JOURNALOFVIROLOGY,Mar. 1992, p. 1535-1542 0022-538X/92/031535-08$02.00/0
CopyrightC1992, American Society for Microbiology
Cleavage of the
Dengue
Virus Polyprotein
at
the
NS3/NS4A
and
NS4B/NS5
Junctions
Is
Mediated
by Viral
Protease
NS2B-NS3,
Whereas
NS4A/NS4B
May Be Processed by
a
Cellular
Protease
ANNIECAHOUR, BARRY FALGOUT, AND CHING-JUH LAI*
Molecular ViralBiology Section, Laboratory of Infectious Diseases, National Institute of Allergy and
Infectious
Diseases,
Bethesda,
Maryland
20892Received13 September 1991/Accepted3December 1991
The cleavage mechanism utilized for processing of the NS3-NS4A-NS4B-NS5 domain of the dengue virus
polyprotein was studied by using the vaccinia virus expression system. Recombinant vaccinia viruses vNS2B-NS3-NS4A-NS4B-NS5, vNS3-NS4A-NS4B-NS5,vNS4A-NS4B-NS5, and vNS4B-NS5wereconstructed.
These recombinantswereusedtoinfectcells,and the labeledlysateswereanalyzed by immunoprecipitation.
Recombinant vNS2B-NS3-NS4A-NS4B-NS5 expressed the authentic NS3 and NS5 proteins, but the other recombinants produceduncleaved polyproteins. These findings indicate that NS2B is required for processing ofthe downstream nonstructural proteins, includingtheNS3/NS4Aand NS4B/NS5junctions, both of which contain a dibasic amino acid sequence preceding the cleavage site. The flavivirus NS4A/NS4B cleavagesite follows a long hydrophobic sequence. The polyprotein NS4A-NS4B-NS5 was cleaved at the NS4A/NS4B junctionintheabsence of other dengue virus functions. One interpretation for this finding is that NS4A/NS4B cleavage is mediated by a host protease, presumably a signal peptidase. Although vNS3-NS4A-NS4B-NS5
expressed onlythepolyprotein, earlier results demonstrated that cleavageatthe NS4A/NS4Bjunction occurred whenananalogousrecombinant, vNS3-NS4A-84%NS4B,wasexpressed. Thus, itappearsthat uncleaved NS3
plus NS5 inhibitNS4A/NS4B cleavage presumably because the putative signalsequence isnotaccessiblefor recognition by the responsible protease. Finally, recombinants that expressed an uncleaved NS4B-NS5
polyprotein, such as vNS4A-NS4B-NS5 or vNS4B-NS5, produced NS5 when complemented with
vNS2B-30%NS3orwithvNS2Bplus v30%NS3. These resultsindicatethat cleavageatthe NS4B/NS5junctioncanbe
mediatedby NS2B and NS3intrans.
The four serotypes ofdengue virus are members of the
Flaviviridae,afamily ofsome 70 viruses,mostofwhichare
transmittedby mosquitosorticks(45). Manyflavivirusesare
human pathogens and cause a variety of diseases such as
yellow fever, dengue, Japanese encephalitis, or tick-borne
encephalitis. Among flaviviruses, dengue viruses have the highest incidence of infection and the widest geographic distribution (16). Forthis reason, intensive efforts are
cur-rently beingdirectedatresearchondenguevirus. Like other
members of the flavivirus family, dengue virus contains a
positive-strandRNAgenomeapproximately 10 kb inlength (27). Complete nucleotide sequences ofthree dengue virus serotypes and several other major flaviviruses have been determined (11, 12, 17, 19, 24, 25, 29, 30, 34, 43, 47). The results of amino acid sequence data or alignment with the established protein sequences of other flaviviruses indicate that the dengue virus RNAgenome codes fora long
poly-proteinwith the order ofNH2-anchored C (anchC-pre-M-E-NS1-NS2A-NS2B-NS3-NS4A-NS4B-NS5-COOH (2-5, 8, 11, 28, 33, 39, 40, 43, 46). This body of sequence information indicates that flaviviruses share the same
ge-nome organizationand presumablythe same mechanism of
gene expression and viralreplication (7, 42). The flavivirus
polyproteinisapparentlycleavedco-andposttranslationally
togenerate the individual viral proteins.
Analysisof amino acid sequences nearthecleavage sites of flavivirus polyproteins suggests that there are several
cleavage mechanisms ofprotein processing. The cleavage
*Correspondingauthor.
site at each of the anchC/pre-M, pre-M/E, EINS1, and NS4A/NS4B junctions follows a long hydrophobic region,
andcleavageat these sites is believedto be mediated by a
host cell signal peptidase. Evidence supporting signal-di-rected cleavage thatgenerates the three structural proteins has been obtained from in vitro translation and processing studies (26, 28, 37). Cleavage at the NS1/NS2A junction, presumably mediated by a novel protease, requires an
eight-amino-acid sequence at the NS1 C terminus and the downstreamNS2A(13, 18).Thecleavage sitesatthe NS2A/ NS2B, NS2B/NS3, NS3/NS4A, and NS4BINS5 junctions share a common sequence motif in which apair of basic amino acids(RR, KR, RK),orQRattheNS2B/NS3 junction indengue viruses, precedeseitherG, S,orA(34, 39).This class ofprocessing events is thought to be mediated by a
virus-coded protease inthe cytoplasm (21, 35). It has been proposed on the basis of limited sequence homology to serine proteases that NS3 is a viral protease (la, 15). The N-terminal third of flavivirus NS3 contains three appropri-ately spacedconserved amino acids(Hisatposition 51, Asp
at94, and Serat 135 indengue type4 virus [DEN4] NS3),
proposedto be the catalytic triad. In addition, amino acid
sequences predicted for the substrate-binding pocket are
also present. Experimental evidence supporting this pro-posal has been obtained by using in vitro translation and
processingofpolyproteinprecursors (10, 31, 44).Our labo-ratorystudied the viral functions thatarerequiredfor these processing events in vivo, using recombinant vaccinia
vi-ruses expressingvariousportionsof the DEN4polyprotein. We observed that DEN4 polyproteins that contain a large
deletionwithin NS2Bwerenotcleaveddespitethepresence
1535
Vol. 66, No. 3
on November 10, 2019 by guest
http://jvi.asm.org/
1536 CAHOUR ET AL.
-4 NS5(900 aa)->
I I I I
SmaI Pst I Hind III
- -
-ATCccc AS4| CG GCCC CTGCGAGA GGCAAAGC TTi CCCTAC|CCCT CGG SACGTIC TCT CCGTTTCSGAT
pATH2/NS5-1 pATH3/NS5-2
2SrpBAIS5-1(aa1-199) TxpZ/NS5-2(aa 199-757)
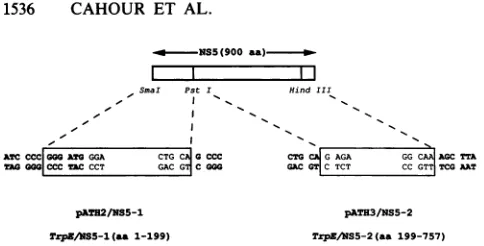
FIG. 1. Sequences of DEN4 NS5 cDNA in TrpE expression
vectors. Sequences at thejunctions between the bacterial TrpE vector, pATH2 or pATH3, and the NS5 cDNA fragment to be
expressedare shown. The two plasmid vectorsdiffer only in the
readingframein thepolylinker region.NS5sequencesbetween the
SmaI and PstI sites were cloned in the corresponding sites of pATH2tocreatepATH2/NS5-1;NS5 sequencesbetween thePstI
andHindIII siteswere cloned inpATH3to create pATH3/NS5-2. Polylinkersequences and non-dengue virus sequencesare inbold
letters. Thelengths of aminoacids (aa)encodedin the expressed
NS5segmentsareindicated.
