0022-538X/88/030715-07$02.00/0
CopyrightC) 1988,American Society for Microbiology
A
Temperature-Sensitive
Mutation in a Herpes Simplex Virus Type
1
Gene Required
for Viral DNA Synthesis Maps to Coordinates
0.609
through
0.614 in
UL
MICHAEL E.
MARCHETTI,
COLTONA.SMITH,
AND PRISCILLA A. SCHAFFER*LaboratoryofTumor VirusGenetics, Dana-Farber Cancer Institute, and Department of Microbiology and Molecular
Genetics,
Harvard MedicalSchool, Boston,
Massachusetts 02115Received 19 October1987/Accepted3December1987
ts701 is atemperature-sensitivemutantofherpes simplex virus type 1 strain KOS induced byhydroxylamine
mutagenesis (C. T. Chu, D. S. Parris, R. A. F. Dixon, F. E. Farber, and P. A. Schaffer, Virology98:168-181,
1979). In the present study, the mutation rendering ts701 temperature sensitive was mapped to coordinates 0.609 through 0.614 in the UL region of the genome. At the nonpermissive
temperature,
ts701 (i) failed to inducethesynthesis of viral DNA, (ii) exhibited a dramatically reduced ability to drive replication of a plasmid containing the herpessimplex virus origin of viral DNA synthesis,oris,
(iii)generated no viral polypeptides of the late(72)
kinetic class, and (iv) produced virions with electron-translucent cores. Northern (RNA) blot hybridization demonstrated that two mRNAs-one of the I kinetic class and one of the'y
kinetic class-hybridized to a1.3-kilobase viral DNA fragment that rescued the mutation ints701. Basedon thephenotype andmapping ofts701, it is likely that its mutation lies in the gene specifying the 65,000-Mr DNA-binding protein(65KDBp)
recently described by Marsden et al.(H. S. Marsden, M. E. M. Campbell, L. Haarr,M. C.Frame, D. S. Parris, M.Murphy, R. G. Hope, M. T. Muller, and C. M. Preston, J. Virol. 61:2428-2437, 1987).The identification of virus-encoded proteins required for herpessimplex virus (HSV)type 1 (HSV-1)DNA synthesis has been accomplished to date by two markedly different procedures. First, studies of
temperature-sensitive
(ts) mu-tants have led to the identification of six distinct viralproteins thatareessential for viralDNAsynthesis(27). The
functions of only two of these
proteins,
the viral DNApolymerase (6) and the
major
DNA-binding protein ICP8 (32),have beendetermined.Recently, a meansofidentifying adenovirus proteins involved in viral DNA synthesis wasadapted forusewithHSV-1byChallberg (4).Thisprocedure utilizes clones of viral DNA fragments representing the
entire genome. These clones are assayed in transfection experiments in all possible combinations to determine whethertheproteins they encode cancollectively drivethe
replication ofa plasmid containing an HSV origin of repli-cation. With this procedure, sevengenes essential forviral
DNA
synthesis
have beenidentified andfinely mapped (M.Challberg, personal communication). Six of these seven genes were identified and mapped previously with ts mu-tants.Todate, nospecific functionhas beenascribedtofive of the seven viral proteins, although biochemical studies
suggestthat,inadditiontothepolymeraseandmajor
DNA-binding protein, thevirus may encode its owntopoisomerase (2, 17, 21), DNA primase (W. Ruyechan, personal
commu-nication), andorigin-binding protein(11).
Inthisreport, wedescribeamutantof HSV-1 strain KOS,
ts701, which is temperature sensitive for viral DNA
synthe-sis. We have mappedthe mutation rendering ts701 temper-ature sensitive to coordinates 0.609 through 0.614. The
location ofthe ts701mutation indicatesthatit lies in thegene
fortheonly essential viral DNAsynthesis protein forwhich no ts mutation has yet been identified (M. Challberg,
per-sonal communication). Several transcripts transverse the
*Correspondingauthor.
coordinates 0.609 through 0.614 (12), but only one is the ,B
kinetic class. It is 1.44 kilobases (kb) long and encodes a
protein of64 kilodaltons, asascertainedby in vitro transla-tion (12). This 64-kilodalton product maybe the
65,000-Mr
DNA-bindingprotein
(65KDBp)
describedby Marsdenetal.(20). Since all HSV genes encoding proteins directly in-volved in viralDNA synthesisareof the,1 kinetic class(27),
it is likely that ts701 is defectivein
65KDBP.
