The Membrane-Proximal “Stem” Region Increases the Stability of the
Flavivirus E Protein Postfusion Trimer and Modulates Its Structure
Karin Stiasny, Stefan Kiermayr, Andrea Bernhart, Franz X. Heinz
Department of Virology, Medical University of Vienna, Vienna, Austria
The flavivirus fusion protein E contains a “stem” region which is hypothesized to be crucial for driving fusion. This sequence element connects the ectodomain to the membrane anchor, and its structure in the trimeric postfusion conformation is still poorly defined. Using E trimers of tick-borne encephalitis virus with stem truncations of different lengths, we show that the N-terminal part of the stem increases trimer stability and also modulates the trimer structure outside the stem interaction site.
F
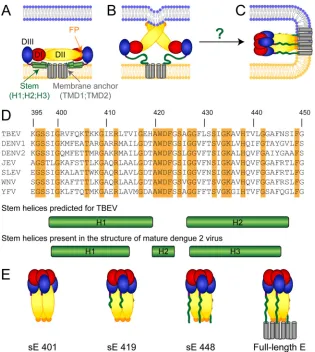
laviviruses, members of a genus in the family Flaviviridae, comprise a number of medically relevant viruses, such as den-gue virus, yellow fever virus, Japanese encephalitis virus, West Nile virus, and tick-borne encephalitis virus (TBEV) (1). These small enveloped viruses infect cells by receptor-mediated endocy-tosis and acidic-pH-induced membrane fusion in endosomes (2,3). The fusion reaction is mediated by the major envelope protein E, a class II fusion protein (4), which contains three domains (DI, DII, and DIII), a membrane-proximal region (the so-called “stem”), and the carboxy-terminal transmembrane anchor (Fig. 1A).
The current flavivirus fusion model (Fig. 1AtoC) is based on atomic structures of truncated dimeric prefusion and trimeric postfusion structures of E that lack the stem-anchor region (solu-ble E [sE]). According to this model, the E dimers dissociate into monomers at acidic pH, leading to the exposure of the fusion peptide (FP) and its insertion into the outer leaflet of the target membrane (Fig. 1B) (5). Membrane merger has been proposed to be driven by the relocation of DIII and a subsequent “zippering” reaction of the stem along DII, thus yielding a hairpin-like post-fusion trimer with the FPs and transmembrane domains at the same end of the molecule (Fig. 1C; reviewed in references4and6). In accord with this model, soluble forms of DIII containing part of the stem and stem-derived peptides inhibited fusion (7–10).
As shown by cryo-electron microscopy (cryo-EM) of dengue virions, the prefusion stem consists of two amphipathic helices (H1 and H3) that lie flat on the viral membrane and flank an additional short perimembrane helix (H2) (11,12) (Fig. 1AandD, lower panels). H2 has been resolved in the 3.5-Å structure of den-gue virus only recently (12). With the exception of the first few amino acids following the C terminus of the E ectodomain (13,
14), details of the stem in the context of the postfusion trimer are presently unknown. In X-ray crystallographic analyses of trun-cated dengue virus sE trimers that included N-terminal portions of the stem, most of the stem parts were disordered (14). The objective of our work was to generate experimental evidence for interactions of the stem with the core of the postfusion E trimer. For this purpose, we analyzed E trimers of TBEV with different C-terminal truncations using two approaches: (i) we assessed the impact of the presence of the stem as well as parts thereof on the thermostability of E trimers, and (ii) we examined possible influ-ences of stem interactions on the E trimer structure with confor-mation-specific monoclonal antibodies (MAbs). Consistent with the hypothesis of stem-DII interactions during fusion, we show
that the presence of the stem, in particular, its N-terminal but not its C-terminal part, increases the thermostability of the trimer. Furthermore, these interactions also appear to modulate the structure of the whole E trimer as shown by substantial differences in the reactivities of two MAbs specific for DIII epitopes that lie outside the proposed stem-DII binding site.