of the entire NS3 sequence. These results showed that
NS2B, in addition to
NS3,
is required for cleavage at theNS2A/NS2B and NS2B/NS3junctions and presumably for
cleavageattheNS3/NS4Ajunction (14).Wefurther showed that NS2B is capable of acting in trans. Only indirect evidenceindicatingthatNS3 canact in transwasobtained. The current study was initiated to examine the cleavage mechanisms utilized for processing of the remaining
non-structural proteins, i.e., NS4A, NS4B, and
NS5,
from theNS3-NS4A-NS4B-NS5regionof the DEN4polyprotein.We presentdatademonstratingthatcleavageattheNS4A/NS4B junction is apparently mediated by a signal peptidase-like
hostproteaseand thatNS4B/NS5 cleavagecanbemediated
byboth NS2B andNS3 in trans.
MATERIALS ANDMETHODS
Expressionof
TrpE/NS5
fusionproteinsandpreparationof antisera. ThebacterialTrpE expressionvectorspATH2and pATH3 (41)were kindlyprovided byG. Ketner(TheJohns Hopkins University, Baltimore, Md.). Initially, weprepared a full-length NS5 cDNA fragment by polymerase chain reaction(PCR), using appropriate primers thatintroducedaSmaI site preceding the ATG codon for initiation ofNS5 synthesisandaKpnIsiteatthe 3' terminus ofdengue virus cDNA. Full-length dengue virus cDNA clone 2A was used
asthetemplate (23).Twosubcloneswereseparatelyinserted
into the appropriate vector forexpression ofNS5 (Fig. 1). Plasmid
pATH2/NS5-1
contained the SmaI-PstI fragment (nucleotides[nt]7560to8152),andpATH3/NS5-2contained the PstI-HindIII fragment (nt 8152 to 9829). Plasmidcon-structswereusedtotransformEscherichiacoli C600. Trans-formantsweregrowninM9CAmediumcontaining ampicil-lin and induced with ,-indoleacrylic acid (10 ,ug/ml) (20). Bacteriawere pelleted and resuspended inTEN buffer (50 mM Tris HC1 [pH 7.5], 0.5 mM EDTA, 0.3 M NaCl) containing 2 mg of lysozyme per ml, disrupted by freeze-thawing, and treated with 3 mg of DNase I per ml. The
insoluble fraction was semipurified by three cycles of
cen-trifugation and washing with TEN buffer containing 0.1% NonidetP-40. Thepelletwasdissolvedby heatingat37°C for 30 minin Laemmli buffer (0.01 MTris HC1 [pH 6.8], 20% glycerol,5mMEDTA,4%sodiumdodecylsulfate[SDS], 50 mM ,-mercaptoethanol,0.002%bromophenol blue)
contain-ing 20 ,ug of phenylmethylsulfonyl fluoride per ml
(22).
Samples were separated on SDS-polyacrylamidegels
and stained with Coomassie blue. Fusionprotein
bands wereexcised, washed with distilled water,
homogenized
in phos-phate-bufferedsaline(PBS)containing0.1%SDS,
and emul-sified with an equalvolume ofcomplete
Freund'sadjuvant.
This mixture was used to inoculate New Zealand White rabbitsintradermallyalongthebackat100to300
pug
of each fusion protein per dose. Booster inoculationsusing
the fusionprotein-polyacrylamide gel homogenate,emulsified in incomplete Freund's adjuvant, were performed at 4 and 8weeks following theprimary immunization. Serum
samples
were collected beginning2 weeks afterthe lastinoculation.
Construction of recombinant vaccinia viruses. DEN4 cDNAfragmentscodingfor various
polyproteins
within theNS3-NS4A-NS4B-NS5 domain were inserted into the
vac-cinia virus intermediate transfer vector pSCll(BglII)
(13).
TheN-terminal amino acidpositions of
NS2B,
NS3,
NS4A,
NS4B, and NS5 have been determined by alignment of the deduced DEN4 polyprotein sequencewith thatofflavivirusKunjin (14). To construct pSCll/NS4A-NS4B-NS5 and pSC11/NS4B-NS5, initiallyaDEN4 cDNA
fragment
coding
for NS4A-NS4B was prepared by polymerase chainreac-tion, using full-length DEN4 2A cDNA as the
template,
oligodeoxyribonucleotide (oligo) 2850
(5'-GCCGGATCCA
CCATGAGTATAACTCTCGAC-3') tointroducethe initia-tion codonprecedingthe firstaminoacid of NS4A, and
oligo
2851 (5'-TCCTGGATCCTACCTCCTAGGGGTTTGTGC-3')toprovidea stopcodon. Similarly,DEN4cDNA
coding
for NS4Bwaspreparedby usingthe 2AcDNAtemplate and oligo 2445 (5'-AAGATCTATGTTGATCTACGTCATATTG AC-3'), which containsaninitiation codon preceding thelast
17amino acidsof the putative 19-amino-acid signal ofNS4B, andoligo 2851as primers. ThePCRproducts wereinserted separately into pSCll(BglII). The DEN4 insert in these pSC11 derivatives wasextended to include the entire NS5 coding sequence by fragmentexchange with
pSC11/NS2B-NS3-NS4A-NS4B-NS5, usingthe BsmI site at nt 7544 and the XhoI site in pSC11 DNA. DEN4 cDNA inserts in the
constructsofpSC11/NS4B-NS5 expressingshortenedforms of NS5 such as 50%NS5 (terminating at nt 8912) and 22%NS5(terminatingat nt8141) were obtainedby
polymer-ase chain reaction, using plasmid pSC11/NS4B-NS5 as the
template
and appropriate primers. Plasmid pSC11/NS2B-NS3-NS4A-NS4B-NS5 or pSC11/NS3-NS4A-NS4B-NS5wasobtained by fragmentexchange between
pSC11/NS2A'-NS2B-NS3-NS4A-NS4B-NS5 and pSC11/NS2B-NS3 or
pSC11/NS3-84%NS4B
(14) at the unique BstBl site at nu-cleotide 5069 and the XhoI site in pSC11. PlasmidpSC11/NS2A'-NS2B-NS3-NS4A-NS4B-NS5
was constructed ear-lierby insertion of theDEN4 sequencebetween theStuI siteat nt 3616 and the 3' flanking PstI site (following the C/G sequence), blunted by T4 polymerase, at the SmaI site of
pSC11
(46a). The yellow fever virusNS2B-32%NS3 cDNA fragment (nt 4181 to 5180 of the yellow fever virus se-quence), encoding a 333-amino-acid polyprotein, was pre-pared by polymerase chainreaction, using appropriateprim-ersandplasmid pES/6,aderivative of clone pGX4EI (36),as
the template. All constructs described were verified by restrictionenzyme mapping and by sequencing of 50 to150
nt across the insertion sites. Both CV-1 and TK-141 cells
were propagated in Eagle's minimum essential medium
supplemented
with 10% fetalcalf serum. Recombinant vac-cinia viruses were constructed by using the wild-type vac-ciniavirusstrainWRandthe pSC11/DEN4DNA derivatives as described earlier (6, 13). Recombinant vaccinia virusJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.56.298.57.180.2]PROCESSING OF DENGUE VIRUS NONSTRUCTURAL PROTEINS 1537
vSC8, containingthe lacZgene but no dengueviruscDNA, wasusedas anegative control (6). Recombinant virus stocks were preparedfrom infected CV-1 cells, and the virus titer was determined by plaque assay on monolayers of CV-1
cells. The nomenclature for the recombinant virus con-structsandtheirencodedpolyproteins followed the previous
practice (14).