MATERIALS ANDMETHODS
Cells and viruses. Verocells werepropagatedasdescribed previously (32). ts701 and tsA24 are derivatives of HSV-1 strainKOS (5, 27).Mutantandwild-type virusesweregrown
andassayedas describedpreviously (32).
Plasmids.Formarkerrescueanalysis,theEcoRI library of KOS derivedby Goldin etal. (13)wasused. Thefirstseven
subclones ofEcoRI-Alisted in Table 1and shown in Fig. 1
(A-2 through 4ofA-1) were cloned into pUC8 by standard techniques (18). Bal 11 was derived by linearizing1 ofA-1
withHindIII,digestingitwith Bal 31nuclease,andligatingit
with HindIII linkers. Plasmids 9, 14, and 8 ofBal 11 were
derivedin the same manner, except that Bal 11 wasusedas
theparental plasmid. Plasmids Bal11-15, Bal 11-13,andBal
11-10 werederivedbylinearizing Bal11with
SalI,
digestingit with Bal 31 nuclease, and ligatingit with EcoRI linkers.
DNA-modifying enzymes and HindIII and EcoRI linkers were obtainedfrom New England BioLabs, Inc. (Beverly,
Mass.) and used asprescribed by themanufacturer. Nucleicacid isolation. Purified HSV-1 and bacterial
plas-mid DNAs were isolated as described previously (13, 18). Cytoplasmic RNAwas isolated fromNonidet P-40extracts ofVerocells as describedpreviously (18).
Markerrescuetest.Markerrescue experimentswere car-ried out as described byWeller et al. (32)with the
transfec-715
on November 10, 2019 by guest
http://jvi.asm.org/
716 MARCHETTI ET AL.
TABLE 1. Markerrescue analysis'
Marker rescue inexpt: Plasmid
1 2 3 4
pUC8 <1.1 x 10-4 <2.0 x 10-3 <9.0 X 10-4 <9.7 x 10-4
EcoRI-A 1.51
A-2 <1.1 x 1O-4
HindIII-K 9.0 X 10-5
A-1 1.01 0.36
1ofA-1 0.18
2ofA-1 <4.0 x 10-3
3 of A-1 ND
4of A-1 <3.3 x 10-3
Bal 11 0.13
9ofBal11 0.42
14ofBal 11 0.66
8 of Bal11 0.04
Bal11-15 <5.5 x 10-3
Bal 11-13 <1.1 x 10-3
Bal11-10 0.44
a Marker rescue analysis was conducted with the indicated plasmids appropriately linearized prior to transfection. The data indicate the percentage of the total transfection progeny found to be ts+. ND, Not determined.
tion protocol of Parris et al. (22), except that DNA was
precipitatedwith CaCl2for 30minand Vero cells were used. Southern and Northern (RNA) blot analyses. Restriction
enzyme-cleavedDNAsseparated by agarose gel
electropho-resis were transferred to nitrocellulose by the method of
Southern (18,28). Forthe experiment showninFig. 4, pUC8 was used as probe.
Formaldehyde gel electrophoresis ofRNAsandNorthern
blot hybridization were performed as described previously
(8). FortheexperimentshowninFig.6,8 of Bal11 was used as aprobe.
a b
E___
E
Probesfor both Southernand Northernblot analyseswere
nick translated with 32P-labeled dCTPand dGTP as
previ-ously described (18).
