Ruimin Pan,aMiroslaw K. Gorny,bSusan Zolla-Pazner,b,cXiang-Peng Konga Departments of Biochemistry and Molecular Pharmacologya
and Pathology,b
NYU School of Medicine, New York, New York, USA; Veterans Affairs New York Harbor Healthcare System, New York, New York, USAc
ABSTRACT
The region consisting of the first and second variable regions (V1V2) of gp120 plays vital roles in the functioning of the HIV-1 envelope (Env). V1V2, which harbors multiple glycans and is highly sequence diverse, is located at the Env apex and stabilizes the trimeric gp120 spike on the virion surface. It shields V3 and the coreceptor binding sites in the prefusion state and exposes them upon CD4 binding. Data from the RV144 human HIV-1 vaccine trial suggested that antibody responses targeting the V1V2 region inversely correlated with the risk of infection; thus, understanding the antigenic structure of V1V2 can contribute to
vac-cine design. We have determined a crystal structure of a V1V2 scaffold molecule (V1V2ZM109-1FD6) in complex with 830A, a
hu-man monoclonal antibody that recognizes a V1V2 epitope overlapping the integrin-binding motif in V2. The structure revealed that V1V2 assumes a five-stranded beta barrel structure with the region of the integrin-binding site (amino acids [aa] 179 to 181) included in a “kink” followed by an extra beta strand. The complete barrel structure naturally presents the glycans on its outer surface and packs into its core conserved hydrophobic residues, including the Ile at position 181 which was highly correlated with vaccine efficacy in RV144. The epitope of monoclonal antibody 830A is discontinuous and composed of three segments: (i)
Thr175, Tyr177, Leu179, and Asp180at the kink overlapping the integrin-binding site; (ii) Arg153and Val154in V1; and (iii) Ile194at
the C terminus of V2. This report thus provides the atomic details of the immunogenic “V2i epitope.”
IMPORTANCE
Data from the RV144 phase III clinical trial suggested that the presence of antibodies to the first and second variable regions (V1V2) of gp120 was associated with the modest protection afforded by the vaccine. V1V2 is a highly variable and immunogenic region of HIV-1 surface glycoprotein gp120, and structural information about this region and its antigenic landscape will be cru-cial in the design of an effective HIV-1 vaccine. We have determined a crystal structure of V1V2 in complex with human MAb
830A and have shown that MAb 830A recognizes a region overlapping the␣47 integrin-binding site. We also showed that V1V2
forms a 5-stranded beta barrel, an elegant structure allowing sequence variations in the strand-connecting loops while preserv-ing a conserved core.
T
he HIV-1 envelope (Env) glycoprotein complex, consisting of glycoproteins gp120 and gp41, mediates the virus entry into the host cell. This heterotrimer is the major target for neutralizing antibodies (Abs) induced in HIV-1-infected patients and for HIV/ AIDS vaccine development (1). Glycoprotein gp120 has five vari-able and five conserved regions (2); the region consisting of the first and second variable regions (V1V2) is the most diverse in both sequence and length. V1V2, including residues 126 to 196 in the HXB2 numbering scheme (3), usually has two nested disulfide bonds; the V1 disulfide bond (between cysteine residues 131 and 157) is located within the V2 disulfide bond (between residues 126 and 196) (4). The average length of V1V2 is about 80 amino acids (aa), with a possibility of large length variations mostly derived from two regions, one in the middle of V1 and the other near the C-terminal end of V2. Recent data have shown that V1V2 is lo-cated at the distal apex of the Env trimer and that the V1V2 regions from the three gp120s, at least in the stabilized BG505 SOSIP.664 construct, join at the center to form a top layer of the trimer (5–7). This layer can shield the third variable region (V3) and the core-ceptor binding site in the prefusion state, and it can also make large movements upon CD4 binding to expose the coreceptor binding site.Due to the nested disulfide bond configuration, it was difficult to predict the structural conformation of V1V2 from sequence analysis and epitope mapping (8). V1V2 was excluded in the early attempts to crystalize monomeric forms of gp120 (9,10). The first
breakthrough in visualizing the V1V2 structure was achieved by Kwong and coworkers, who crystallized complexes of broadly neutralizing monoclonal antibodies (MAbs) PG9 and PG16 with the V1V2 inserted in a scaffold molecule (Protein Data Bank iden-tifier [PDB ID] 1FD6); two clade C V1V2s, CAP45 (referred to here as V1V2CAP45-1FD6) and ZM109 (V1V2ZM109-1FD6), were
used (11,12). The 1FD6 scaffold molecule was among many tried and is a small protein with a region that can accommodate the disulfide bond and glycans of V1V2 (11). These structures showed that V1V2 contains 4 beta strands (named strands A, B, C, and D) forming a Greek key motif, an elegant solution to allow a con-served motif with two sub-variable regions. However, regions of V1V2 were still missing from these structures. These include part of V1 in V1V2ZM109-1FD6 and the sub-variable C-terminal V2
Received25 March 2015Accepted16 May 2015
Accepted manuscript posted online27 May 2015
CitationPan R, Gorny MK, Zolla-Pazner S, Kong X-P. 2015. The V1V2 region of HIV-1 gp120 forms a five-stranded beta barrel. J Virol 89:8003–8010.
doi:10.1128/JVI.00754-15.
Editor:W. I. Sundquist
Address correspondence to Xiang-Peng Kong, [email protected].
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00754-15
on November 7, 2019 by guest
http://jvi.asm.org/
region in both structures. The missing V2 region contains a highly conserved integrin-binding motif,179Leu-Asp-Ile/Val181, which can bind the␣47 integrin, the gut mucosal homing receptor for peripheral T cells (13). Although the precise function of this motif is still being questioned (14), this region may play a potentially important role in facilitating the virion getting onto cells in order for it to get into cells, and targeting␣47 reduces mucosal trans-mission in an animal model (15).
