Copyright @ 1970 American Society for Microbiology Printed .n U.S.A.
Proteins Specified by Herpes Simplex Virus
II.
Viral Glycoproteins Associated
with
Cellular
Membranes
PATRICIA G. SPEAR, JOHN M. KELLER, AND BERNARD ROIZMAN
DepartmentofMicrobiology, University of Chicago, Chicago, Illinois 60637
Received for publication 10 November 1969
Membranes prepared from HEp-2 cells infected with herpes simplex virus and
free from soluble proteins, virus, ribosomes, and other cellular constituents were
solubilized and subjectedto electrophoresis onacrylamide gels. The electrophero-grams showed thefollowing. (i) The synthesis of host proteins and glycoproteins ceases after infection. However, the spectrum ofhost proteins in membranes re-mainsunaltered. (ii)Between 4and22hr postinfection, at leastfour glycoproteins are synthesized and bound to the smooth cytoplasmic membranes. On
electro-phoresis, theseglycoproteins formtwomajor andtwominor bands in the geland
migrate with proteins ranging from50,000to100,000daltons in molecular weight.
(iii) Thesameglycoproteins are presentin all membranes fractionatedby density and inpartially purified virus. The implications ofthedataarediscussed.
The preceding paper of this
series
dealt with the proteins synthesized in HEp-2 cells infectedwith
herpessimplex virus (29). In that paper, we showed thatproteins
aremade inthe cytoplasm andselectively
migrate into the nucleus. The pro-teinssynthesized
after infection differ from thoseof
uninfected cells. Most of theproteins
madeafter
4hrofinfection are structural components of thevirion.
We havesubsequently reported
that some ofthe
proteins
made ininfected
cellsbind
tomembranes
(P.
G. Spear, S. B.Spring,
andB.
Roizman,
Bacterial. Proc., p.170,
1969).
In this paper, we
describe
in greaterdetail
the nature andproperties of
theproteins
andof
the membranes towhich theseproteins
arebound.Thecircumstances
which
led us toexamine themembranes
of infected cells are of interest. Work in this and in other laboratories fumished ample evidence that the membranes ofcells
infected withherpesviruses
mustnecessarily
becomemodi-fied after
infection.
The evidence comes from three series ofobservations.
(i)
Thenucleocapsid
acquires
aninner
envelope in the nucleus andatthe same
time becomes infectious (30;
B.Roiz-man, S. B.
Spring,
and J. Schwartz,Fed.
Proc.,inpress). However, theouterenvelopeisderived fromamembraneof the cell.
Although
somevirus strains obtain their envelopes from the innerlamella
of the nuclear membrane (7, 17, 25,26,
31), others appeartoutilizeallmembranesof infected cells (10, 24,27).
These observations in-dicate that the virusspecifies
the site of envel-opment,and,
moreover, electronmicroscopic
observations suggest that the membranes are
modified at the time of envelopment. (ii) After envelopment, thevirion is usually found in struc-tures delineated by a membrane. Recent studies (23) have shown thatthese structures are prob-ably modified
endoplasmic
reticulum and serve as channels or ducts for the egress of the virus from infected cells. (iii) Herpes simplex viruses have been known for many years to alter the"social
behavior" ofinfectedcells. These
altera-tions
are specified by the virus andrange
fromrounding
of cells with little or noclumping
toformation of
looseclumps,
tight clumps, and,
finally, fusion of cells (9,
20).
Concurrent with thechange in the socialbehavior,
themembranes
of infected cells become modified with respectto
both structure and
immunological
specificity (19,
21). (iv)
Histochemical
studies
on humanamnion cells infected with herpes simplex virus(22)
re-vealed the appearance of
glycoprotein
material inthe nucleus and aproliferationof
glycoprotein
structures distributed around the nucleus and in thecytoplasm.