Polypeptideanalysis. Lysatesof radioactivelylabeled cells wereanalyzed by sodium dodecylsulfate-polyacrylamide
gel,
electrophoresis by the method of Laemmli(16) as modified
byManservigietal. (19). Cellswereinfectedat amultiplicity of infection of20 PFUpercell, incubatedat34 or 39.7°Cfor various times with 100 ,uCi of
[35S]methionine
per ml, and harvested asdescribed byDeLuca etal. (7).Viral DNAphenotypes. The viral DNA phenotype of ts701
b'
a'cloris
oris c a 4---11----471E -
-L
H
EcoRIA (21.2) A-2 (5.46)
H
H
H S
i I
S SS s
S E
l
-H S
H4 H
F II
H S
H S
H E
I
H E
i
H E
I
HindIII K (9.26)
E
-4
A-I
(6.49)of A-l (3.8)
3,4 of A-I (0.8)
2 of A-I (1.9) BaI I I (3.3)
9 of Bal I (2.9) 14 of Bal I (2.0)
8 of Bal I1 (1.3) Bal 1 1-15 (1.1)
Bal 1 1-13 (2.7)
Bal I 1-10 (3.2) FIG. 1. Subclones ofEcoRI-A used inmarkerrescuetestswith ts701. At thetopof thefigure isadiagram of the HSV-1genomeillustrating
thelocationof the EcoRI A fragment. Thelocation of the diploidorisin thecrepeatsis alsoshown(oris). Beneath the expandedEcoRIA
fragmentareshown thesubclones used inmarkerrescuetests.E, EcoRI; H,HindIII;S, Sall. Numbers of parenthesestothe rightof plasmid designations aresizes of thefragmentsinkilobases.
H
i
----J. VIROL.
-.Li
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.88.539.427.687.2]MAPPING OF HSV-1 KOS MUTANT ts701 717 was determined in Vero cells infected at a multiplicity of
infectionof20 PFU percellat34or39.7°C by the method of
Aron etal. (1).
oris
replication assay. Vero cells (3 x 106) in 100-mm2plates were transfected with 0.1 mg of pUCori (a pUC8 plasmid bearing
oris
as a BamHI fragment) and 18 mg of salmontestis DNA by the procedure of Weller etal. (32).After 1 hofincubation of 37°C,the cells wereinfectedwith 20 PFUofmutant orwild-type viruspercell andincubated for anadditional 18 h atvarious temperatures. TotalDNA was then extracted and cleaved withBamHIand DpnI, and
fragmentswereresolvedon an0.8%agarosegelasdescribed
previously (18). The gelwas blotted ontonitrocellulose and probedwithpUC8 DNA asdescribed previously (28).
Electron microscopy. Vero cells wereinfectedwith 5 PFU
of eitherts701 orKOSpercellandincubatedat34 or39.7°C for 36 h. At this time, the monolayers were washed twice
with cold Tris-buffered salineand fixed with2.5%
glutaral-dehyde. Monolayerswereprocessed for thin sectioning and examined by electron microscopy as described
previously
(26).the
-Yl
polypeptide ICP5. ts701 and tsA24 generated the same amounts of the early polypeptide thymidine kinase(ICP36)asdid KOS at 39.7°C.
Viral DNA synthesis. To date, all HSV ts mutants that fail toinducethesynthesis of
Y2
polypeptides at thenonpermis-sivetemperaturehave been defectivein viral DNAsynthesis
atthistemperature (27). Wethusdeterminedthe viral DNA phenotype of ts701 at 39.7°C. ts701 failed to induce the
synthesis of detectable viral DNA at 39.7°C, whereas it induced -85% of wild-typelevels at340C (Fig. 3).ts701 was as efficient as wild-type virus in inhibiting cellular DNA
synthesisatboth temperatures.
Tofurtherassess theability ofts701 toinduce viralDNA
synthesis, we tested its ability to drive the replication ofa
340C
0 N
4 N u
39.7 5C
X XN o O
RESULTS
Marker rescue analysis. Using gel-purified EcoRI
frag-mentsin markerrescuetests, Chuetal. showed previously
that the mutation in ts701 lies within EcoRI-A (5). We
confirmed this finding by using cloned fragments; only
EcoRI-A yielded ts+ recombinants (data not shown). Sub-fragments ofEcoRI-A were thencloned and used in
subse-quent marker rescue experiments (Fig. 1 and Table 1).