The V1V2 region is immunogenic in humans (8, 16), and many V1V2-specific MAbs have been isolated from HIV-1-in-fected patients. We have recently classified V1V2 epitopes into three major types: V2q, V2p, and V2i (17,18). The V2q type is defined by quaternary neutralizing epitope (QNE) MAbs, includ-ing human MAbs 2909, PG9, and PG16, and also a panel of rhesus MAbs, including 2.5B (19–21). Crystal structures of PG9 and PG16 in complex with the V1V2 scaffolds have shown that these MAbs recognize an N-linked glycan at residue 160 and another at either 156 or 173 as well as the C-terminal region of strand B of V1V2 through a beta sheet interaction using long complementa-rity-determining-region (CDR) H3s harbored by these QNE MAbs (11,12). The V2p type is defined by human MAbs CH58 and CH59 isolated from a vaccinee of the phase III RV144 human vaccine trial (22). Their epitopes are also located in strand B, over-lapping the V2q epitope region, but have a helical or helical-coil structure, different from the beta strand structure of the V2q type (22). The V2i type is defined by a panel of seven human MAbs, including 830A, 697-30D, and 2158 (17,23). Extensive immuno-logical, mutagenesis, and computational modeling data have shown that the V2i epitopes overlap the integrin-binding site and likely recognize discontinuous regions in V1V2 (17,18,23). How-ever, the precise nature of the V2i epitopes is still unknown.
Data from the correlation analysis of the RV144 human clinical vaccine trial have suggested that the IgGs targeting V1V2 induced by the vaccine inversely correlate with the risk of infection (24), and immunologic data delineated the cross-reactivity of these Abs (25,26). A subsequent sequence sieve analysis that compared the V1V2 sequences from viruses infecting placebo and vaccine recip-ients identified two positions in V2, 169 and 181, that distin-guished viruses from vaccine recipients. The RV144 vaccine effi-cacy against viruses matching the Lys vaccine residue at position 169 was 48%, whereas the vaccine efficacy against viruses mis-matching the Ile vaccine residue at 181 was 78% (27). We re-cently modeled the full-length V1V2, including the V2i epitope region, and provided a mechanistic understanding of mis-matching sieve residue 181 (18). To further understand the structure and function of V1V2 and to precisely map the V2i epitopes, we determined the structure of the antigen-binding fragment (Fab) of V2i MAb 830A (an IgG3 MAb with VH4-34 heavy chain and VL kappa 3-15 light chain genes) in complex with a V1V2 scaffold (V1V2ZM109-1FD6). Our structure reveals
the atomic details of the 830A epitope and the integrin-binding site. In addition, our data showed that the V1V2 region forms a five-stranded beta barrel, a structure that is uniquely suitable for the functions of V1V2.
MATERIALS AND METHODS
Fab 830A production and complex formation. Fully glycosylated V1V2ZM109-1FD6 was produced by transient transfection using a mam-malian expression system as previously reported (11). Briefly, the V1V2 scaffold protein with all the glycan sites was produced in 293S GnTI⫺/⫺
cells that produce only high-mannose glycans, which are more homo-geneous than complex glycans and better suited to crystallization (11, 28). It was then purified by the use of nickel-nitrilotriacetic acid (Ni-NTA) beads. The DNAs of the heavy and light chains of Fab 830A were synthesized by Genewiz (South Plainfield, NJ) and subcloned into the pVRC-8400 expression vector with the secretion signal (MRP TWAWWLFLVLLLALWAPARG) at their N termini and a 6⫻His tag at the C terminus of the heavy chain. Fab 830A were expressed and also purified with the His tag affinity columns. Purified V1V2ZM109-1FD6 and Fab 830A were mixed at a 1:1 molar ratio and further purified by size exclusion chromatography.
Crystallization, data collection, and structure determination and refinement.Data representing the initial crystallization conditions of Fab 830A/V1V2ZM109-1FD6 were obtained by robotic screening using the va-por diffusion hanging drop method. Diffracting crystals of Fab/epitope complex were obtained with a well solution of 16.5% polyethylene glycol 8000 – 0.1 M Tris (pH 8.5). Many crystals were produced, but they rarely diffracted well. Final data were collected at the beamline General Medical Sciences and Cancer Institutes Structural Biology Facility Collaborative Access Team (GM/CA CAT) at the Advanced Photon Source (APS), Ar-gonne National Laboratory, and processed using the HKL 2000 package (29) and XDS (30). The structure was determined by molecular replace-ment using the structures of PG9-bound V1V2ZM109-1FD6 (PDB ID 3U2S) and a homologous Fab fragment (PDB ID 3KDM) as the starting models. Cycles of refinement for the structure were carried out in Coot and Phenix (31,32). Final structural analysis was carried out using ICM (33), and figures were generated using ICM and PyMOL (Schrödinger, LLC).
RESULTS
Structure determination of Fab 830A/V1V2zm109-1FD6
com-plex.To crystallize human MAb 830A in complex with V1V2, we
cloned its Fab fragment and expressed it in 293 cells, as it was difficult to obtain a large amount of IgG from the hybridoma cells. Fully glycosylated (with all 6 glycosylation sites) V1V2zm109-1FD6
was also produced in 293 cells, and the complex of Fab 830A and
V1V2zm109-1FD6 was obtained by mixing them at a 1:1 molar
ratio and further purifying by size exclusion chromatography. The crystals of the complex diffracted to better than 3-Å resolution and belonged to the orthorhombic space group P212121with two Fab/
epitope complexes in the asymmetric unit. We determined the structure by molecular replacement and refined the model to 3.0-Å resolution with a finalRworkvalue of 22% (Rfree⫽29.5%)
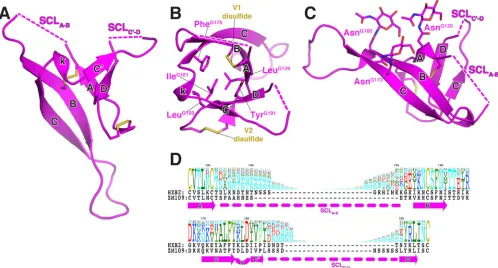
(Table 1andFig. 1). Since the two noncrystallographic complexes in the crystal are highly similar (Fig. 1AandC), we chose only one for description. We numbered the residues of the light and heavy chains following the Kabat and Wu convention, preceded by “L” and “H,” respectively, and the residues of V1V2ZM109-1FD6 by a
“G” (for “gp120”).
Structure of V1V2ZM109.The V1V2 structure we observed in
the 830A complex still has the four previously described beta strands in the PG9 complex, i.e., strand A (from ThrG128 to
AsnG130), strand B (from ArgG153 to AsnG160), strand C (from
LysG171to TyrG177), and strand D (from LeuG190to LeuG193) (Fig.