In
this
andsubsequent
papersof
theseries,
wehope
toelucidate
thebiochemical basis for
thealteration of
the membranes in herpesvirus-in-fected ceUs.MATERIALS AND METHODS
Media andsoutions.Minimal essential mediumof Eagle (8) was obtained from
Microbiological
Asso-ciates, Inc., Bethesda, Md., and supplemented with 10% calf serum for the growth of cells. Complete mixture 199 and a special formulation lackingar-ginne,
leucine, isoleucine, andvaline were obtained123
on November 11, 2019 by guest
http://jvi.asm.org/
SPEAR, KELLER, AND ROIZMAN
from Grand Island Biological Co., Grand
Island,
N.Y. Solutions of reagent-grade urea (8 M) were deionized onmixed-bed columns ofanion-exchange resin (Amberlite MB-1).
Chemicals. Special mixtures of leucine, isoleucine, andvalinecontaining equalactivities ofeither the 3H or14Cisotope werepreparedfor usby Schwarz Bio-Research, Inc., Orangeburg, N.Y. D-Glucosamine-6-8H(>200mc/mmole)wasfurnished by New England Nuclear Corp., Boston, Mass. Eastman Organic Chemicals, Rochester, N.Y.,suppliedtheN, N,N', N'-tetramethylethylenediamine. Acrylamideand ethylene diacrylate wereobtained from K & K Laboratories, Inc., Plainview, N.Y.
Cells. HEp-2 cells were obtained from Microbio-logical Associates,Inc.
Virus andinfection of cells. The Fstrainof herpes simplex viruswasisolated fromarecurrenteruption onthe face and waspassagedinHEp-2cells less than sixtimes. The effectsofthe F strain on thesocial be-haviorofinfected cellsandtheimmunological proper-tiesofthevirus werepreviously described (9). Inall oftheexperiments described here, HEp-2 cellswere
infected by exposing monolayer cultures (approxi-mately4X 107cells) for1hrat37 C tosufficientvirus to yield effective multiplicities of approximately 20
(plaque-forming units) PFU/cell. The inoculumwas
then aspirated and replaced with 35 ml of mixture 199 supplemented with 1% calf serum. Subsequent manipulations of the infected cells are described in thefollowing sections.
Labeling of cells with radioactiveisotopes.Infected or uninfected cells were radioactively labeled with aminoacids orglucosamine,orboth, duringthetime intervals indicated below. The special 199 mixture, supplemented with one-tenth the usual concentra-tions of leucine, isoleucine, and valine, was used to incubate the cells with the 3H or 14C mixtures of leucine, isoleucine, andvaline.Completemixture 199 or199deficient in the threeaminoacidswasusedto labelthecells withD-glucosamine-6-8H.
Preparationofvirus.Infectedcells were washed two timeswithisotonic salinecontaining 108M calcium acetate buffer, pH 7.0. The washes and the
extra-cellular fluid were combined and centrifuged at
4,000X gfor10min to removecells and debris. The supernatantfluidwasthen layered on3-mlcushions
of 60% (w/w) sucroseincellulosenitrate tubes[1 by
3.5inches(2.54 by 8.9cm)]andcentrifugedat81,500 X g (average) and4 Cfor 2 hr in the SW 27rotor.
Fromeachtube, the visible band on topofthe60% sucrosecushion wasaspiratedwithasyringe, diluted with2volumes of0.05M
tris(hydroxymethyl)amino-methane (Tris) buffer (pH 7.0), and centrifuged at 55,000 X g (average) and 4 Cfor 1 hr. The pellet containingviruswas suspended inasmallvolumeof waterfor solubilization.