Subclonesthatrescued thets701mutation included A-1;1of
A-i;
Bal 11; 9, 14, and 8 of Bal 11; and Bal 11-10. 8 of Bal11proved to be the smallest rescuing fragment. This latter result, combined with the fact that Bal 11-10 alsorescued, localized the ts701 mutation to a 0.7-kb length of DNA
betweencoordinates0.609and0.614 on thephysicalmapof HSV-1.
Two ts+ recombinants derived from the marker rescue
experiment with 8 of Bal11 were analyzed with respect to
plating efficiency inVero cellsatthepermissive (34°C) and nonpermissive (39.7°C) temperatures. The platingefficiency (PFU per milliliter at 39.7°C/PFU per milliliter at 34°C) of KOS was 0.67, whereas the ts+ recombinants exhibited plating efficiencies of0.40 and 0.43. By contrast,theplating
efficiency
ofts701was 2.2 x10-'.
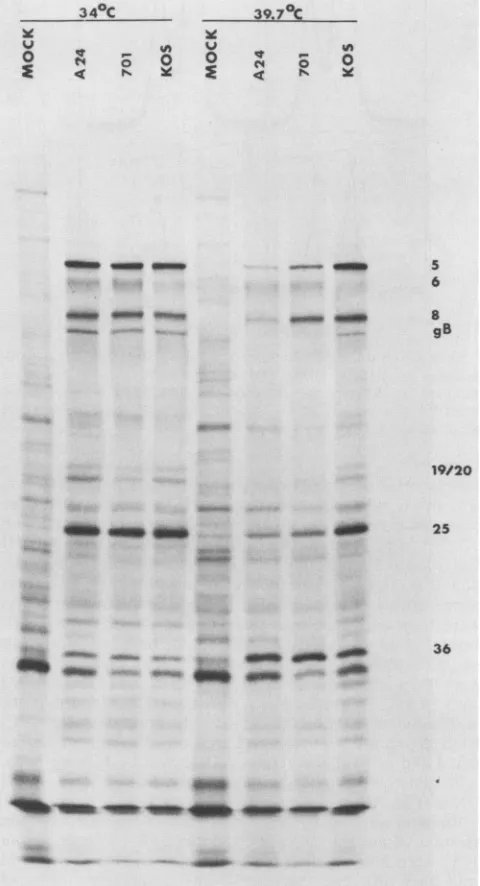
Polypeptideprofileof ts701 at39.7°C. We nextdetermined the spectrum of viral polypeptides induced by ts701 at the
permissiveandnonpermissivetemperatures. tsA24, a
DNA-negative ts mutant defective in ICP8 (complementation
group 1-1), andKOS, thewild-type virus, were included as
controls in these experiments. The three viruses exhibited virtually indistinguishable polypeptide profiles at 34°C (Fig. 2).
At39.7°C, ts701,liketsA24, (i) failedtoinducedetectable
levels of late polypeptides of the _Y2 class
(polypeptides
whose synthesis is absolutely dependent upon viral DNA
synthesis) and (ii) induced markedly reduced levels of late
polypeptides ofthe
-Yl
class(polypeptidessynthesized in theabsence ofviral DNAsynthesisbut whosesynthesisreaches maximum ratesonly whenviral DNA replication is allowed to occur). This phenotypeis exemplifiedby the absenceof ICP19 and ICP20(Y2)andbythereduced levels of gB, ICP5,
andICP25
(-Yl)
in mutant-infectedcells. Although theprofilesofts701 and tsA24 werequalitativelysimilar at39.7°C,ts701
induced higherlevels ofthe early polypeptide ICP8 and of
-mmW _-_
- mo
5
6
gB
19/20
25
36
-do-W. _ - d
u-|
_W
_.OM
- 41FIG. 2. Polypeptideprofile ofts701. Verocellswereinfectedat 34or39.7°Cwith the indicatedvirusesat amultiplicityof infection of20 PFUpercell,labeledwith100,uCiof[35S]methionineper ml from 5 to 18 h postinfection, and processed for sodium dodecyl sulfate-polyacrylamidegelelectrophoresis. Thenumbersattheright indicateinfected-cellpolypeptides.