For our studies, we generated four different E protein forms of TBEV (strain Neudörfl; GenBank accession numberU27495) in their trimeric conformations (Fig. 1E): (i) virion-derived sE lack-ing the whole stem-anchor region (sE 401) (13,15), (ii) recombi-nant sE 419 (truncated after amino acid 419), (iii) recombirecombi-nant sE 448 (truncated after amino acid 448), and (iv) virion-derived full-length E (16). Since the presence of helices in the stem has been described only in the prefusion conformation of dengue virus E (11,12) (Fig. 1D) but is unknown in the postfusion structure, we have based the design of the truncated forms of E trimers on sec-ondary-structure predictions (13) (Fig. 1D). According to this ra-tionale, we used truncations after amino acid 419 (sE 419 contain-ing H1 of the prefusion form) and after amino acid 448 (sE 448 containing the entire stem) (Fig. 1D).
Virion-derived sE 401 trimers and full-length E trimers were produced as previously described (15,16). Briefly, sE dimers— obtained from virions by limited proteolysis (17)—were incu-bated with liposomes at acidic pH and membrane-bound trimers were isolated by solubilization with n-octylglucoside (n-OG) and ultrafiltration (15). Full-length E trimers were prepared by solu-bilization of purified low-pH-treated virions and rate zonal su-crose density gradient centrifugation (16,18).
The recombinant sE 419 and sE 448 proteins were produced in
DrosophilaSchneider 2 (S2) cells using the pT389 vector (kindly
provided by Thomas Krey and Felix Rey, Institut Pasteur, France) containing the coding sequences for prM and truncated E. The sE proteins had either one (sE 448) or two (sE 419) cleavable C-ter-minal Strep-tags for purification. The S2 cells stably expressing the proteins were generated by cotransfection with a blasticidin selec-tion vector as previously described (19). Seven to 11 days after
Received13 May 2013Accepted20 June 2013
Published ahead of print26 June 2013
Address correspondence to Karin Stiasny, [email protected]. Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01283-13
on November 7, 2019 by guest
http://jvi.asm.org/
induction of expression by CuSO4, the proteins were purified by
Streptactin affinity chromatography (IBA BioTAGnology, Ger-many).
The sE 419 protein was predominantly secreted as a dimer, but a small trimer peak was also observed when the cell culture super-natant was analyzed by sedimentation in sucrose gradients (Fig. 2A). The sE 448 protein, on the other hand, was already a trimer in the cell culture supernatant (Fig. 2B). Trimerization was likely to be induced by the moderately acidic pH of the insect cell cultures or during passage through the slightly acidic trans-Golgi network, and the length of the stem apparently influences the efficiency of this conversion.
After removal of the Strep-tag from sE 419 by enterokinase cleavage at 4°C, the tagless proteins were converted quantitatively into trimers by acidification in the presence of liposomes followed
by solubilization with n-OG according to the protocol established for virion-derived sE 401 (15). sE 448 was purified from the me-dium by Streptactin chromatography in the presence of 1% n-OG to prevent aggregation. For MAb binding studies, we also ex-pressed a tagless protein that was purified by rate zonal gradient centrifugation (16,18).
The oligomeric state of the different trimer preparations was determined by sedimentation analyses as well as chemical cross-linking with dimethyl suberimidate (DMS) followed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) as previously described (18). In sucrose gradients, all four prepa-rations sedimented to positions corresponding to a trimer (Fig. 2C
toF) and cross-linking resulted in a pattern typical for E trimers in SDS-PAGE (18) (Fig. 2CtoF, insets).
In our previous work, we demonstrated that full-length E
trim-FIG 1Schematic representations of the flavivirus E protein. Color code for protein E: domain I (DI), red; domain II (DII), yellow; domain III (DIII), blue; fusion peptide (FP), orange; stem helices 1 (H1), 2 (H2), and 3 (H3), green; transmembrane domains 1 (TMD1) and 2 (TMD2), gray. (A to C) Diagram of the full-length E protein before fusion (A), at an intermediate stage (B), and after fusion (C). (A) E dimer at the surface of mature virions with the stem helices lying flat on the viral membrane (light orange). (B) Acidic-pH-induced E dimer dissociation and insertion of the FPs into the target membrane (blue). (C) Hairpin-like postfusion trimer with the FPs and the membrane anchor juxtaposed in the fused membrane. The arrangement of the stem is not known (indicated by the green wavy lines). (D) Amino acid sequence alignment of stem regions (amino acids 395 to 450) of the following flavivirus E proteins (TBEV numbering) was performed using the program ClustalW: TBEV (GenBank accession numberU27495), dengue virus type 1 (DENV1) (GenBank accession numberAB189120), DENV2 (GenBank accession numberM19197), Japanese encephalitis virus (JEV) (GenBank accession numberAF315119), Saint Louis encephalitis virus (SLEV) (GenBank accession numberM16614), West Nile virus (WNV) (GenBank accession numberAF206518), and yellow fever virus (YFV) (GenBank accession numberX03700). Identical amino acids are highlighted in orange. The stem helices based on predictions for TBEV (13) (upper panel) as well as the stem helices resolved in the structure of mature dengue 2 virions (12) (lower panel) are indicated below the alignment. (E) Schematic representations of the four different trimer preparations used in this study (left to right): sE 401 lacking the stem-anchor region; sE 419 containing the N-terminal part of the stem; sE 448 containing the whole stem; full-length E. The stem regions are indicated by green wavy lines.