1C). Each strand in the 830A complex structure is slightly shorter than those observed in the PG9 complex (Fig. 1D). However, we have observed an additional beta strand from IleG181to LeuG184
located between strand C and D and we have named it strand C=
(Fig. 1Cand2A). Together with strand C=, these five beta strands now form a complete beta barrel structure (Fig. 2). The shorter lengths of the beta strands allow them to curve around the barrel so that it has a cylindrical shape. The V1V2 beta barrel has several
on November 7, 2019 by guest
http://jvi.asm.org/
characteristics (Fig. 2). V1V2 of ZM109 has six N-linked glycosy-lation sites, and we observed (for only singleN-acetylglucosamine [NAG] moieties) three of them on AsnG130, AsnG160, and AsnG173,
all extending out from the barrel toward one side (Fig. 2C). The remaining glycosylation sites, AsnG138, AsnG188a, and AsnG188d, are
in the flexible strand-connecting loops (SCLs) (Fig. 2), which are partly missing in the structure. Combining the five-stranded beta barrel structure with the sequence variation of V1V2, we can now redefine the highly variable regions in V1V2 more precisely: the V1 variable region is the strand-connecting loop between strand A and B (SCLA-B), while the V2 variable region is the
strand-con-necting loop between strand C=and D (SCLC=-D) (Fig. 2D). These
two strand-connecting loops have high sequence diversity and a high percentage of N-linked glycosylation sites among the differ-ent HIV-1 strains (Fig. 2D).
Hydrophobic core.The V1V2 beta barrel has a hydrophobic
core formed by a number of hydrophobic residues (Fig. 2B), in-cluding ValG127and LeuG129from strand A, the disulfide bond of
V1 between CysG131and CysG157, PheG159and IleG161near the end
of strand B, ValG172, AlaG174, and PheG176of strand C, IleG181of
strand C=, and TyrG191and LeuG193of strand D. The side chain of RV144 sieve residue IleG181on strand C=is buried in the core, with a buried surface area of⬃80 Å2. A mutation of 181 to Val, a
mismatching amino acid in the RV144 sieve analysis (27), would have a reduced buried surface area of 67 Å2. On the other hand, a
Met or a Leu, also mismatching amino acids in the RV144 sieve analysis, at this position would be slightly too bulky to pack into the hydrophobic core. Thus, the Ile amino acid seems ideal for position 181 of the hydrophobic core of the beta barrel.
The integrin-binding site.The sequence of the ZM109␣47
integrin-binding motif is LeuG179-AspG180-IleG181. The first two
residues, LeuG179 and AspG180, are located in a short 3
10helical
[image:3.585.39.287.76.292.2]turn between strands C and C=(named “kink”), while IleG181is located on strand C=(Fig. 2BandD). In contrast to the buried
TABLE 1Crystallization and refinement statisticsa
Parameter 830A/V1V2ZM109result(s)
Data collection
Space group P212121
Cell dimensions
a, b, c (Å) 64.73, 159.17, 170.36 ␣,,␥(°) 90, 90, 90
Resolution (Å) 3.00 (3.18–3.00)
CC (1/2) 99.9 (79.4)
Rmeas 5.8 (83.3)
I/I 19.16 (2.26)
Completeness (%) 99.6 (99.5)
Redundancy 4.5 (4.5)
Refinement
Resolution (Å) 49.03–3.00
No. of reflections 35,968 Rwork/Rfree 22.0/29.5
RMS deviations
Bond length (Å) 0.010
Bond angle (°) 1.422
aStatistics in parentheses refer to the outer resolution shell. CC (1/2), percentage of
correlation between intensities of random half data sets; RMS, root mean square.
FIG 1Structure of Fab 830A in complex with V1V2ZM109-1FD6. (A) Molecules in the asymmetric unit (ASU) represented as ribbons. There are two Fab
830A/V1V2ZM109-1FD6 complexes in the ASU: the two V1V2ZM109-1FD6 complexes are colored magenta and salmon, respectively, while the heavy chains of Fab
830A are colored green and light blue and the light chains are colored cyan and wheat, respectively. (B) One of the Fab 830A/V1V2ZM109-1FD6 complexes in a
side view. Only the Fv region of 830A is shown, and the 1FD6 scaffold is colored gray. (C) Superimposition of the two V1V2s in the ASU. The five strands (A, B, C, C=, and D) and the short 310helical turn (k, “kink”) are labeled. Note that the strands connecting the loop regions have slightly different conformations in the
two molecules. (D) Superimposition of 830A-bound V1V2 (magenta) and PG9-bound V1V2 (gray). Note the extra C=strand in 830A-bound V1V2.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.138.450.417.666.2]IleG181, the highly conserved LeuG179 and the 100% conserved AspG180are exposed on the surface of the kink region (Fig. 3A).
But attempts to computationally dock the ZM109 integrin-bind-ing motif onto the␣47 head structure were not successful; the integrin-binding site in our structure is too recessed to allow reaching the deep binding site shown in the␣47 head structure (34).
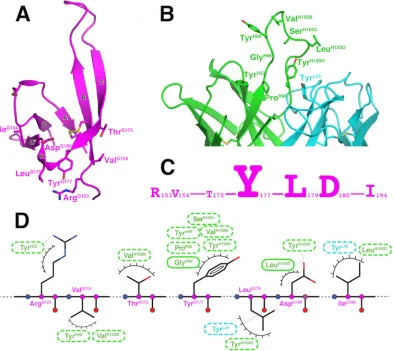
830A-V1V2ZM109interactions.Our structure revealed the
de-tails of the epitope of 830A (Fig. 3). It comprised three compo-nents: (i) residues ArgG153 and ValG154 from V1, (ii) residues
ThrG175and TyrG177from strand C and LeuG179and AspG180from the kink region, and (iii) IleG194from the V2 stem just after the
C-terminal strand D (Fig. 3A). The side chain of ArgG153in the first component forms a hydrogen bond with the hydroxyl group of TyrH32from CDR H1 of 830A, and ValG154forms minor con-tacts against two residues, TryH99and ValH100B, at the tip of CDR
H3 (Fig. 3D). The most prominent interaction between 830A and V1V2 is in the second component, including TyrG177, the last
res-idue of strand C, and LeuG179and AspG180, the first two residues of the ␣47 integrin-binding motif in the kink region. The side chains of TyrG177and LeuG179pack against a wall formed by the backbone of GlyH98and the side chains of TyrH100Hand TyrL49
from the antibody side. These three residues have a total contact surface area of 147 Å2with 830A, which is more than 70% of the
total buried surface area on the V1V2 side (Fig. 3C). In addition, the side chain of AspG180can form hydrogen bonds with the
back-bone nitrogen of IleH100Dand the hydroxyl group of TyrH100H
from the CDR H3 of 830A, and the carbonyl oxygen of LeuG179 forms a hydrogen bond with the hydroxyl group of TyrL49from
CDR L1. Similarly to residues in the first component of the epitope, IleG194in the third component, located in the stem of V2,
forms van der Waals contacts with hydrophobic residue IleH100D of CDR H3. We did not observe any contacts between 830A and glycans of V1V2ZM109, consistent with enzyme-linked
immu-nosorbent assay (ELISA) data showing that deglycosylation of
V1V2ZM109-1FD6 did not affect 830A binding (data not shown).