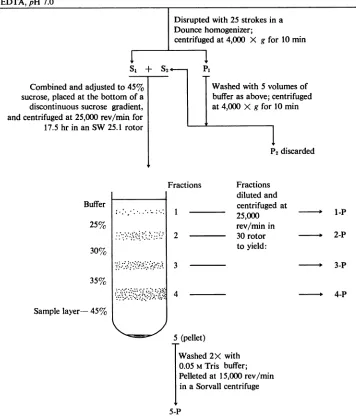
Preparation of membrane fractions. Smooth mem-branes were purified from infected and uninfected cellsbyaproceduredevelopedby Bosmannetal. (2) for the isolationofsmoothendoplasmicreticulumand plasma membranes from HeLacells. A flowdiagram ofthe steps in the procedure is presentedin Fig. 1. Briefly, 20- to 22-hr infected cells were washed twice
with saline containing 103 M calcium acetate buffer (pH 7.0) and then suspended in approximately 10 volumes of 0.02 M Tris buffer (pH 7.0) containing 0.01 Methylenediaminetetraacetic acid (EDTA). The cells weredisrupted in a tight-fitting Dounce homog-enizer; homogenization was monitored by phase microscopyto avoidexcessive nuclearbreakage. The homogenate was centrifuged at 4,000 X g for 10 min to removenuclei and debris. In someexperiments, the
pellet (P1)waswashed once with 5 volumes of buffer and thesupernatantfluids (Si and S2) were combined andmade 45% (w/w) with respect to sucrose. About 10-ml samples of the 4,000 X g supematant fluid containing 45% (w/w) sucrose were placed in the bottom of cellulose nitrate tubes [1 by 3 inches (2.54 by 7.62 cm)] and overlaid with 6.5-ml amounts of 35and30% sucrose, 5 ml of 25% sucrose, andfinally
2.5milof 0.05 M Tris buffer (pH 7.0). The discontinu-ous gradients were then centrifuged in the SW 25.1 rotor forapproximately20 hr at64,000 X g (average) and 4 C.Aftercentrifugation,four visible bands and a
pellet were obtained. The bands (fractions 1 to 4) were carefully aspirated with a syringe, and corre-sponding fractions from replicate gradients were pooled. Fractions 1 to 4 were diluted with approxi-mately 4volumes of 0.05 M Tris buffer andcentrifuged at 55,000 X g(average) and 4 Cfor 1 hrin the 30 rotor. The pellets (fraction1-Pto 4-P) weresuspended in a small volume of 0.05 M Tris buffer. The pooled pellets (fraction 5) from the sucrose gradients were washed two times with 0.05 M Trisbuffer andpelleted at 27,000 X g for 10 min in a Sorvall centrifuge (fraction 5-P). As described below, purified smooth membranes were present in fraction 2-P and mostof theinfectious virus wasfound in fraction 4-P.
Acrylamide gelelectrophoresis. Fractions obtained during the course of membrane purification were solubilized for gel electrophoresis as described by Summers et al. (34). Briefly, sodium dodecyl sulfate (SDS) wasadded to the cell fractions to yield afinal concentration of 1 to 2%; the extracts were then acidified by the addition of a one-tenth volume of glacialacetic acid. Urea and,-mercaptoethanol were
added to final concentrations of 0.5 M and 1%, respectively. After incubation at 60 C for 1 hr, the extracts weredialyzed for 15 to 20 hragainst0.01 M sodium phosphate buffer (pH 7.1) containing 0.1% SDS, 0.5 M urea, and 0.1% ,B-mercaptoethanol. The extractswerethendialyzedforanadditional3to 4 hr against the same buffer containing 10% sucrose; if necessary,theywereconcentratedby vacuumdialysis. Acrylamide gel electrophoresis was performed in a
continuous system essentially as described by Sum-mers etal.(34), except that the gels were0.6by 20 cm and consisted of 5.7% (w/v) acrylamide (5.3% acrylamide plus 0.4% ethylene diacrylate), 0.1% N, N, N', N', tetramethylethylenediamine, 0.1 M sodiumphosphatebuffer (pH 7.2), 0.5 M urea, 0.1% SDS, and 0.1% ammonium persulfate. Ethylene diacrylatewasused as across-linkingagent instead of the usual N-N-bismethylene acrylamide because gels