Stiasny et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.135.450.64.416.2]FIG 2Sedimentation analyses ofDrosophilaS2 cell culture supernatants (CC-SN) containing sE 419 (A) or sE 448 (B) proteins as well as of purified (s)E trimer preparations (C to F). The sedimentation direction is from left to right. The different preparations were subjected to sedimentation in 7% to 20% sucrose gradients in TAN buffer (pH 8.0) (50 mM triethanole amine; 100 mM NaCl) containing 0.1% Triton X-100 (18,27). The gradients were fractionated, and the amount of (s)E protein in each fraction was determined by a quantitative four-layer ELISA after denaturation of the samples with 0.4% SDS (28). The inset shows the Western blot analysis after chemical cross-linking of the purified trimers with dimethylsuberimidate (18). The positions of monomers (M), dimers (D), and trimers (T) in the gradients and blots are indicated. Representative examples of at least two independent experiments are shown.
Flavivirus Postfusion E Trimer Stem Interactions
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.134.450.62.638.2]ers were resistant to thermal denaturation up to a temperature of 70°C (16). sE trimers lacking the stem-anchor region (sE 401) were stable only up to 60°C (16), and preliminary experiments with sE 419 had indicated that it was stable up to at least 65°C (data not shown). Based on these results, we determined the thermosta-bility endpoints of sE 401, sE 419, sE 448, and full-length E trimers more precisely by exposure to 70°C and 75°C. The effect of this treatment on the E trimer peaks—in comparison to incubation at 37°C (control temperature)—was assessed by sucrose density gra-dient analyses (16,20). As expected, sE 401 trimers were strongly denatured at 70°C, with only a small residual trimer peak (Fig. 3). In contrast, approximately 90% of sE 419, sE 448, and full-length E still sedimented as trimers after incubation at 70°C; denatur-ation was observed only at 75°C in these cases (Fig. 3), indicating enhanced thermostabilities compared to sE 401. This shows that the N-terminal part of the stem alone accounted for the increased observed thermostability. Consistent with these results, a single amino acid substitution within this region (F403I) strongly re-duced the thermostability of the TBE full-length E trimer (20). Interestingly, stability appeared to decrease slightly with increas-ing length of the stem-anchor region, although these differences were statistically not significant (analysis of variance [ANOVA];
P⫽0.15).
To obtain information about a possible structural influence of stem-DII interactions on the whole E trimer, we probed the four trimers with five well-characterized conformation-dependent E protein-specific MAbs which recognize epitopes in domains II and III (Fig. 4A): MAb A1 (epitope located at the tip of DII that includes the FP), MAb A2 (epitope located in DII between the epitopes of MAbs A1 and A3 and involving the FP), MAb A3 (epitope located in DII involving the b- and d-sheets as well as the bc-loop [21]), MAb B4 (epitope located at the lateral ridge of DIII), and MAb IM3 (epitope located in DIII involving the de-loop [21]). The MAb binding sites have been mapped by the use of
competition assays, neutralization-escape mutants, and specifi-cally engineered mutations (22–26). In previous work, we have demonstrated that these MAbs react with sE 401 dimers as well as full-length E dimers and thus recognize the prefusion form of E (25,26). To compare the reactivities of the four different trimers with these MAbs, we used a blocking enzyme-linked immunoas-say (ELISA) format and measured whether the binding of the MAbs to virions coated to the solid phase was inhibited by the trimers in solution (25,26). The results of these assays are shown inFig. 4DtoG.