The CDR H3 of 830A is 18 amino acids long (Kabat definition), and it stands tall in the center of the antigen-binding site, protrud-ing about 10 Å from its base (Fig. 3B). This is different from two other V2i MAbs, 697 and 2158, whose CDR H3s are collapsed, making their antigen-binding sites flat (18,23). The CDR H3 of 830A contributes the majority of its antigen binding, and all resi-dues of the 830A paratope are from the heavy chain except TyrL49
(Fig. 3B).
DISCUSSION
Here we have defined the epitope of one of the human V2i MAbs, 830A, at an atomic resolution, and the epitope of 830A overlaps the␣47 integrin-binding motif. It consists of multiple structural components, including regions from V1, the kink, and the C ter-minus of V2 (Fig. 3A,C, andDand4A). Data from one of our previous mutagenesis studies suggested that mutations of residues in strand B of V1V2, including residues 168 and 169, can also influence 830A binding (17), but we did not observe direct
con-FIG 2Five-stranded V1V2ZM109beta barrel. (A) The five-stranded beta barrel structure of V1V2. Regions missing in electron density in the two
strand-connecting loops (SCLs) between strands A and B (SCLA-B) and between strands C=and D (SCLC=-D) are represented by dashed lines and found at one end of the
barrel. (B) A top view of the hydrophobic core. For clarity, only side chains of selected hydrophobic residues are shown. (C) A side view of the V1V2 barrel. Note the three observed glycans pointing toward one side of the barrel. (D) Alignment of V1V2 sequences. The amino acid profile of the 2013 premade envelope alignment of the LANL database sequences (http://www.hiv.lanl.gov) is shown. The size of each amino acid in the profile is drawn proportionally to its weight of conservation at the particular location. The V1V2 sequences of HXB2 and ZM109 are placed below the profile. The regions of beta strands observed in the crystal structure and the strand-connecting loops (SCLA-Band SCLC=-D) are illustrated by arrows and dashed lines, respectively.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.43.541.69.337.2]tacts between these residues and 830A in the crystal structure (Fig. 3). It is therefore possible that mutations of those residues can alter the global conformation of V1V2, thus affecting the confor-mation of the 830A epitope. The multicomponent epitope of 830A is likely conformationally flexible, as the kink and strand C=
region were not observed in several previously published crystal structures (5,11,12). This conformational flexibility of the V2i epitopes provides a possible explanation for why the neutralizing activity of 830A and other V2i MAbs is substantially improved with prolonged incubation (unpublished data and reference35). Although the seven human V2i MAbs (830A, 2297, 697, 1361, 1393A, 1357, and 2158) have been described as a family (17,18), their gene usages differ. Monoclonal antibodies 830A and 2297 are encoded by VH4, while the other five are encoded by VH1 (23). In particular, four of these MAbs (697, 1361, 1393A, and 2158) are encoded by VH1-69, and we have determined the Fab structures for two of them, 697 and 2158, and have shown that they have a hydrophobic surface patch in their antigen-binding sites (18,23). This is not observed in the case of 830A. Functionally, VH4 MAbs 830A and 2297 have a narrower neutralization breadth even
though 830A has at least 5% more somatic mutations than the other V2i MAbs (23). It is therefore conceivable that the seven V2i MAbs can be divided into subgroups, each with a distinct V1V2 binding mode, which can be elucidated only by future structural studies.
Stabilization of the V2 kink region by Fab 830A binding likely helped the visualization of the strand C=region and the beta barrel structure of V1V2, although the full glycosylation of the construct we used may also have helped to stabilize the beta barrel structure. In the previously determined PG9- and PG16-bound V1V2ZM109
-1FD6 structures (11,12), the Fab molecules bound the N-terminal region of strand C, which is at the other end of the beta barrel from the kink region. Thus, the kink/strand C=was flexible in those complexes, preventing their visualization in the crystal structures (11,12). Overall, regions of V1V2, such as the connecting loops, are likely quite flexible and can adopt various conformations. However, the five-stranded beta barrel core structure of V1V2 represents a more complete and comprehensive solution for the structure and function of V1V2 than the four-stranded Greek key motif (11). The cylindrical shape consists of a hydrophobic core
FIG 3Details of the antigen-antibody interactions. (A) Key residues of V1V2ZM109recognized by Fab 830A. The side chains of residues with areas of contact with
830A that are more than 10 Å2in size are shown. Note that two of the three integrin-binding site residues, LeuG179and AspG180, are exposed and are part of the
epitope of 830A. (B) Key residues of the 830A paratope. (C) Amino acids of the 830A epitope, with the size of each residue drawn proportionally to its area of contact with the antibody. (D) Schematic diagram of the antigen-antibody interaction. Hydrogen-binding interactions are indicated by dashed lines between the residues, while van der Waals contacts are represented by “eyelashes.” Residues in solid ovals contribute to the interactions by their main chain atoms, and those in dashed ovals contribute to the interactions by their side chain atoms. Residues in cyan and green are from the 830A light and heavy chains, respectively.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.97.491.69.420.2]with side chains of charged/polar residues projecting out from the barrel surface, and glycosylation sites decorating the outer surface. The two extreme sequence- and length-variable regions of V1 and V2 (SCLA-Band SCLC=-D) are precisely defined, and both point to
one end of the barrel (Fig. 2). This configuration allows length variations for these SCLs (Fig. 2D) without affecting the main barrel structure. Interestingly, data have suggested that immune pressure can drive the length of these regions, which may serve as a tool for the virus to evade the immune response (36–38). How-ever, these SCLs are located at the periphery of the trimer and it is not yet known if they can protect any key sites of Env vulnerability. In view of the high sequence conservation of the kink/strand C=
(residues 175 to 184;Fig. 2D), one may speculate that at least one of the functions of these SCLs is to protect this conserved region of V2. Thus, increasing the lengths of SCLC=-D would increase the
flexibility of strand C=, leading to conformationally masking the integrin-binding site and strand C=(39).