Radiolabeling and immunoprecipitation. CV-1 cells were
infected
with recombinant vaccinia viruses or the controlvaccinia virus vSC8 at a multiplicity of infection of 5 PFU per cell diluted in Eagle's minimum essential medium
con-taining 2% fetal calf serum (MEM2). At 16 to 24 h postin-fection, the medium was replaced with methionine-free MEM2 for 1 h and then with methionine-free medium
containing
L-[5S]methionine
(100 ,uCi/ml; specific activity,-800Ci/mmol) for2h. Mediumwas removedafter
labeling,
and thecells were rinsed twice with cold PBS and lysed in RIPA buffer (1% deoxycholate, 1% Nonidet P-40, 0.1% SDS, 0.1 MTris-HCl [pH7.5], 0.15 M NaCl)containing 20 ,ug of phenylmethylsulfonyl fluoride per ml. The labeled
lysate of DEN4-infected cells was kindly provided by M.
Bray ofourlaboratory.
Immunoprecipitation
was performed by using NS5-spe-cific antisera raised againstTrpEINS5
fusion proteins orDEN4hyperimmune mouseascitic fluid(HMAF). Briefly, a
50- to
100-pd
aliquot ofalabeled lysatewas mixedwith theNS5-specific antiserumor HMAF (3 to 5
IlI)
and incubated onice for 2 h. Thenanexcessof Pansorbinwasadded to themixture, and after incubation on ice for another 1 h, the
immune precipitates were collected by centrifugation and
washedtwice withRIPAbuffer containing2% SDS.
SDS-polyacrylamide gel electrophoresis. Immune
precipi-tates wereresuspended in Laemmli buffer and boiled for5
min before loading. Sampleswere analyzed by electropho-resison anSDS-8% polyacrylamide gel
(acrylamide/bisacry-lamide ratio of60:1.6). Gels were treated for fluorography and usedtoexpose X-rayfilm.
RESULTS
TrpE/NS5
fusion proteins and NS5-specific sera. In ourearlier
studies,
DEN4 HMAFwas used forprecipitation
of labeled lysates of infected cells, and we observed thatprecipitation
ofdengue
virusNS5wasbarely
detectable.Tofacilitate detection of NS5 orits
polyprotein
precursors, weprepared
NS5-specific
sera. Since NS5(103
kDa)
isarela-tively
largeprotein,
twosubfragments
ofNS5 cDNA wereseparately
expressed
in the form ofTrpE
fusionproteins by
usingabacterial TrpEvector. Asdetailed in
Materials
and Methods and shownschematically
inFig. 1,
fusionprotein
TrpE/NS5-1
contained amino acids 1 to 199 and fusionprotein
TrpE/NS5-2
contained amino acids 199 to 757 ofDEN4 NS5.
Analysis
of the bacteriallysate
showed thatboth
TrpE/NS5-1
andTrpE/NS5-2
of thepredicted
sizes(52
and 97
kDa,
respectively)
were detected in the insoluble fraction(Fig.
2A). These fusionproteins
were notdetected in the soluble fraction(data
notshown).
Rabbit antisera raisedagainst
each of these fusionproteins
weretestedfor theirability
to bind radiolabeled NS5 indengue
virus-infected celllysate.
Figure
2B shows that both antiseraspecifically
precipitated
a 103-kDa labeledband,
aspre-dicted for the NS5
protein.
No otherprotein
bands were seen in theprecipitates.
These results indicated that both antisera werespecific
fordengue
virus NS5. Since nodifferencein
NS5-binding
efficiency
wasevident betweentheA M 1 2 3 4
97
-B (5-I) (5-2)
H Pl PI M _ -trpE/5 -2
68- _ -
-I _ -trpE/5-1
43-
-29-@
N S5
-NS3-_
E-5. NS
I1-_~ 4 -9 2
-69
16-46
;14k-.mmw..
,.Imo- m
40M
qw
-3O
I8
preM- * - 18
FIG. 2. Characterization of TrpE/NS5 fusion proteins and NS5-specific sera. (A) TrpE/NS5 fusion proteins. The insoluble fraction of theE.coli C600 lysate was analyzed by SDS-polyacrylamide gel electrophoresis, and theprotein bands were stained by Coomassie blue. Lanes: M,protein size markers, shown in kilodaltons on the left; 1, control bacteria; 2, bacteria transformed by pATH2; 3, bacteria transformed bypATH2/NS5-1; 4, bacteria transformed by pATH3/NS5-2; (B) NS5-specific sera. Immunoprecipitation of an
L-[35S]methionine-labeled
lysateofDEN4-infected cells wascarried out totestthe NS5bindingspecificity of the rabbit antiserum raised against the pATH2/NS5-1 or the pATH3/NS5-2 fusion protein. Lanes: H, dengue virus HMAF; PI, preimmune serum; I, postim-munization serum; M,'4C-labeled
protein size markers, shown in kilodaltons on the right. DEN4 proteins are indicated on the left.twoantisera, only the antiserum raised against
TrpE/NS5-1
was used in thisstudy.
NS2B is required for proteolytic processing of dengue NS3-NS4A-NS4B-NS5. Because ofourinterest in
identifying
therequirements for dengue
polyprotein
processing
invivo,
we constructed a series of recombinant vaccinia viruses
expressing
variouslengths ofthe DEN4 NS2B-NS3-NS4A-NS4B-NS5sequence. DEN4polyproteins
encodedbytheserecombinant virusesarediagrammed in
Fig.
3A. TheN-ter-minal sequence of
polyprotein
NS2B-NS3-NS4A-NS4B-NS5 or NS3-NS4A-NS4B-NS5 was the same as that of NS2BorNS3constructed earlier(14);Met-Gly
preceded
the first amino acid(Ser)
ofNS2B, and Metpreceded
the Nterminus of NS3. Similarly,
polyprotein
NS4A-NS4B-NS5 containedaMetresiduepreceding
thepredicted
N-terminalsequenceof NS4A.TheNS4B-NS5
polyprotein
specified
by
vNS4B-NS5 contained the initiating Met
plus
the last 17amino acids ofNS4A
preceding
the Nterminusof NS4B.Allfour
polyproteins
containedthefull-length
NS5 sequenceatthe C terminus. These recombinants were used to infect cells, and the labeled
lysates
wereanalyzed
by
immunopre-cipitation using
HMAF or theNS5-specific
antiserum. Asshown in
Fig.