The completely truncated sE 401 trimer—for which the crystal structure is known (13)—reacted with all five MAbs and blocked their binding to the virion (Fig. 4D). The presence of the N-ter-minal part of the stem in sE 419, however, led to a strongly reduced binding of MAbs A2, A3, B4, and IM3 (Fig. 4E). Only MAb A1— directed to the FP at the tip of DII (Fig. 4A)—was not affected and still reacted with sE 419 in an unimpaired manner. In the case of A2 and A3, whose epitopes are located upstream of A1 in DII and competitively inhibit each other (22) (Fig. 4A), the blocking effect might be caused by steric hindrance of antibody binding, consid-ering that residues 402 to 419 may extend to the A3 epitope, as suggested by modeling (13) (Fig. 4B). Alternatively, the reduced reactivity of A2 and A3 could also be caused by an interaction-induced conformational change in DII. In the case of B4 and IM3, however, the epitopes lie in DIII and thus outside the possible stem-trimer core interactions (Fig. 4AandB). This makes epitope shielding highly unlikely and provides strong evidence for a mod-ulation of the E trimer structure through the interaction of the N-terminal part of the stem with DII, which was also shown to increase trimer stability (Fig. 3). This structural feature was also maintained in sE 448 (Fig. 4F) and full-length E (Fig. 4G), which additionally lost the binding of the FP-specific MAb A1. The most likely interpretation of these results is that the epitope accessibility
FIG 3Thermostability of truncated sE trimers and full-length E trimers. Trimers were exposed to 37°C, 70°C, or 75°C and subjected to rate zonal sucrose gradient centrifugation as described in the legend toFig. 2. The extents of trimer stability are expressed as percentages of E found in the trimer peak fractions after incubation at elevated temperatures (70°C and 75°C) relative to the E protein present in the trimer peak fractions after incubation at 37°C (set to 100%). Data are from at least three independent experiments, and the error bars represent the standard errors of the means (SEM). n.t., not tested.
Stiasny et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.137.451.67.284.2]of A1 is decreased by interactions of the C-terminal part of the stem with DII residues around the FP at the tip of the trimer.
Consistent with our experimental evidence for interactions of the N-terminal part of the stem with the core of the E trimer, modeling of residues 402 to 412 of the stem of TBEV as a helix revealed an excellent fit with conserved residues in DII (13). The fit was less satisfactory for residues 413 to 419, and it was specu-lated that— because of stem interactions—the structure of the DII part of the full-length E trimer could be slightly different from the crystal structure of the truncated sE 401 trimer (13).
At present, there is still no direct structural evidence that the N-terminal part of the stem forms a helix in the postfusion trimer, as suggested in the modeling study (13). In crystal structures of den-gue virus trimers containing N-terminal portions of the stem, only the first stem residues (up to amino acid 404 [TBEV numbering]) exhibited intra- and intermolecular interactions with DII, whereas the remaining residues were disordered (14). A loop in DIII, however, was in a different conformation in these structures compared to the stemless trimer (14). This may be due to crystallizing of the trimers at different pHs (14) but could also indicate that the presence of the
FIG 4Surface representations of the TBEV sE trimer (Protein Data Bank [PDB] accession no. 1URZ) (A, B, and C) and blocking ELISAs with truncated sE trimers as well as full-length E trimers (D to G) performed using five conformation-sensitive MAbs. (A, B, and C) The color code for protein E is as described for Fig. 1, with the linkers between DI and DIII as well as DIII and the stem shown in purple. The binding sites of the MAbs are indicated in cyan. (B) Hypothetical run of stem residues 402 to 419 as a helix as suggested by modeling (13). The last residue resolved in the sE structure (sE 401) is highlighted. (C) Hypothetical runs of the stem as suggested by Klein et al. (14) are indicated in green and labeled with a question mark. (D to G) Serial dilutions of the trimers were preincubated with a predetermined concentration of the respective MAbs, and the fraction of MAb not blocked by the antigen was detected in ELISA with TBEV-coated plates (25,26). The results were expressed as “% MAb Reactivity,” defined as the quotient of absorbance in the presence of the blocking antigen and the absorbance in the absence of the blocking antigen as follows: (A490in the presence of blocking antigen/A490in the absence of blocking antigen)⫻100. Data are from at least three
independent experiments, and the error bars represent the standard errors of the means (SEM).