A beta barrel structure also allows V1V2 to function as an in-dividual module. V1V2 is located on the distal apex of the trimer, packed on top of V3, but it has to make a large movement upon CD4 binding to expose V3 and the coreceptor binding site. As individual functional modules, V1V2 and V3 can pack against each other loosely but can also move away independently from the gp120 core when CD4 binds. Such functional modules are com-mon in signal transduction protein molecules (40). During prep-aration of the manuscript, a 3.5-Å resolution structure of the tri-meric form of envelope BG505 SOSIP.664 was published (7), and its V1V2 region has essentially the same overall structure as we observed here; thus, the V1V2 structure that we resolved and de-scribe in the context of an antibody-bound V1V2 scaffold likely represents V1V2 in its native conformation. We noticed that the structures of the strand C region observed in the CH58 and CH59 complexes are very different from that of the scaffolded V1V2s or the SOSIP. The CH58/CH59 conformations likely present in free gp120 molecules, which were used for the RV144 vaccine.
The antigenic sites on V1V2 have become crucial targets for vaccine design since the correlate study and sieve analysis of the RV144 clinical vaccine trial highlighted the role of antibodies to this region in reducing the risk of HIV infection (24–27). The structural data presented here provide additional support for the important role of these antibodies. The central component of the 830A epitope, the kink region overlapping with the integrin-binding site, is highly conserved in V1V2 (41,42). This immuno-genic region is likely another vulnerable site in V1V2, in addition to the strand C targeted by PG9 and other QNE MAbs. Precise structural mapping of this epitope region provides an opportunity to rationally design scaffold immunogens to focus antibody re-sponses on this region (43,44). One challenge of targeting this epitope region is the flexibility of strand C=, which may render this region less immunogenic than the strand C region. However, strand C=can be stabilized by an additional disulfide bond as in the case of HIV-2 and some simian immunodeficiency virus (SIV) envelopes (45). In fact, mutations to introduce such a disulfide bond into certain virus strains improved the neutralization po-tency of 830A (C. Hioe, personal communications). Thus, stabi-lizing this region may improve the antigenicity and immunoge-nicity of this conserved region in V1V2. Taking the results together, precise epitope mapping of V1V2-specific MAbs such as 830A will improve our ability to design immunogens targeting key antigenic epitopes in the V1V2 region of HIV-1 gp120.
Protein structure accession number.Atomic coordinates and
structure factors for the Fab 830A/V1V2ZM109-1FD6 complex
have been deposited in the RCSB Protein Data Bank under ID code 4YWG.
ACKNOWLEDGMENTS
We thank Peter Kwong for the construct of 1FD6-V1V2ZM109and Brian Foley for help with downloading the Env sequence alignment from the LANL database.
This work was supported in part by the National Institutes of Health under awards AI100151, AI082274, AI084119, and HL059725 and by funds for the Department of Veterans Affairs. GM/CA-CAT at APS has
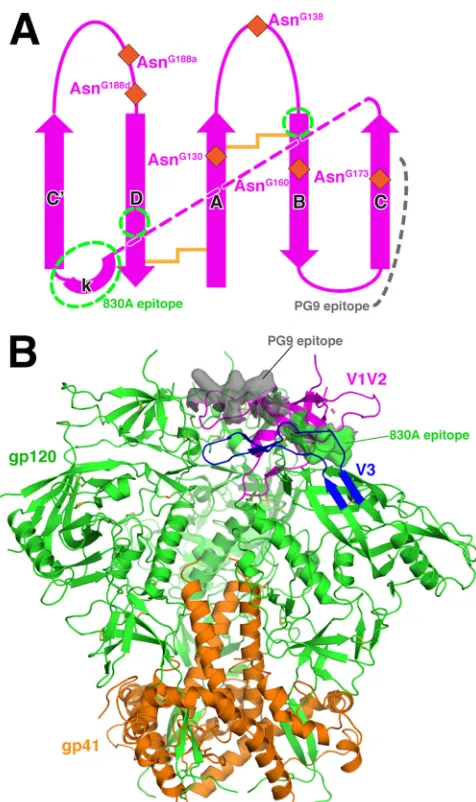
FIG 4Topology of V1V2 and location of 830A epitope in the trimer context. (A) The topology of V1V2ZM109. Regions of the 830A epitope are indicated by
green dashed circles. The locations of the six glycosylation sites in V1V2ZM109
(only three were observed in the 830A complex) are indicated by brown dia-monds. The pink dashed line depicts the connection of strand C with the kink. (B) The location of the 830A epitope in the context of the BG505 SOSIP.664 prefusion trimer (PDB ID 4TVP). The trimeric complex of gp120 (green) and gp41 (orange) is viewed from the side, and regions V1V2 and V3 of one of the gp120s are colored magenta and blue, respectively. The 830A epitope is shown as a green surface, while the PG9 epitope is also shown as a gray surface (for simplicity, the glycans are not displayed).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.45.283.63.464.2]been funded in whole or in part with federal funds from the National Cancer Institute (Y1-CO-1020) and the National Institute of General Medical Sciences (Y1-GM-1104). Use of the Advanced Photon Source was supported by the U.S. Department of Energy, Basic Energy Sciences, Of-fice of Science, under contract DE-AC02-06CH11357.
The content is solely our responsibility and does not necessarily rep-resent the official views of the National Institutes of Health or the Depart-ment of Veterans Affairs.
REFERENCES
1.Wyatt R, Sodroski J.1998. The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens. Science280:1884 –1888.http://dx.doi.org/10 .1126/science.280.5371.1884.
2.Modrow S, Hahn BH, Shaw GM, Gallo RC, Wong-Staal F, Wolf H.
1987. Computer-assisted analysis of envelope protein sequences of seven human immunodeficiency virus isolates: prediction of antigenic epitopes in conserved and variable regions. J Virol61:570 –578.