VOL. 62,1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.314.554.227.670.2]718 MARCHETTI ET AL.
thickening of the nuclear membrane (Fig. 5D), a character-istic of KOS-infected cells (Fig. 5A). These findings are consistent with theDNA-negative phenotype and
polypep-tide profile ofts701-infected cells at 39.7°C (Fig. 2 and 3)
(26).
mRNAs that map to coordinates 0.609 through 0.614. To determine the sizes and kinetic classes of transcripts en-codedby sequences that contain the mutation in ts701, we extracted cytoplasmic RNA at 6 hpostinfection from
cyclo-co heximide- and phosphonoacetic acid (PAA)-treated Vero
C:
o cells infected with HSV-1 KOS and at 11 h
postinfection
o from untreated Vero cells also infected with HSV-1 KOS. RNAs weresubjected to Northern blot analysis by using as
C aprobe the smallest fragment that rescued the mutation in
ts701,
the 1.3-kbfragment,
8 of Bal 11(Fig. 6).
Twounique
,< mRNAs wereidentifiedinKOS-infected cellsintheabsence ofdrug. The largertranscript (-2.7kb) was not synthesized 3 inKOS-infected cells treated with either the DNA synthesis o inhibitorPAAor aninhibitor ofprotein synthesis,
cyclohex-3 imide,indicating that this transcript is of the -y kinetic class. The smaller transcript (-1.4 kb) was synthesized in the presence of PAA but not in the presence ofcycloheximide,
indicatingthat itis of the 1 kinetic class.SinceallHSV genes
encoding proteins directly involved inDNAsynthesisareof the 1 kinetic class(27), itisperhaps morelikelythat the ts mutationin ts701 lies in thegenespecifying the1 transcript.
DISCUSSION
In vitro replication assays (4; M. Challberg, personal
communication)havepredictedtheexistence ofsevengenes Fraction Number
FIG. 3. Viral DNAphenotypeof ts701. Cesium chloridedensity gradient profiles of ts701- and KOS-infected cell lysates were
obtained as described previously (1). The peak of viral DNA occurred at a density of 1.727 g/cm3, and that the cellular DNA occurredat 1.699g/cm3.
plasmid bearing the HSV origin ofreplication, ori5 (4).oris
acts in cis when the appropriate HSV gene products are
provided in trans(29, 30). Vero cells weretransfected with
pUCori, a plasmid consisting of pUC8 into whose BamHI
site
oris
hasbeen cloned(N. DeLuca, personal communica-tion)(Fig. 4). Cellsweretheninfected with virusesatvarious temperaturesand harvestedat18 hpostinfection.The DNAs were then extracted and cleaved with BamHI and DpnI.DpnIdigestionwasusedtodifferentiate between unreplicat-ed and replicated pUCori DNAs (23). The digested DNAs
were then separated in agarose gels, blotted, and probed
with pUC8. At 34°C, KOS, ts701, and tsA24 drove the replication of pUCori efficiently. By contrast, at 39.7°C, KOS drove the replication of pUCori efficiently, whereas tsA24 and ts701 did not. Thus, as shownby twomethods, ts701 is defective in itsabilitytoinduce thesynthesis of viral DNA at39.7°C.
Electron microscopy. Electron photomicrographs ofthin sections ofmutant-infected cells after 36hof incubation of 34°C were indistinguishable from those of thin sections of
wild-type virus-infected cells at this temperature (data not shown). At 39.7°C, however, infection with ts701 did not lead tothe production ofmature, enveloped, extracellular particles orintracellular virions with electron-dense cores,
as did infection with KOS (Fig. 5A, B, and C); only
translucent, DNA-less, incomplete nucleocapsids were seen
innuclei(Fig. 5DandE). Additionally, ts701failedtoinduce
340
0
11-.
z i
o 0 o f4
v E Y b <
PUC 8
39.70
0
I-z
0 u
0
:E _
0 o0 Y P% <
I..
I
FIG. 4. Southern blotanalysisof theorisreplicationassay.Cells
were transfected with pUCori (a pUC8 plasmid bearing oris as a
BamHI fragment),infected at theindicated temperatureswith the
indicated virusesatamultiplicityof infection of 20 PFUpercell,and harvestedat18 hpostinfection.Total DNAwasextracted,cutwith
BamHI andDpnI, runon anagarosegel, blotted,andprobedwith pUC8.ThesequencesdetectedarethepUC8sequencesofpUCori.