3B(lane 2),
vNS2B-NS3-NS4A-NS4B-NS5
expressed a
protein
with an apparent molecular size of73kDa,as
predicted
fortheNS3protein. Previously,
we have shown thatananalogous
recombinant,
vNS2B-NS3-NS4A-84%NS4B,
expressesauthenticNS3,
as identifiedby
comi-grationwith theNS3
product
ofDEN4 virus(14).
Inlane2,
twoprotein
bands,
migrating
atapproximately
52 and 54 kDa,appeared
to beDEN4specific
sincethey
wereimmu-noprecipitated by
HMAF. These bands werepreviously
observed
(14)
andprobably
derivedfrominternal
cleavages
of NS3. Severalother
large
polyproteins ranging
from 85to VOL. 66,1992on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.326.563.80.251.2]J. VIROL. 1538 CAHOUR ET AL.
N 2 NNj533 N54A NS4B
lI
\5577
__I_-ra1-
:
FU
'IB 1 2 3 4 5 6 7
210-W.
to i^; _ ..:.
210- 1' -NS3-NS4A-NS4B-NS5
_ -NS4A-NS4B-NS5
- --NS4B-N55
- NS5
694- ..1
46 * ,
46
-+ + + HMAF
+ + + + antiNS5
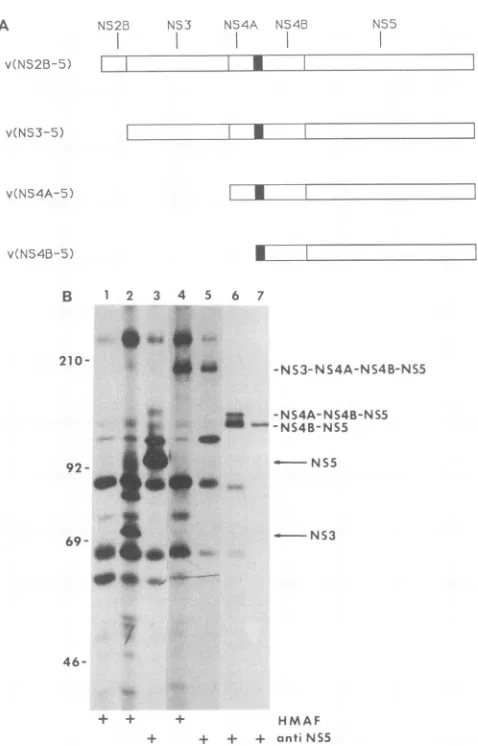
FIG. 3. Processingof various lengths of the NS2B-NS3-NS4A-NS4B-NS5domain ofthedengueviruspolyprotein.(A)Diagramsof various lengthsof dengue virus polyproteins expressed by
recom-binantvaccinia viruses. Theputative signalsequencethatprecedes theNS4A/NS4B cleavage junction is marked withafilled box. (B)
Analysis of dengue virus proteins expressed by recombinant
vac-cinia viruses. Radiolabeled lysates of CV-1 cells infected with recombinantvacciniaviruses shown in panel A were
immunopre-cipitated with polyvalent HMAF or the NS5-specific serum. The
labeledprecipitates wereanalyzed bySDS-8% polyacrylamidegel
electrophoresis. Recombinant vaccinia viruses used forinfection:
lane 1, vSC8control; lanes2and 3, vNS2B-NS3-NS4A-NS4B-NS5;
lanes 4 and5,vNS3-NS4A-NS4B-NS5; lane 6,vNS4A-NS4B-NS5;
lane7,vNS4B-NS5. Positionsof molecularsize markersareshown
in kilodaltonsontheleft.
210 kDa, whichwerenotseenin the control lysate,werealso
precipitated.Thesepolyproteins mostprobably represented
processingintermediatesorincompleteprocessingproducts,
for example, NS3-NS4A (86 kDa), NS2B-NS3-NS4A (100 kDa), and NS4A-NS4B-NS5 (146 kDa). Because antisera
specific for NS2B, NS4A, or NS4B were not available,
definite identificationofthese protein bands was not
possi-ble. When theanti-NS5 serum was used, alabeled band of
approximately 103 kDawas detected, aspredicted forNS5.
This finding indicates that cleavage at bothNS3/NS4A and
NS4BINS5 junctions had occurred. In contrast,
vNS3-NS4A-NS4B-NS5 did not express NS3 or NS5; rather, it
expresseduncleavedNS3-NS4A-NS4B-NS5. This result
in-dicates that NS2B is required for processing of the
down-stream nonstructural proteins, including the
NS3/NS4A
and NS4B/NS5junctions, bothofwhichcontain adibasic aminoacid sequence preceding the cleavage site.
Cleavage at the NS4A/NS4Bjunction. By
alignment
withthe established N-terminal sequence ofKunjin virus NS4B
(39), the N-terminal amino acid of DEN4NS4B is placedat Asn-2242. In the flavivirus sequence, the cleavage site between NS4A and NS4B is preceded by a runof
hydropho-bic amino acids that could serve as a signal for
cotransla-tional cleavage mediated by asignal peptidase. Inthecaseof
DEN4, this putative signal sequence includes 19 hydropho-bic amino acids. Since antisera specific forDEN4 NS4Aor NS4B were not available, we used the NS5-specific sera to determine whether the NS4B-NS5 cleavage product was made from the polyprotein NS4A-NS4B-NS5 precursor. Cells were infected with vNS4A-NS4B-NS5, and the labeled lysate was analyzed. Figure 3B (lane 6) shows that the NS5-specific antiserum precipitated two bands, both of which were larger than NS5; one was identified as uncleaved NS4A-NS4B-NS5 polyprotein on the basis of molecular size, and the other was identified as NS4B-NS5. This finding indicates that cleavage at the
NS4A/NS4B
junction had apparently occurred in the absence of the previously identi-fied NS2B-NS3 viral protease. This cleavage is probably mediated by a signal peptidase and may occur cotranslation-ally. This notion is further supported by a pulse-chase study which indicates that the relative amounts of NS4A-NS4B-NS5 and NS4B-NS4A-NS4B-NS5 did not change during the chase period (data not shown). The observation that NS4B-NS5 (lane 6) comigrated with the product of vNS4B-NS5 suggests that the signal sequence was cleaved from the vNS4B-NS5 product or that the presence of the signal did not affect the mobility. It should be noted that cleavage atNS4A/NS4B
did not occur when polyprotein NS3-NS4A-NS4B-NS5 was expressed, since NS4B-NS5 could not be seen. However, in a previous study we observed that cleavage at theNS4A/
NS4B junction did occur when vNS3-NS4A-84%NS4B was expressed (14). We speculate that uncleaved NS3 plus NS5 prevented NS4A/NS4B cleavage from taking place, perhaps because the putative signal sequence in this polyprotein is not accessible for recognition by the responsible protease.
Cleavage at the NS4B/NS5 junction by NS2B and NS3 in trans. Recombinants that expressed an uncleaved NS4B-NS5 polyprotein such as vNS4A-NS4B-NS4B-NS5 or vNS4B-NS4B-NS5 were used for coinfection of cells with vNS2B alone, v30%NS3 alone, or vNS2B plus
v30%NS3.