3.Ratner L, Fisher A, Jagodzinski LL, Mitsuya H, Liou RS, Gallo RC, Wong-Staal F.1987. Complete nucleotide sequences of functional clones of the AIDS virus. AIDS Res Hum Retroviruses3:57– 69.http://dx.doi.org /10.1089/aid.1987.3.57.
4.Leonard CK, Spellman MW, Riddle L, Harris RJ, Thomas JN, Gregory TJ.1990. Assignment of intrachain disulfide bonds and characterization of potential glycosylation sites of the type 1 recombinant human immu-nodeficiency virus envelope glycoprotein (gp120) expressed in Chinese hamster ovary cells. J Biol Chem265:10373–10382.
5.Julien JP, Cupo A, Sok D, Stanfield RL, Lyumkis D, Deller MC, Klasse PJ, Burton DR, Sanders RW, Moore JP, Ward AB, Wilson IA.2013. Crystal structure of a soluble cleaved HIV-1 envelope trimer. Science342:
1477–1483.http://dx.doi.org/10.1126/science.1245625.
6.Lyumkis D, Julien JP, de Val N, Cupo A, Potter CS, Klasse PJ, Burton DR, Sanders RW, Moore JP, Carragher B, Wilson IA, Ward AB.2013. Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Sci-ence342:1484 –1490.http://dx.doi.org/10.1126/science.1245627.
7.Pancera M, Zhou T, Druz A, Georgiev IS, Soto C, Gorman J, Huang J, Acharya P, Chuang GY, Ofek G, Stewart-Jones GB, Stuckey J, Bailer RT, Joyce MG, Louder MK, Tumba N, Yang Y, Zhang B, Cohen MS, Haynes BF, Mascola JR, Morris L, Munro JB, Blanchard SC, Mothes W, Connors M, Kwong PD.2014. Structure and immune recognition of trimeric pre-fusion HIV-1 Env. Nature514:455– 461.http://dx.doi.org/10 .1038/nature13808.
8.Moore JP, Sattentau QJ, Yoshiyama H, Thali M, Charles M, Sullivan N, Poon SW, Fung MS, Traincard F, Pinkus M, Robey G, Robinson JE, Ho DD, Sodroski J.1993. Probing the structure of the V2 domain of human immunodeficiency virus type 1 surface glycoprotein gp120 with a panel of eight monoclonal antibodies: human immune response to the V1 and V2 domains. J Virol67:6136 – 6151.
9.Kwong PD, Wyatt R, Robinson J, Sweet RW, Sodroski J, Hendrickson WA.1998. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature393:
648 – 659.http://dx.doi.org/10.1038/31405.
10. Huang CC, Tang M, Zhang MY, Majeed S, Montabana E, Stanfield RL, Dimitrov DS, Korber B, Sodroski J, Wilson IA, Wyatt R, Kwong PD.
2005. Structure of a V3-containing HIV-1 gp120 core. Science310:1025– 1028.http://dx.doi.org/10.1126/science.1118398.
11. McLellan JS, Pancera M, Carrico C, Gorman J, Julien JP, Khayat R, Louder R, Pejchal R, Sastry M, Dai K, O’Dell S, Patel N, Shahzad-ul-Hussan S, Yang Y, Zhang B, Zhou T, Zhu J, Boyington JC, Chuang GY, Diwanji D, Georgiev I, Kwon YD, Lee D, Louder MK, Moquin S, Schmidt SD, Yang ZY, Bonsignori M, Crump JA, Kapiga SH, Sam NE, Haynes BF, Burton DR, Koff WC, Walker LM, Phogat S, Wyatt R, Orwenyo J, Wang LX, Arthos J, Bewley CA, Mascola JR, Nabel GJ, Schief WR, Ward AB, Wilson IA, Kwong PD. 2011. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature480:336 –343.http://dx.doi.org/10.1038/nature10696. 12. Pancera M, Shahzad-ul-Hussan S, Doria-Rose NA, McLellan JS, Bailer
RT, Dai K, Loesgen S, Louder MK, Staupe RP, Yang Y, Zhang B, Parks R, Eudailey J, Lloyd KE, Blinn J, Alam SM, Haynes BF, Amin MN, Wang LX, Burton DR, Koff WC, Nabel GJ, Mascola JR, Bewley CA, Kwong PD.2013. Structural basis for diverse N-glycan recognition by HIV-1-neutralizing V1-V2-directed antibody PG16. Nat Struct Mol Biol
20:804 – 813.http://dx.doi.org/10.1038/nsmb.2600.
13. Arthos J, Cicala C, Martinelli E, Macleod K, Van Ryk D, Wei D, Xiao Z, Veenstra TD, Conrad TP, Lempicki RA, McLaughlin S, Pascuccio M, Gopaul R, McNally J, Cruz CC, Censoplano N, Chung E, Reitano KN, Kottilil S, Goode DJ, Fauci AS.2008. HIV-1 envelope protein binds to and signals through integrin alpha4beta7, the gut mucosal homing recep-tor for peripheral T cells. Nat Immunol9:301–309.http://dx.doi.org/10 .1038/ni1566.
14. Parrish NF, Wilen CB, Banks LB, Iyer SS, Pfaff JM, Salazar-Gonzalez JF, Salazar MG, Decker JM, Parrish EH, Berg A, Hopper J, Hora B, Kumar A, Mahlokozera T, Yuan S, Coleman C, Vermeulen M, Ding H, Ochsenbauer C, Tilton JC, Permar SR, Kappes JC, Betts MR, Busch MP, Gao F, Montefiori D, Haynes BF, Shaw GM, Hahn BH, Doms RW.
2012. Transmitted/founder and chronic subtype C HIV-1 use CD4 and CCR5 receptors with equal efficiency and are not inhibited by blocking the integrin alpha4beta7. PLoS Pathog8:e1002686.http://dx.doi.org/10.1371 /journal.ppat.1002686.
15. Byrareddy SN, Kallam B, Arthos J, Cicala C, Nawaz F, Hiatt J, Kersh EN, McNicholl JM, Hanson D, Reimann KA, Brameier M, Walter L, Rogers K, Mayne AE, Dunbar P, Villinger T, Little D, Parslow TG, Santangelo PJ, Villinger F, Fauci AS, Ansari AA. 2014. Targeting alpha4beta7 integrin reduces mucosal transmission of simian immunode-ficiency virus and protects gut-associated lymphoid tissue from infection. Nat Med20:1397–1400.http://dx.doi.org/10.1038/nm.3715.