0-I2
0
%._
E
Ur
J. VIROL.
i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.67.298.69.397.2] [image:4.612.324.558.398.662.2]MAPPING OF HSV-1 KOS MUTANT ts701
.-.KX
-. E
0~~
ytr:'_* RI ,-~~~~~~~~~~~~~~~~
_',-9.I I
- K-*~
r}
. ../
aov
Ar V
,.. 6 t~~~~~~'
W\f*)t91',t4 S*
*-~~~a
* \ .:-'4
0--X
Vs
- .1
I. A" Q.,Gv
i.
..4.
.A ....
~~c~...
FIG. 5. Electronphotomicrographs of thin sections of Vero cells infectedwith 5 PFU of KOS(A, B,andC)orts701(DandE)percell
for 36 hat39.7°C. Mature, enveloped, extracellular particles with dense (full)cores(-*)canbeseeninpanel A,ascan aregionofthickening
of the nuclear membrane (-*). Full(DNA-containing) particles (-*)andparticles containing internalstructures butlackingdensecores
(partial, DNA-less particles) (-) can be seen in panels B and C. A particle withno internal structures (->) can be seen in panel B.
ts701-infectedcells(panels DandE) characteristicallycontainednofullparticles. Inaddition, nomature,enveloped particleswere seenin
the extracellularspace. Nuclei, however, contained smalltomoderate numbers ofpartial nucleocapids ().
whose productsareessential for viral DNA synthesis. Of the
seven, onlytwohave been characterizedextensively, anda third has beencharacterizedtoalesserextent.These include the DNA polymerase (6), the major DNA-binding protein ICP8(32),and65KDBP(20), although other proteins directly involved in viral DNA synthesis have been reported to be inducedfollowingHSV infection. The latterproteinsinclude
atopoisomerase (2, 17, 21),aDNAprimase (W. Ruyechan,
personal communication), andanHSV-induced proteinthat binds specificallyto
oris
(11).HSV DNAalso encodes severalenzymes involvedin the
generation of DNA precursor molecules. These include a
thymidine kinase(9),analkaline nuclease(15, 24),a ribonu-cleotide reductase (10),auracylDNAglycosylase (3),anda
dCMP deaminase (25). Deletion of thegenes for the former threeenzymesdoesnotaffect theviabilityof the virus in cell cultures (S. Weller, personal communication).
Prior to this report, ts mutations had been described in only six of the seven essential DNA replication genes identified by the in vitro replication assay. The characteri-zation of ts701 now brings this number to seven. ts701 is temperature sensitive for viral DNA synthesis, and its
mu-tation maps to coordinates 0.609 through 0.614. Northern blot analysis indicates that at least two mRNA transcripts
A
:4I
0
p
719 VOL.62, 1988
.-5 0
to
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.148.453.69.541.2]720 MARCHETTI ET AL.
0~~~
o
~~~v
40 0 0
Fw
28SSt
18
S*-FIG. 6. Northern blot analysis of KOS-infected cells with the
smallestfragment (8 of Bal 11) thatrescuedthemutationin ts701as a probe. The larger transcript (y) was not synthesized in KOS-infectedcells treated with either theDNA synthesisinhibitor PAA or aninhibitor ofprotein synthesis, cycloheximide (CYCLO.). The
smaller (1) transcript was synthesized in the presence of PAA but not cycloheximide.