Lysates of infected cells were analyzed by immunoprecipitation with the NS5-specific serum. The results (Fig. 4) show that a labeled band of the predicted size for NS5 was detected during coinfection with vNS2B plusv30%NS3
but not with vNS2B orv30%NS3 alone. Note that the shift in mobility of NS4B-NS5 in lane 11 was probably an artifact since it was not observed in other experiments. As estimated by the intensity of the labeled protein bands, more than 50% of each polyprotein precursor was converted to the NS5 product. Interestingly, the NS4A-NS4B-NS5 polyprotein (lane 7) appeared to be completely processed, whereas a significant portion of NS4B-NS5 persisted. This finding suggests that NS4A-NS5 is more efficiently processed than NS4B-NS5 at the NS4B/NS5junction. These results indicate that cleavage at the NS4B/NS5 junction in the polyprotein was mediated by both NS2B and NS3 in trans. Additional evidence was sought to confirm that the appearance of an NS5 band in Fig. 4 resulted from proper processing at the NS4B/NS5 junction rather than from a spurious cleavageA
v(NS2B-5)
v(NS3-5)
v(NS4A-5)
vf(NS4B-5)
92
-.7
* jib. .":."
46
-,- 4mIt
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.54.293.72.445.2]PROCESSING OF DENGUE VIRUS NONSTRUCTURAL PROTEINS
vNS4A-NS4B-NS5 vNS4B-NS5
2B 2B coin
fecting
_ 2B 3 3 - 2B 3 3 virus
210-_
92
-_0 j&j4S -N54A-NS48-N55
m
~_
_i _ _-NS4B-NS5 NS4B-50%NS5 -69-N54B-2 2%N55- _ 4 6 -_W
[image:5.612.64.304.79.234.2]1 2 3 4 5 6 7 8 9 10 11
FIG. 4. Demonstration that cleavage oftheNS4B/NS5junction requires both NS2B and NS3. CV-1 cells were infected with
vNS4A-NS4B-NS5orwithvNS4B-NS5,producingthe DEN4
poly-proteintobe tested for cleavage.Lanes 4 and 8, control showing the uncleaved polyproteins; lanes 5 and 9, coinfection with vNS2B alone; lanes 6 and10,coinfection with v30%NS3alone; lanes7and 11, coinfection with vNS2B plusv30%NS3. Other controls: lane 2, infection with vSC8; lane 3, infection with vNS5 showing the authentic NS5. All radiolabeled cell lysates were
immunoprecipi-tatedby theanti-NS5serum.Protein sizemarkersareshown in lane 1 andindicated inkilodaltonsonthe left.
within NS5. Recombinants that expressedNS4B-NS5 trun-cated atthe C terminus tothe size of50%NS5or 22%NS5
were constructed and used for coinfection with
vNS2B-30%NS3, whichwasshown toprovide the NS2B-NS3
pro-tease activity (14). Figure 5 shows that coinfection yielded the predicted shortened NS5 band of50%NS5 or22%NS5
resulting frompropercleavageattheNS4B/NS5junction of the truncated NS4B-NS5 polyprotein.
Functionalsimilarity between NS2B-NS3 proteases of
den-gue and yellow fever viruses. Comparison of sequences
between DEN4 and yellow fever virus indicates that the amino acid homology is 37% for NS2B and 50% for the protease domain ofNS3 (consisting of the N-terminal 180 amino acids). We wereinterested in comparing the
NS2B-NS3 protease activities of dengue virus and yellow fever virus for cleavage at the DEN4 NS4B/NS5 junction. A recombinant vaccinia virus expressing yellow fever virus
NS2B-32%NS3,corresponding approximately in sizeto
den-gue virus NS2B-30%NS3, was constructed and used for
coinfection with vNS4B-NS5. The results (Fig. 6) indicate that both theNS2B-NS3 proteaseofyellowfever virus and that ofdengue virus cleaved the DEN4NS4B-NS5 polypro-teinwithhigh efficiency, sincethepolyproteinprecursorwas
mostly processed. This resultsuggests thatbothviral
prote-ases cleaved the NS4BINS5junction, presumably through the recognition ofthe cleavagesequence RR/G, despitethe significant sequence divergence that exists betweendengue virus andyellowfever virus proteases.
DISCUSSION
The current study examined the requirements for the
processing of DEN4 nonstructural proteins NS4A, NS4B, and NS5 from the polyprotein precursor. The sequences at
thecleavagesites that define theseproteins predictthattwo different proteolytic events are involved: the viral
NS2B-NS3 protease isresponsible forNS3/NS4A and NS4BINS5 cleavages, and asignal peptidase-like protease cleaves
be-
30-22°,oN55
[image:5.612.358.532.83.332.2]coinfection - + +
FIG. 5. Verification ofpropercleavageatNS4B/NS5. CV-1cells
were infected with recombinant vaccinia viruses that expressed C-terminally truncated NS4B-NS5. Cleavage at the NS4B/NS5 junctionwastestedby coinfection withvNS2B-30%NS3. Anti-NS5 serum wasused forimmunoprecipitation of radiolabeledlysatesof infected cells. Lanes: 1, vNS4B-22%NS5; 2, vNS4B-22%NS5 plus vNS2B-30%NS3; 3, vNS4B-50%NS5; 4, vNS4B-50oNS5 plus vNS2B-30%NS3. Protein size markers areindicated in kilodaltons onthe left.
tween NS4A and NS4B. Processing of the
NS2B-NS3-NS4A-NS4B-NS5 polyproteinappears tobe efficient in the recombinant vaccinia virus expression system. Incontrast,
recombinant vaccinia virus vNS3-NS4A-NS4B-NS5
ex-pressed only the uncleaved polyprotein. In an experiment
not shown, the addition of NS2B in trans, achieved by coinfection of cells with vNS3-NS4A-NS4B-NS5 and vNS2BorvNS2B-30%NS3, restored cleavage atthe NS4B/
NS5 junction because properly cleaved NS5 was readily
detected.However, cleavageattheNS3/NS4Ajunctionwas
notobserved in the same experiment, as indicated by the absence of NS3. In contrast, NS3 was detected when vNS2A-NS2B was used for coinfection. In this regard,
NS2A-NS2B may be more efficient than NS2B alone at
pairing with NS3 to produce the viral protease activity. Cleavage at NS3/NS4A apparently takes place efficiently when NS2B ispresentin cis butinefficientlywhen NS2B is provided in trans. These observations suggest that the functional requirement for components of the NS2B-NS3 viralproteasemaydifferforcleavagesatthe NS3/NS4Aand NS4B/NS5 junctions, althoughboth sitesappeartoshare the
consensus cleavage sequence. The requirements for
cleav-age atthe NS4B/NS5 junction were studied by expressing
polyproteinNS4A-NS4B-NS5 orNS4B-NS5 inacoinfection
procedure. EfficientprocessingofNS5 fromeither polypro-teinwas observed when both NS2Band NS3 weresupplied
intransbutnotwith either alone. Thisobservationprovides directevidence that the twocomponents ofthe viral
prote-ase canboth actintrans.