16. Gorny MK, Moore JP, Conley AJ, Karwowska S, Sodroski J, Williams C, Burda S, Boots LJ, Zolla-Pazner S.1994. Human anti-V2 monoclonal antibody that neutralizes primary but not laboratory isolates of human immunodeficiency virus type 1. J Virol68:8312– 8320.
17. Mayr LM, Cohen S, Spurrier B, Kong XP, Zolla-Pazner S.2013. Epitope mapping of conformational V2-specific anti-HIV human monoclonal an-tibodies reveals an immunodominant site in V2. PLoS One8:e70859.http: //dx.doi.org/10.1371/journal.pone.0070859.
18. Spurrier B, Sampson J, Gorny MK, Zolla-Pazner S, Kong XP.2014. Functional implications of the binding mode of a human conformation-dependent V2 monoclonal antibody against HIV. J Virol88:4100 – 4112. http://dx.doi.org/10.1128/JVI.03153-13.
19. Gorny MK, Stamatatos L, Volsky B, Revesz K, Williams C, Wang XH, Cohen S, Staudinger R, Zolla-Pazner S.2005. Identification of a new quaternary neutralizing epitope on human immunodeficiency virus type 1 virus particles. J Virol79:5232–5237.http://dx.doi.org/10.1128/JVI.79.8 .5232-5237.2005.
20. Robinson JE, Franco K, Elliott DH, Maher MJ, Reyna A, Montefiori DC, Zolla-Pazner S, Gorny MK, Kraft Z, Stamatatos L.2010. Quater-nary epitope specificities of anti-HIV-1 neutralizing antibodies generated in rhesus macaques infected by the simian/human immunodeficiency vi-rus SHIVSF162P4. J Virol84:3443–3453.http://dx.doi.org/10.1128/JVI .02617-09.
21. Walker LM, Phogat SK, Chan-Hui PY, Wagner D, Phung P, Goss JL, Wrin T, Simek MD, Fling S, Mitcham JL, Lehrman JK, Priddy FH, Olsen OA, Frey SM, Hammond PW, Kaminsky S, Zamb T, Moyle M, Koff WC, Poignard P, Burton DR.2009. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Sci-ence326:285–289.http://dx.doi.org/10.1126/science.1178746.
22. Liao HX, Bonsignori M, Alam SM, McLellan JS, Tomaras GD, Moody MA, Kozink DM, Hwang KK, Chen X, Tsao CY, Liu P, Lu X, Parks RJ, Montefiori DC, Ferrari G, Pollara J, Rao M, Peachman KK, Santra S, Letvin NL, Karasavvas N, Yang ZY, Dai K, Pancera M, Gorman J, Wiehe K, Nicely NI, Rerks-Ngarm S, Nitayaphan S, Kaewkungwal J, Pitisuttithum P, Tartaglia J, Sinangil F, Kim JH, Michael NL, Kepler TB, Kwong PD, Mascola JR, Nabel GJ, Pinter A, Zolla-Pazner S, Haynes BF.2013. Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity38:176 – 186.http://dx.doi.org/10.1016/j.immuni.2012.11.011.
23. Gorny MK, Pan R, Williams C, Wang XH, Volsky B, O’Neal T, Spurrier B, Sampson JM, Li L, Seaman MS, Kong XP, Zolla-Pazner S.7 March 2012, posting date. Functional and immunochemical cross-reactivity of V2-specific monoclonal antibodies from HIV-1-infected individuals. Vi-rologyhttp://dx.doi.org/10.1016/j.virol.2012.02.003.
24. Haynes BF, Gilbert PB, McElrath MJ, Zolla-Pazner S, Tomaras GD, Alam SM, Evans DT, Montefiori DC, Karnasuta C, Sutthent R, Liao HX, DeVico AL, Lewis GK, Williams C, Pinter A, Fong Y, Janes H, DeCamp A, Huang Y, Rao M, Billings E, Karasavvas N, Robb ML, Ngauy V, de Souza MS, Paris R, Ferrari G, Bailer RT, Soderberg KA,
on November 7, 2019 by guest
http://jvi.asm.org/
Andrews C, Berman PW, Frahm N, De Rosa SC, Alpert MD, Yates NL, Shen X, Koup RA, Pitisuttithum P, Kaewkungwal J, Nitayaphan S, Rerks-Ngarm S, Michael NL, Kim JH.2012. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N Engl J Med366:1275–1286.http://dx .doi.org/10.1056/NEJMoa1113425.
25. Zolla-Pazner S, deCamp A, Gilbert PB, Williams C, Yates NL, Williams WT, Howington R, Fong Y, Morris DE, Soderberg KA, Irene C, Reich-man C, Pinter A, Parks R, Pitisuttithum P, Kaewkungwal J, Rerks-Ngarm S, Nitayaphan S, Andrews C, O’Connell RJ, Yang ZY, Nabel GJ, Kim JH, Michael NL, Montefiori DC, Liao HX, Haynes BF, Tomaras GD.2014. Vaccine-induced IgG antibodies to V1V2 regions of multiple HIV-1 subtypes correlate with decreased risk of HIV-1 infection. PLoS One9:e87572.http://dx.doi.org/10.1371/journal.pone.0087572. 26. Zolla-Pazner S, deCamp AC, Cardozo T, Karasavvas N, Gottardo R,
Williams C, Morris DE, Tomaras G, Rao M, Billings E, Berman P, Shen X, Andrews C, O’Connell RJ, Ngauy V, Nitayaphan S, de Souza M, Korber B, Koup R, Bailer RT, Mascola JR, Pinter A, Montefiori D, Haynes BF, Robb ML, Rerks-Ngarm S, Michael NL, Gilbert PB, Kim JH.2013. Analysis of V2 antibody responses induced in vaccinees in the ALVAC/AIDSVAX HIV-1 vaccine efficacy trial. PLoS One8:e53629.http: //dx.doi.org/10.1371/journal.pone.0053629.