traverse these coordinates (Fig. 6). One of these is a y
transcript which probably corresponds to the late
tran-script(s) mapped between coordinates 0.610 and 0.633 by Frinketal. (12). It isunlikely thatthis transcript encodes a
protein involved in DNA synthesis, since viral DNA repli-cation isa prerequisite for its production (14) (Fig. 6). The
othertranscripttraversingcoordinates 0.609 through 0.614 is ofthe kinetic class and probablycorrespondstothe 1.4-kb early transcript mapped by Frink et al. (12). By in vitro translation, it was found that this transcript encodes a
64-kilodalton protein which likely corresponds to 65KDBP described by Marsden et al. (20). It has been shown that
65KDBP
specifically associates with the HSV-encodedpoly-merase, asdemonstrated by theobservation that itcopurifies
with the polymerase duringcolumn chromatography andon
immunoaffinity columns (31; M. Gallo, M. Muller, H. Mars-den, and D. Parris, personal communication). Using interty-pic HSV-1 x HSV type 2 recombinants and type-specific antibodies, Marsden et al. mapped the gene encoding
65KDBP
to coordinates 0.574 through 0.682 (20).These coordinates encompass the area to which we have
mapped the mutation in ts701. What function 65KDBP per-formsisunknown,but thisreport suggeststhatitis essential for viral replication. However, its amino acidsequence has been reported to share homology with those ofeucaryotic primases (H. Marsden, personal communication). We are currently constructing mutants containing deletion muta-tions in the gene which is mutated ints701 and which most likely encodes
65KDBp.
Thesemutants should prove useful in elucidating the role of 65KDBP inthe synthesis of HSV DNA.ACKNOWLEDGMENTS
We thank N. DeLucafor plasmids and for valuable discussions, R. Byrn, N. DeLuca, and W. Sacks forassistance in many of the experiments, and M. Kaveny formanuscript preparation.
This investigation was supported by Public Health Service grant CA-21082 from the NationalCancerInstitute. C.A.S. was supported by National Science Foundation Graduate Fellowship RCF-84-50074.
LITERATURE CITED
1. Aron, G. M., D,J. M. Purifoy,and P.A. Schaffer. 1975. DNA synthesisand DNApolymeraseactivityofherpessimplex virus type 1 temperature-sensitive mutants. J. Virol. 16:498-507. 2. Biswal, N., P. Feldan, and C. C. Lery. 1983. A DNA
topoisom-erase activity co-purifies with the DNA polymerase byherpes simplex virus. Biochim. Biophys. Acta 740:379-389.
3. Caradonna, S. J., and Y. C. Cheng. 1981. Induction of uracil-DNA glycosylase and dUMP nucleotidohydrolase activity in herpes simplex virus-infected human cells. J. Biol. Chem. 256: 9834-9837.
4. Challberg, M. D. 1986. A methodfor identifyingthe viral genes required for herpesvirus DNA replication. Proc. Natl. Acad. Sci. USA 83:9094-9098.
5. Chu, C. T., D. S. Parris, R. A. F. Dixon, F. E. Farber, and P. A. Schaffer. 1979. Hydroxylamine mutagenesis ofHSV DNAand DNA fragments: introduction of mutationsintoselected regions of the viral genome. Virology 98:168-181.
6. Coen, D. M.,D. P. Aschman, P. T. Gelep, M. J. Retondo, S. K. Weller, and P. A. Schaffer. 1984. Fine mapping and molecular cloning of mutations in the herpessimplex virusDNA polymer-ase locus. J. Virol. 49:236-247.
7. DeLuca, N. A., M. A. Courtney, and P. A. Schaffer. 1984. Temperature-sensitive mutants in herpes simplex virus type 1 ICP4 permissive for early gene expression. J.Virol. 52:767-776. 8. DeLuca, N. A., A. M. McCarthy, and P. A. Schaffer. 1985. Isolation and characterization of deletion mutants of herpes simplex virus type 1 in the gene encoding immediate-early regulatoryprotein ICP4. J. Virol. 56:558-570.
9. Dubbs, D. R., and S. Kit. 1964. Mutantstrains of herpessimplex virus deficient in the thymidine kinase-inducing activity. Virol-ogy 22:493-502.
10. Dutia, B. M. 1983. Ribonucleotidereductase induced by herpes simplex virus has a virus-specified constituent. J. Gen. Virol. 64:513-521.
11. Elias, P., M. E.O'Donnel, E. S. Mocarski, and I. R. Lehman. 1986. A DNA binding protein specific for anorigin ofreplication of herpes simplex virustype 1. Proc. Natl. Acad. Sci. USA83: 6322-6326.
12. Frink, R. J., K. P. Anderson, and E. K. Wagner. 1981. Herpes simplex virus type 1 HindIll fragment L encodes spliced and complementary mRNA species. J. Virol. 39:559-572.
