion ofa fraction ofthe unique
se-quence to partofthe inverted repeats in the S component of the herpes simplex virus type 1 genome. J. Gen. Virol. 67:1035-1048.
24. Umene, K. 1987.Transition from a heterozygous to a homozy-gous stateof apairof loci in the inverted repeat sequences of the L component of the herpes simplex virus type 1 genome. J. Virol. 61:1187-1192.
25. Umene, K. 1987. Restriction endonucleases recognizing DNA sequencesof four base pairsfacilitate differentiationof herpes simplex virus type 1 strains.Arch. Virol. 97:197-214.
26. Umene, K., and L. W. Enquist. 1981. A deletion analysis of lambda hybrid phagecarrying the Usregion of herpessimplex virus type 1 (Patton). I. Isolation of deletion derivatives and identification ofc/i-like sequences. Gene 13:251-268.
27. Umene, K., andL. W.Enquist. 1985. Isolation of novelherpes simplex virus type 1 derivatives with tandem duplications of DNA sequences encoding immediate-early mRNA-5 and an
originofreplication.J. Virol. 53:607-615.
28. Varmuza,S.L.,andJ. R. Smiley. 1984. Unstableheterozygosity in a diploid region of herpes simplex virus DNA. J. Virol. 49:356-362.
29. Vlazny, D.A., and R.W.Hyman. 1985. Errantprocessingand structural alterations of genomes present in a varicella-zoster virus vaccine. J. Virol. 56:92-101.
30. Watson, R. J., M. Sullivan, and G. F. Vande Woude. 1981. Structures oftwo spliced herpes simplex virus type 1 immedi-ate-early mRNAs which map at thejunctions oftheunique and reiterated regions of the virus DNA S component. J. Virol. 37:431-444.
31. Watson,R.J.,andG. F.Vande Woude. 1982. DNAsequenceof
animmediate-early gene (IEmRNA-5) of herpes simplex virus type 1. Nucleic Acids Res. 10:979-991.
32. Whitton, J. L.,andJ. B. Clements. 1984. Thejunctionsbetween the repetitive and the short unique sequences of the herpes simplexvirusgenomearedeterminedby thepolypeptide-coding regions oftwospliced immediate-earlymRNAs. J.Gen. Virol. 65:451-466.