Processing between NS4A and NS4B occurred in the
92-I
1 2 3 4
210-_00 -NS5
OM-- 50%NS5
VOL.66, 1992 1539
on November 10, 2019 by guest
http://jvi.asm.org/
1540 CAHOUR ET AL.
vNS4B-NS5
I ., I
(vNS2B-NS3)
210--NS4B-NS5
_ _0M _Wm* -NS5 92
-69- *
1 2 3 4 5 6
FIG. 6. ComparisonofcleavagesatNS4B/NS5 junction by
den-guevirus NS2B-NS3andby yellowfevervirus NS2B-NS3. Exper-iments were carried out essentially as described forFig. 4 and 5. Complementation was carried out by coinfection of cells with vNS4B-NS5 plus vNS2B-30%NS3 of DEN4orvNS2B-32%NS3 of yellow fever virus. Lane 4, without coinfection; lane 5, with vNS2B-30%NS3 of dengue virus; lane6, with vNS2B-32%NS3 of yellow fever virus; lane 1, protein size markers, indicated in kilodaltonson the left. Other controls: lane2, vSC8;lane 3,vNS5. The bandmigratingbetween NS4B-NS5and NS5 isbackground.
absence of the NS2B-NS3 viral protease, as expression of
NS4A-NS4B-NS5 polyprotein yielded NS4B-NS5. The
un-cleaved precursor polyprotein NS4A-NS4B-NS5 was also
detected, and in some gels the level of the precursor was
equaltothe level of the cleaved NS4B-NS5 product (Fig. 3, lane 6). This finding indicates that cleavage atNS4A/NS4B
wasapparently less efficient thanthatoccurring duringother
signal-directed cleavages of the DEN4 virus polyprotein, such as at the pre-M/E and E/NS1 junctions, for which uncleaved precursors were notdetected (48). Experiments to demonstrate the requirement of the putative signal se-quencefor processing ofNS4A-NS4B-NS5 bv using an in
vitro translation procedure in the presence of microsomal membranes have notyieldedadefinitive result. A
recombi-nant vaccinia virus expressing NS4A-NS4B-NS5 polypro-teinthat lacks 17 aminoacids of theputative signal preceding the NS4A/NS4B junction was analyzed. Unexpectedly,
cleavageatNS4A/NS4Bwasnotcompletely abrogated (data
not shown). Since NS4A consists of mostly hydrophobic aminoacids, it is temptingto speculate that another portion ofNS4A, upstreamof the deletion, could serveas a
surro-gate signal and allow a low level of NS4A/NS4B cleavage. Interestingly, cleavage atNS4A/NS4B did notoccurwhen polyprotein NS3-NS4A-NS4B-NS5 was expressed. On the
other hand, in a previous study the results showed that
NS4A/NS4B cleavage appearedto occurwhen
NS3-NS4A-84%NS4B wasexpressed (14). One interpretation for these
findings is that thepresence of uncleaved NS3 and NS5 in
the molecule prevents proper recognition of the putative
signal sequence by a signal peptidase-like protease. This observation suggests that cleavage at the NS3/NS4A or
NS4B/NS5 junction takes place priorto the processing of
NS4A/NS4B.
Thus, in general, the NS2B-NS3 protease is responsible for processing at cleavage sites that consist of two basic amino acids followed by G, A, or S in the nonstructural
protein region. However, the cleavage efficiencies vary
accordingtothe individualcleavage sites. Variations of the amino acidsequenceatthe cleavage site mayinfluence the
cleavage
efficiency.
A deviation from this motif is theQR/G
sequence at the
NS2B/NS3
junction
found among all fourd%,ngue
virus serotypes.Previously,
we obtained evidencesuggesting
thatcleavage
atNS2B/NS3
containing
theQR/G
sequence followed
cleavage
atNS2A/NS2B,
which utilizestheRRISsequence
(14).
We also noted thatupon coinfectionof cells with
vNS3-NS4A-NS4B-NS5
andvNS2B-30%NS3,
processing
atNS3/NS4A,
where thecleavage
sequence isRK/S,
was less efficient than that atNS4B/NS5,
whichcontains the
cleavage
sequence RRIG. It is not knownwhether this
temporal
orderofcleavage
is determinedby
thesequence variation at the
cleavage
site alone. A kineticanalysis
of theprocessing
ofdengue
virus nonstructuralproteins
similartothatconductedwithyellow
fever virus(9)
should
help
to address thisquestion.
Atpresent,
it is notknown whetheramino acids outside the
cleavage
sequencemotifarealsoinvolved in
influencing
thecleavage
efficiency.
Comparison
ofthe NS4B/NS5cleavage junctions
of DEN4and
yellow
fever virus shows that both viruses utilize thesame
RR/G
sequence. Thisfinding
allowed us toemploy
DEN4 NS4B-NS5
polyprotein
tocomparethespecificity
ofproteases from both viruses. Under the conditions ofour
analysis, yellow
fever virus protease was able to processDEN4NS4B-NS5at an
efficiency
similartothatoccurring
inthe
homologous
dengue
virus system. Thisfinding
suggests
thatthe functional domains in both components of the viral
proteasemustbeconserved
despite
the fact that theoverall amino acidhomology
isonly
37% for NS2B and 50% for theproteasedomainof NS3. Studies
using
chimeric NS2B-NS3proteasesconstructed betweenDEN2and
yellow
fevervirushave
yielded
aninteresting
insight
into theenzyme-substrate
binding
and interaction between NS2B and NS3during
proteolytic processing
(31a).
A chimeric proteasethatcon-tains the
yellow
fever virus proteasedomain of NS3substi-tuting
for thecorresponding
sequence of DEN2 fails toprocess DEN2
NS2A/NS2B
andNS2B/NS3
cleavage
sites. On the otherhand,
another chimericconstructthat includesa C-terminal
portion
ofyellow
fever virus NS2B and theyellow
fever virus NS3proteasedomain isabletocleave theDEN2
NS2A/NS2B
andyellow
fever virusNS2BINS3
junc-tions. This indicates thatyellow
fever virusproteaseactivity
requires
proper interaction betweenhomologous
yellow
fever virus NS2B and
NS3, presumably through
the forma-tion of acomplex. Thus,
these results and others areconsistent with ourearlier
finding
thattheprotease domain of NS3 is necessary butnot sufficient fortheviral proteaseactivity.
Others have identified conserved sequences inflavivirus NS3 with
homology
toserineproteases,including
theproposed
catalytic
trial and substratebinding
domains(1,
15).
Studiestoidentify
suchconserved sequencesinNS2B,
andtofurther
assign
theirfunctional role incleavage,
remainto be
completed.
The results in this report and our
previous study
showed that the viral NS2B-NS3 protease isresponsible
for mostintergenic cleavages
inthenonstructuralprotein region (14).
Also,
this viralenzyme appearstobe involved incleavages
at severalsites
containing
theconsensuscleavage
sequence within the NS3 nonstructuralprotein.
Additionalcleavage
sites thatcontainthedibasic amino acidsequencemotifare
found at the anchC/C and
pre-M/M
junctions
in the struc-turalprotein region.
It was observed earlier thatpre-M
issubsequently
cleavedtoformmatureMduring
virusassem-bly
and release(38).
Similarly,
cleavage
ofthe C terminus from anchC to generate mature C was also observed(28).