13. Goldin, A. L., R. M. Sandri-Goldin, M. Levine, and J. C. Glorioso. 1981. Cloning of herpes simplex virus type 1
se-quences representingthe whole genome. J. Virol. 38:50-58. 14. Holland, L. E., K. P. Anderson, C. Shipman, Jr., and E. K.
Wagner. 1980. Viral DNAsynthesisis requiredfor the efficient expression of specific herpes simplex virus type 1 mRNA species. Virology 101:10-24.
15. Keir, H. M., and E. Gold. 1963. Deoxyribonucleicacid nucleo-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.117.255.72.438.2]tidyltransferase anddeoxyribonucleasefrom cultured cells in-fected withherpes simplex virus. Biochim. Biophys. Acta 72: 263-276.
16. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
17. Leary,K., and B. Franke. 1984. Theinteraction of a topoisom-erase-like enzyme from herpes simplex virus type 1-infected cellswith non-viral circular DNA. J. Gen. Virol. 65:1341-1350. 18. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
19. Manservigi, R., P. Spear, and A. Buchan. 1977. Cell fusion inducedby herpessimplexvirus ispromoted andsuppressed by different viral glycoproteins. Proc. Natl. Acad. Sci. USA 74: 3913-3917.
20. Marsden, H. S.,M. E. M.Campbell, L.Haarr,M. C. Frame, D.S. Parris,M.Murphy,R.G.Hope, M. T. Muller,andC.M. Preston. 1987. The 65,000-Mr DNA-binding and virion trans-inducingproteins ofherpes simplex virus type 1.J. Virol. 61: 2428-2437.
21. Muller,M.T., C.S.Bolles,and1). S.Parris. 1985. Association of typeIDNA topoisomerasewithherpes simplex virus. J. Gen. Virol. 66:1565-1574.
22. Parris,D.S., R. A.F.Dixon,and P. A.Schaffer. 1980. Physical mapping of herpes simplex virus type 1 tsmutantsby marker rescue: correlationof the physical and genetic maps. Virology 100:275-287.
23. Peden,K. W.C., J.M.Pipas, S. Person-White, andD. Nathans. 1980. Isolation ofmutantsofananimal virus in bacteria. Science
209:1392-1396.
24. Preston, C. M., and M. G. Cordingley. 1982. mRNA- and DNA-directed synthesis of herpes simplexvirus-coded exonu-cleasein Xenopus laevis oocytes. J. Virol. 43:386-394. 25. Rolton, H. A., and H.M. Keir. 1974. Deoxycytidylate
deami-nase:evidence for a new enzyme in cells infectedbythevirus of herpes simplex. Biochem. J.143:403-409.
26. Schaffer, P. A., J. P. Brunschwig, R. M. McCombs, and M. Benyesh-Melnick. 1974. Electron microscopic studies of temper-aturesensitivemutantsofherpes simplex virus type 1.Virology 62:444-457.
27. Schaffer, P. A., V. G. Preston, E. K. Wagner, and G. B. Devi-Rao. 1987. Genetic maps, vol. 4, p. 93-98. Cold Spring HarborLaboratory, ColdSpring Harbor, N.Y.
28. Southern, E.M. 1975. Detection of specific sequences among DNAfragments separated by gel electrophoresis. J. Mol. Biol. 98:503-517.
29. Stow, N.D.1982. Localization of an origin of DNA replication within theTRs/IRs repeated region of the herpes simplex virus type 1genome. EMBO J. 1:863-867.
30. Stow, N. D.,and E. C. McMonagle. 1983. Characterization of theTRs/IRs origin of DNAreplication of herpes simplex virus type 1. Virology 130:427-438.
31. Vaughan, P. J., D. J. M. Purifoy, and K. L. Powell. 1985. DNA-binding protein associated with herpes simplex virus DNApolymerase. J. Virol. 53:501-508.
32. Weller, S. K., K. J. Lee, D. J. Sabourin, and P. A. Schaffer. 1983. Geneticanalysis oftemperature-sensitive mutantswhich define the gene for the major herpes simplex virus type 1 DNA-binding protein. J. Virol. 45:354-366.