Flavivirus Postfusion E Trimer Stem Interactions
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.100.488.63.511.2]stem indeed influences the conformation or position of DIII as shown by our MAb B4 and IM3 binding data (Fig. 4).
In summary, we provide evidence for contacts of the N-termi-nal part of the stem with the trimer core that not only increase the thermostability of the trimer but also modulate the structure out-side the putative stem-core binding site compared to stemless sE trimers. The C-terminal part of the stem seems to be involved in interactions with the tip of DII, including the FP. Our data are consistent with a model for the full-length E trimer proposed by Klein et al. (14) in which the first amino acids of the stem zipper along DIIs, the central part loops out, and C-terminal stem resi-dues contact the FP region (Fig. 4C). To define the precise contacts of the stem with DII as well as the effect of stem packing on the overall trimer conformation, high-resolution structures of trim-ers containing the whole stem region or at least parts thereof will be necessary.
ACKNOWLEDGMENTS
We thank Walter Holzer, Cornelia Stöckl, and Andrea Reiter for their excellent technical assistance as well as Beate Geller and Agnes Leitner for their help in generating expression plasmids.
This work was supported by the Austrian Science Fund FWF (P22914-B13).
REFERENCES
1.Simmonds P, Becher P, Collett MS, Gould EA, Heinz FX, Meyers G, Monath T, Pletnev A, Rice CM, Stiasny K, Thiel HJ, Weiner A, Bukh J.
2011. Family Flaviviridae, p 1003–1020.InKing AMQ, Lefkowitz E, Ad-ams MJ, Carstens EB (ed), Virus taxonomy. IXth Report of the Interna-tional Committee on Taxonomy of Viruses. Elsevier Academic Press, San Diego, CA.
2.Kaufmann B, Rossmann MG.2011. Molecular mechanisms involved in the early steps of flavivirus cell entry. Microbes Infect.13:1–9.
3.Smit JM, Moesker B, Rodenhuis-Zybert I, Wilschut J.2011. Flavivirus cell entry and membrane fusion. Viruses3:160 –171.
4.Harrison SC.2008. Viral membrane fusion. Nat. Struct. Mol. Biol.15:
690 – 698.
5.Stiasny K, Kossl C, Lepault J, Rey FA, Heinz FX.2007. Characterization of a structural intermediate of flavivirus membrane fusion. PLoS Pathog.
3:e20. doi:10.1371/journal.ppat.0030020.
6.Pierson TC, Kielian M.2013. Flaviviruses: braking the entering. Curr. Opin. Virol.3:3–12.
7.Liao M, Kielian M.2005. Domain III from class II fusion proteins func-tions as a dominant-negative inhibitor of virus membrane fusion. J. Cell Biol.171:111–120.
8.Liao M, Sanchez-San Martin C, Zheng A, Kielian M. 2010. In vitro reconstitution reveals key intermediate states of trimer formation by the dengue virus membrane fusion protein. J. Virol.84:5730 –5740. 9.Schmidt AG, Yang PL, Harrison SC.2010. Peptide inhibitors of
dengue-virus entry target a late-stage fusion intermediate. PLoS Pathog.
6:e1000851. doi:10.1371/journal.ppat.1000851.
10. Schmidt AG, Yang PL, Harrison SC.2010. Peptide inhibitors of flavivirus entry derived from the E protein stem. J. Virol.84:12549 –12554. 11. Zhang W, Chipman PR, Corver J, Johnson PR, Zhang Y,
Mukhopad-hyay S, Baker TS, Strauss JH, Rossmann MG, Kuhn RJ.2003. Visual-ization of membrane protein domains by cryo-electron microscopy of dengue virus. Nat. Struct. Biol.10:907–912.