Evidence is not available to suggest that
cleavage
at thepre-M/M
junction
is also mediatedby
NS2B-NS3 viralJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.88.263.79.234.2]PROCESSING OF DENGUE VIRUS NONSTRUCTURAL PROTEINS 1541
protease.Rather, it has been shown that cleavage ofpre-M
to generate matureMoccursinanacidic vesicular
compart-ment in the virus-infected cells (32). Consistent with the notion that viral protease is not involved in cleavage at pre-MIM is our observation that the mature M protein was not detected in the lysate of cells coinfected with vC-pre-M-E-NS1-NS2A and vNS2B-30%NS3 (4a). On the other hand, studieson processing of transientlyexpressedyellow fever virus proteins showed that yellow fever anchC/C cleavage appears to be mediated by the viral NS2B-NS3 protease (1). These studies provide evidence indicating the NS2B-NS3viral proteaseis also responsible for the delayed-type processing of the virion capsid protein.
ACKNOWLEDGMENTS
We thank L. Markofffor helpful discussions, R. Chanock for critical reading of the manuscript, and T. Heishman for expert editorial assistance.
REFERENCES
1. Amberg,S., andC. Rice. Personal communication.
la.Bazan, J. F., and R. J. Fletterick. 1989. Detection ofa trypsin-like serine protease domain in flaviviruses and pestiviruses. Virology 171:637-639.
2. Bell, J. R., R. W. Kinney, D. W. Trent, E. M. Lenches, L. Dalgarno, andJ. H. Strauss. 1985. Amino-terminal amino acid sequencesof structuralproteins of three flaviviruses. Virology 143:224-229.
3. Biedrzycka, A.,M. R. Cauchi,A. Bartholomeusz,J.J. Gorman, and P. J. Wright. 1987. Characterization of protease cleavage sitesinvolved in the formation of theenvelopeglycoproteinand three non-structural proteins of dengue virus type 2, New Guinea C strain. J. Gen. Virol. 68:1317-1326.
4. Boege, V.,F. X.Heinz, G. Wengler,andC.Kunz. 1983.Amino acid compositions and amino-terminal sequences of the struc-tural proteins of flavivirus European tick-borne encephalitis
virus. Virology 126:651-657. 4a.Bray, M. Unpublisheddata.
5. Castle, E.,U.Leidner, T.Nowak,G.Wengler, andG.Wengler. 1986. Primary structure of the West Nile flavivirus genome regioncodingfor all nonstructuralproteins. Virology149:10-26. 6. Chakrabarti,S.,K.Brechling,andB. Moss.1985.Vaccinia virus expression vector: coexpression of P-galactosidase provides
visualscreeningofrecombinant virus plaques.Mol. Cell. Biol. 5:3403-3409.
7. Chambers,T.J., C.H.Hahn, R.
Galier,
and C.M. Rice. 1990. Flavivirus genome organization, expression, and replication.Annu. Rev. Microbiol.44:649-688.
8. Chambers,T.J.,D. W.McCourt,and C.M. Rice. 1989. Yellow fever virusproteins NS2A,NS2B,and NS4B: identification and partialN-terminal amino acid sequenceanalysis.Virology169:
100-109.
9. Chambers, T. J., D. W. McCourt, and C. M. Rice. 1990. Production of yellow fever virus proteins in infected cells: identification of discrete polyprotein species and
analysis
of cleavage kineticsusing region-specific polyclonal antisera. Vi-rology 177:159-174.10. Chambers,T.J.,R.C.Weir,A.Grakoui,D. W.McCourt,J.F. Bazan,R.J.Fletterick,and C. M. Rice. 1990.Evidence that the N-terminal domain of nonstructural protein NS3 from
yellow
fever virus is a serine protease responsible forsite-specific
cleavagesin the viral polyprotein.Proc. Natl. Acad. Sci. USA 87:8898-8902.
11. Coia, G., M. D.Parker, G. Speight,M. E. Byrne, and E. G. Westaway. 1988. Nucleotide and
complete
amino acidse-quencesofKunjinvirus:definitive gene order and characteris-ticsof thevirus-specifiedproteins. J.Gen. Virol. 69:1-21. 12. Deubel, V.,R.M. Kinney,and D. W. Trent. 1988. Nucleotide
sequenceand deduced amino acid sequence of the nonstructural proteinsofdenguetype 2virus,Jamaicagenotype:
comparative
analysisof thefull-lengthgenome. Virology165:234-244.
13. Falgout,B.,R.Chanock,andC.-J.Lai.1989. Properprocessing ofdengue virus nonstructural glycoprotein NS1 requires the N-terminal hydrophobic signal sequenceand the downstream nonstructuralproteinNS2A.J. Virol. 63:1852-1860.
14. Falgout, B., M. Pethel, Y. Zhang, and C.-J. Lai. 1991. Both nonstructural proteins NS2B and NS3 are required for the proteolytic processingofdenguevirus nonstructuralproteins.J. Virol.65:2467-2475.
15. Gorbalenya,A.E.,A. P.Donchenko,E. V.Koonin,and V. M. Blinov. 1989.N-terminal domains ofputativehelicases of Flavi and Pestiviruses may be serineproteases. Nucleic Acids Res. 17:3889-3897.
16. Gubler, D. J. 1989. Aedes aegypti and Aedes aegypti-borne
disease control in the 1990s: top down or bottom up. Am. J. Trop. Med.Hyg.40:571-578.
17. Hahn,Y. S.,R.
Gafler,
T.Hunkapiller,J.M.Dalrymple, J.H.Strauss,and E.G. Strauss.1988. Nucleotide sequenceofdengue 2 RNA andcomparison of the encoded proteinswith thoseof otherflaviviruses.Virology162:167-180.
18. Hori, H.,and C.-J. Lai. 1990. Cleavageofdengue virus NS1-NS2A requires anoctapeptide sequenceat the C terminus of NS1. J. Virol.64:4573-4577.
19. Irie, A.,P. M.Mohan,Y.Sasagnri,R.Putnak,and R. Padman-aban. 1989. Sequenceanalysis of cloned dengue virus type 2 genome(NewGuinea-C strain). Gene 75:197-211.
20. Kleid,D.G.,D.Yansura,B.Small,D.Dowbenko,D.M.Moore,
M. J. Grubman, P. D. McKercher, D. 0. Morgan, B. H.
Robertson, and H. L. Bachrach. 1981. Cloned viral protein
vaccine for foot-and-mouth-disease: responses in cattle and swine. Science 214:1125-1129.
21. Krausslich, H. G., and E. Wimmer. 1988. Viral proteinases.
Annu. Rev. Biochem. 57:701-754.
22. Laemmli,U.K.1970.Cleavageof structural
proteins during
theassembly of the head ofbacteriophage T4. Nature
(London)
227:680-685.
23. Lai, C.-J., B. Zhao, H. Hori, and M. Bray. 1991. Infectious RNAtranscribedfrom stablyclonedfull-lengthcDNAof den-gue type4virus. Proc. Natl.Acad. Sci. USA 88:5139-5143. 24. Mackow, E.,Y.Makino,B.Zhao,Y.-M.Zhang,L.Markoff,A.