27. Rolland M, Edlefsen PT, Larsen BB, Tovanabutra S, Sanders-Buell E, Hertz T, deCamp AC, Carrico C, Menis S, Magaret CA, Ahmed H, Juraska M, Chen L, Konopa P, Nariya S, Stoddard JN, Wong K, Zhao H, Deng W, Maust BS, Bose M, Howell S, Bates A, Lazzaro M, O’Sullivan A, Lei E, Bradfield A, Ibitamuno G, Assawadarachai V, O’Connell RJ, deSouza MS, Nitayaphan S, Rerks-Ngarm S, Robb ML, McLellan JS, Georgiev I, Kwong PD, Carlson JM, Michael NL, Schief WR, Gilbert PB, Mullins JI, Kim JH.2012. Increased HIV-1 vaccine efficacy against viruses with genetic signatures in Env V2. Nature490:417– 420.http://dx.doi.org/10.1038/nature11519.
28. Reeves PJ, Callewaert N, Contreras R, Khorana HG.2002. Structure and function in rhodopsin: high-level expression of rhodopsin with restricted and homogeneous glycosylation by a tetracycline-inducible N-acetylglucosaminyltransferase I-negative HEK293S stable mammalian cell line. Proc Natl Acad Sci U S A99:13419 –13424.http://dx.doi.org/10.1073 /pnas.212519299.
29. Otwinowski Z, Minor W.1997. Processing of X-ray diffraction data collected in oscillation mode, p 307–326.InCarter CW, Sweet R (ed), Macromolecular crystallography, part A, vol 276. Academic Press, New York, NY.
30. Kabsch W.2010. Xds. Acta Crystallogr D Biol Crystallogr66:125–132. http://dx.doi.org/10.1107/S0907444909047337.
31. Emsley P, Cowtan K.2004. Coot: model-building tools for molecular graphics. Acta Crystallogr D Biol Crystallogr60:2126 –2132.http://dx.doi .org/10.1107/S0907444904019158.
32. Adams PD, Grosse-Kunstleve RW, Hung LW, Ioerger TR, McCoy AJ, Moriarty NW, Read RJ, Sacchettini JC, Sauter NK, Terwilliger TC.
2002. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr D Biol Crystallogr58:1948 – 1954.http://dx.doi.org/10.1107/S0907444902016657.
33. Abagyan R, Totrov M, Kuznetsov D.1994. ICM—a new method for protein modeling and design: applications to docking and structure
pre-diction from the distorted native conformation. J Comput Chem15:488 – 506.http://dx.doi.org/10.1002/jcc.540150503.
34. Yu Y, Zhu J, Mi LZ, Walz T, Sun H, Chen J, Springer TA. 2012. Structural specializations of alpha(4)beta(7), an integrin that mediates rolling adhesion. J Cell Biol196:131–146.http://dx.doi.org/10.1083/jcb .201110023.
35. Upadhyay C, Mayr LM, Zhang J, Kumar R, Gorny MK, Nadas A, Zolla-Pazner S, Hioe CE.2014. Distinct mechanisms regulate exposure of neutralizing epitopes in the V2 and V3 loops of HIV-1 envelope. J Virol
88:12853–12865.http://dx.doi.org/10.1128/JVI.02125-14.
36. Curlin ME, Zioni R, Hawes SE, Liu Y, Deng W, Gottlieb GS, Zhu T, Mullins JI.2010. HIV-1 envelope subregion length variation during dis-ease progression. PLoS Pathog 6:e1001228. http://dx.doi.org/10.1371 /journal.ppat.1001228.
37. van Gils MJ, Bunnik EM, Boeser-Nunnink BD, Burger JA, Terlouw-Klein M, Verwer N, Schuitemaker H.2011. Longer V1V2 region with increased number of potential N-linked glycosylation sites in the HIV-1 envelope glycoprotein protects against HIV-specific neutralizing antibod-ies. J Virol85:6986 – 6995.http://dx.doi.org/10.1128/JVI.00268-11. 38. Evering TH, Kamau E, St Bernard L, Farmer CB, Kong XP, Markowitz
M.2014. Single genome analysis reveals genetic characteristics of neuro-adaptation across HIV-1 envelope. Retrovirology11:65.http://dx.doi.org /10.1186/s12977-014-0065-0.
39. Kwong PD, Doyle ML, Casper DJ, Cicala C, Leavitt SA, Majeed S, Steen-beke TD, Venturi M, Chaiken I, Fung M, Katinger H, Parren PW, Rob-inson J, Van Ryk D, Wang L, Burton DR, Freire E, Wyatt R, Sodroski J, Hendrickson WA, Arthos J.2002. HIV-1 evades antibody-mediated neutral-ization through conformational masking of receptor-binding sites. Nature
420:678 – 682.http://dx.doi.org/10.1038/nature01188.
40. Pawson T.1995. Protein modules and signalling networks. Nature373:
573–580.http://dx.doi.org/10.1038/373573a0.
41. Zolla-Pazner S, Cardozo T.2010. Structure-function relationships of HIV-1 envelope sequence-variable regions refocus vaccine design. Nat Rev Immunol10:527–535.http://dx.doi.org/10.1038/nri2801.
42. Totrov M.2014. Estimated secondary structure propensities within V1/V2 region of HIV gp120 are an important global antibody neutraliza-tion sensitivity determinant. PLoS One9:e94002.http://dx.doi.org/10 .1371/journal.pone.0094002.
43. Totrov M, Jiang X, Kong XP, Cohen S, Krachmarov C, Salomon A, Williams C, Seaman MS, Abagyan R, Cardozo T, Gorny MK, Wang S, Lu S, Pinter A, Zolla-Pazner S.21 July 2010, posting date. Structure-guided design and immunological characterization of immunogens pre-senting the HIV-1 gp120 V3 loop on a CTB scaffold. Virologyhttp://dx .doi.org/10.1016/j.virol.2010.06.027.
44. Zolla-Pazner S, Kong XP, Jiang X, Cardozo T, Nadas A, Cohen S, Totrov M, Seaman MS, Wang S, Lu S.2011. Cross-clade HIV-1 neu-tralizing antibodies induced with V3-scaffold protein immunogens fol-lowing priming with gp120 DNA. J Virol85:9887–9898.http://dx.doi.org /10.1128/JVI.05086-11.
45. Bohl C, Bowder D, Thompson J, Abrahamyan L, Gonzalez-Ramirez S, Mao Y, Sodroski J, Wood C, Xiang SH.2013. A twin-cysteine motif in the V2 region of gp120 is associated with SIV envelope trimer stabilization. PLoS One8:e69406.http://dx.doi.org/10.1371/journal.pone.0069406.