12. Zhang X, Ge P, Yu X, Brannan JM, Bi G, Zhang Q, Schein S, Zhou ZH.
2013. Cryo-EM structure of the mature dengue virus at 3.5-A resolution. Nat. Struct. Mol. Biol.20:105–110.
13. Bressanelli S, Stiasny K, Allison SL, Stura EA, Duquerroy S, Lescar J, Heinz FX, Rey FA.2004. Structure of a flavivirus envelope glycoprotein in its low-pH-induced membrane fusion conformation. EMBO J.23:728 – 738.
14. Klein DE, Choi JL, Harrison SC. 2013. Structure of a dengue virus envelope protein late-stage fusion intermediate. J. Virol.87:2287–2293. 15. Stiasny K, Bressanelli S, Lepault J, Rey FA, Heinz FX.2004.
Character-ization of a membrane-associated trimeric low-pH-induced form of the class II viral fusion protein E from tick-borne encephalitis virus and its crystallization. J. Virol.78:3178 –3183.
16. Stiasny K, Kossl C, Heinz FX.2005. Differences in the postfusion con-formations of full-length and truncated class II fusion protein E of tick-borne encephalitis virus. J. Virol.79:6511– 6515.
17. Heinz FX, Mandl CW, Holzmann H, Kunz C, Harris BA, Rey F, Harrison SC.1991. The flavivirus envelope protein E: isolation of a solu-ble form from tick-borne encephalitis virus and its crystallization. J. Virol.
65:5579 –5583.
18. Allison SL, Schalich J, Stiasny K, Mandl CW, Kunz C, Heinz FX.1995. Oligomeric rearrangement of tick-borne encephalitis virus envelope pro-teins induced by an acidic pH. J. Virol.69:695–700.
19. Zlatkovic J, Stiasny K, Heinz FX.2011. Immunodominance and func-tional activities of antibody responses to inactivated West Nile virus and recombinant subunit vaccines in mice. J. Virol.85:1994 –2003. 20. Pangerl K, Heinz FX, Stiasny K.2011. Mutational analysis of the
zipper-ing reaction durzipper-ing flavivirus membrane fusion. J. Virol.85:8495– 8501. 21. Rey FA, Heinz FX, Mandl C, Kunz C, Harrison SC.1995. The envelope
glycoprotein from tick-borne encephalitis virus at 2 A resolution. Nature
375:291–298.
22. Guirakhoo F, Heinz FX, Kunz C.1989. Epitope model of tick-borne encephalitis virus envelope glycoprotein E: analysis of structural proper-ties, role of carbohydrate side chain, and conformational changes occur-ring at acidic pH. Virology169:90 –99.
23. Mandl CW, Guirakhoo F, Holzmann H, Heinz FX, Kunz C. 1989. Antigenic structure of the flavivirus envelope protein E at the molecular level, using tick-borne encephalitis virus as a model. J. Virol.63:564 –571. 24. Holzmann H, Stiasny K, York H, Dorner F, Kunz C, Heinz FX.1995. Tick-borne encephalitis virus envelope protein E-specific monoclonal an-tibodies for the study of low pH-induced conformational changes and immature virions. Arch. Virol.140:213–221.
25. Stiasny K, Kiermayr S, Holzmann H, Heinz FX.2006. Cryptic properties of a cluster of dominant flavivirus cross-reactive antigenic sites. J. Virol.
80:9557–9568.
26. Kiermayr S, Stiasny K, Heinz FX.2009. Impact of quaternary organiza-tion on the antigenic structure of the tick-borne encephalitis virus enve-lope glycoprotein E. J. Virol.83:8482– 8491.
27. Stiasny K, Allison SL, Schalich J, Heinz FX.2002. Membrane interac-tions of the tick-borne encephalitis virus fusion protein E at low pH. J. Virol.76:3784 –3790.
28. Heinz FX, Stiasny K, Puschner-Auer G, Holzmann H, Allison SL, Mandl CW, Kunz C.1994. Structural changes and functional control of the tick-borne encephalitis virus glycoprotein E by the heterodimeric as-sociation with protein prM. Virology198:109 –117.
Stiasny et al.


![FIG 4 Surface representations of the TBEV sE trimer (Protein Data Bank [PDB] accession no](https://thumb-us.123doks.com/thumbv2/123dok_us/149081.26225/5.585.100.488.63.511/surface-representations-tbev-trimer-protein-data-bank-accession.webp)