Buckler-White,M.Guiler,R.Chanock,andC.-J. Lai.1987. The nucleotide sequence ofdenguetype 4virus:
analysis
of genescodingfornonstructuralproteins. Virology159:217-228. 25. Mandl, C. W., F. X. Heinz, E. Stockl, and C. Kunz. 1989.
Genome sequence of tick-borne
encephalitis
virus (Westernsubtype) and comparative analysis of nonstructural
proteins
with otherFlaviviruses. Virology 173:291-301.
26. Markoff,L.1989.Invitro
processing
ofdengue
virusstructuralproteins: cleavage of the pre-membrane
protein.
J. Virol. 63: 3345-3352.27. Monath,T. P.1985.Flaviviruses,p. 955-1004.In B. N. Fields,
D. M. Knipe,R. M.Chanock,J. L.Melnick, B.Roizman,and R. E.Shope (ed.),
Virology.
RavenPress,NewYork. 28. Nowak, T., P. M.Farber, G. Wengler, and G.Wengler.
1989.Analysisof theterminal sequences of West Nile virus structural
proteinsandof thein vitrotranslation of these
proteins
allow theproposal of a complete scheme of the
proteolytic cleavages
involved in their
synthesis. Virology
169:365-376.29. Osatomi, K., and H.
Sumiyoshi.
1990.Complete
nucleotide sequence ofdengue
type 3 virusgenome RNA.Virology
176:643-647.
30. Pletnev, A. G., V. F. Yamshchikov, and V. M. Blinov. 1990. Nucleotide sequence of the genome and
complete
amino acid sequence of thepolyprotein
oftick-borneencephalitis
virus.Virology174:250-263.
31. Preugschat, F., C.-W. Yao, andJ. H. Strauss. 1990. In vitro
processing ofdengue 2 nonstructural
proteins
NS2A, NS2B,andNS3.J. Virol.
64:4364-4374.
31a.Preugschat,F. E.,M. Lenches,andJ. M. Strauss. 1991. Flavi-virus enzyme-substrate interactions studied withchimeric pro-teinases: identification of an
intragenic
locusimportant
for substraterecognition.
J. Virol. 65:4749-4758.32.
Randolph,
V.B.,G.Winkler,andV.Stollar.1990.Acidotropic
amines inhibit
processing
offlavivirusprM
protein.
Virology
VOL.66, 1992
on November 10, 2019 by guest
http://jvi.asm.org/
1542 CAHOUR ET AL.
174:450-458.
33. Rice, C. M., R. Aerbersold,D. B. Teplow, J. Pata, J. R. Bell, A. V.Vorndam,D. W.Trent,M. W.Brandiss, J.J.Schlesinger, and J. H. Strauss. 1986. Partial N-terminal amino acid se-quences of three nonstructural proteins of two flaviviruses. Virology 151:1-9.
34. Rice, C., E.M.Lenches, S.R.Eddy, S.J.Shin,R. L.Sheets,and J.H.Strauss. 1985. Nucleotide sequence ofyellow fever virus: implications for flavivirus geneexpression and evolution. Sci-ence229:726-733.
35. Rice, C. M., and J. H. Strauss. 1991. Production of flavivirus polypeptides by proteolytic processing. Semin. Virol. 1:357-367.
36. Ruiz-Linares, A., M.Bouloy, M. Girard,and A.Cahour. 1989. Modulations of thein vitro translationalefficiencies of yellow fever virus mRNAs: interactions between coding and noncoding regions. Nucleic Acids Res. 17:2463-2476.
37. Ruiz-Linares, A., A. Cahour, P. Despres, M. Girard, and M. Bouloy.1989.Processingof the yellow fever virus polyprotein: role of cellular proteases in the maturation of the structural proteins. J. Virol. 63:4199-4209.
38. Shapiro, D., W. E. Brandt, and P. K. Russell. 1972. Change involvingaviral membrane glycoprotein during morphogenesis of group B arboviruses. Virology 50:906-911.
39. Speight, G., G. Coia, M. D. Parker, and E. G. Westaway. 1988. Gene mapping and positive identification of non-structural pro-teins NS2a, NS2b, NS3, NS4b, and NS5 of the flavivirus Kunjin and their cleavage sites. J. Gen. Virol. 69:23-34.
40. Speight, G., and E. G. Westaway. 1989. Carboxyl-terminal analysis of nine proteins specified by the flavivirus Kunjin: evidence that only the intracellular core protein is truncated. J. Gen. Virol. 70:2209-2214.
41. Spindler, K. R., D. S. E. Rosser, and A. J. Berk. 1984. Analysis
of adenovirustransforming proteins from early regions 1A and 1B with antisera to inducible fusion antigens produced in Escherichiacoli. J. Virol. 49:132-141.
42. Strauss, J. H., E. G. Strauss, C. S. Hahn, R. Galier, W. R. Hardy, and C. M.Rice. 1987. Replication of alphaviruses and flaviviruses: proteolyticprocessingofpolyproteins, p. 209-226. In M. A. Brinton and R. R. Rueckert(ed.),Positive strand RNA Viruses.AlanR. Liss, New York.
43. Sumiyoshi, H., C. Mori, I. Fuke, K. Morita, S. Kuhara, J. Kondou, Y. Kikuchi, H. Nagamatu, and A. Igarashi. 1987. Complete nucleotide sequence of the Japanese encephalitis virusgenome RNA. Virology 161:497-510.
44. Wengler, G., G. Czaya, P. M. Farder, and J. H. Hegemann. 1991. In vitro synthesis of West Nile virus proteins indicates thattheaminoterminal segment of the NS3 protein contains the active centre of the protease which cleaves the viralpolyprotein aftermultiple basic amino acids. J.Gen. Virol. 72:851-858. 45. Westaway, E. G., M. A. Brinton, S. Y. Gaidamovich, M. C.
Horzinek, A.Igarashi, L.Kaariainen, D. K. Lvov, J.S. Porter-field, P. K. Russell, and D. W. Trent. 1985. Flaviviridae. Intervirology 24:183-192.
46. Wright,P.J.,M. R.Cauchi,and M. L.Ng. 1989. Definition of the carboxyl termini of the three glycoproteins specified by dengue virus type 2. Virology 171:61-67.
46a.Zhang, Y.,andC.-J. Lai. Unpublished data.
47. Zhao, B., E. Mackow, A. Buckler-White, L. Markoff, R. M. Chanock, C.-J. Lai, and Y. Makino. 1986. Cloning full-length dengue type4viral DNA sequences: analysis of genes coding for structural proteins. Virology 155:77-88.
48. Zhao, B., G. Prince,R. Horswood, K. Eckels, P. Summers, R. Chanock, and C. J. Lai. 1987. Expression of dengue virus structuralproteins and nonstructural protein NS1 by a recom-binant vaccinia virus. J. Virol. 61:4019-4022.
J.VIROL.
![FIG. 2.ofleft;bacteriaoutblue.electrophoresis,L-[35S]methionine-labeledpATH3/NS5-2;againstspecifickilodaltonsmunizationLanes: the Characterization of TrpE/NS5 fusion proteins and NS5- sera](https://thumb-us.123doks.com/thumbv2/123dok_us/1310047.84299/3.612.326.563.80.251/bacteriaoutblue-electrophoresis-methionine-labeledpath-againstspecifickilodaltonsmunizationlanes-characterization-fusion-proteins.webp)