polymerizedwith theformer agentcanbesolubilized
124 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
PROTEINS SPECIFIED BY HERPES SIMPLEX VIRUS
Procedure forPurifying Cellular Membranes
16-hrinfectedcells,washedwithsaline-103 M calciumacetateand
suspended
in 10 volumes of 0.02 M Tris-0.01 M EDTA,pH 7.0Combinedandadjustedtot sucrose, placedatthebottom
discontinuoussucrose gradi andcentrifugedat25,000rev/min 17.5hrinanSW25.1 ri
Buffer
25%
30%
35%
Samplelayer-45%
Disruptedwith 25strokes in a Douncehomogenizer;
centrifugedat 4,000 X gfor 10 min
Si
+ S2 P145% Washedwith5volumesof
ofa bufferasabove;centrifuged
ient, at4,000 X gfor 10 min
lfor
otor
I
sP2discarded
Fractions Fractions diluted and centrifuged at
I - 25,000
rev/mmnin
2 30 rotor
toyield:
3
.4
- 1-P
b 2-P
3-P
4-P
5 (pellet)
Washed2Xwith 0.05 MTris buffer;
Pelletedat15,000rev/min inaSorvallcentrifuge
5-P
FIG.1. Flowdiagramofthe purification procedure used to obtain smooth membranes (2).
athigh pH (4), thereby facilitatingthe quantitation of radioactive proteins byliquid scintillation spectrom-etry.Theprotein samples (50 to 100 ,uliters),
contain-ingtrace amountsofbromophenolblue, were layered
ontopofthegelsandsubjectedtoelectrophoresisfor about 15 hr at a regulated voltage of 3 v/cm. The upper and lower buffer chambers contained 0.1% SDS in 0. 1 Msodium phosphatebuffer,pH 7.2. The
followingprocedureforquantitating radioactive pro-teinsinacrylamide gels by usinganonaqueousliquid scintillation system has been described elsewhere(28). (i)Thegelswereslicedinto 2-mmsegments,andeach
segmentwasplaced inascintillationvial. (ii) A2-ml
amount of concentrated ammonium hydroxide was
addedtoeachvial,and the vialswereagitatedatroom
temperatureuntil the segmentswerehydrolyzed. (iii} Astrip of glass fiberpaper (2 by4 cm; ReeveAngel, grade GF81)wasplacedin eachvial so that the paper touched thebottom and absorbed theliquid. (iv)The glassfiber wicks,still invials,wereheatedunder infra-redlampsuntiltheywerecompletely dry.(v) Scintilla-tionfluid [7.4 gof 2,5-diphenyloxazole plus 0.3 gof 1,4-bis-2-(5-phenyloxazolyl)-benzene in 1 liter of toluene]wasaddedto coverthewicks.
5, 125
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.496.74.430.100.519.2]SPEAR, KELLER, AND ROIZMAN
Electron microscopy of membrane fractions. Sam-ples of the various fractions from infected or unin-fected cellswerediluted withwaterand centrifugedat
27,000X gfor1 hrin the Sorvall centrifuge. The
pel-lets were then suspended in 1.6% osmium tetroxide
preparedin 0.067Ms-collidinebuffer, pH 7.2. After 1
hr at 0 C, the membranes were removed from the osmium tetroxide solution by centrifugation. The pel-letswere then mixed with liquidlonagarmadeupin
1% uranyl acetate; afterthe agarhadsolidified, the
embeddedpelletswerecutinto 2-mm cubes. Theagar
cubes were soaked in a 1% uranyl acetate solution overnight at 4 C, dehydrated in a graded series of
ethyl alcohol, and embedded inEpon. Thin sections of the membraneswereprepared byusingaPorter-Blum
MT-2ultramicrotome andwereexamined after stain-ing with uranyl acetate and lead citrate on an AEI
EM-6Bmicroscope.
RESULTS
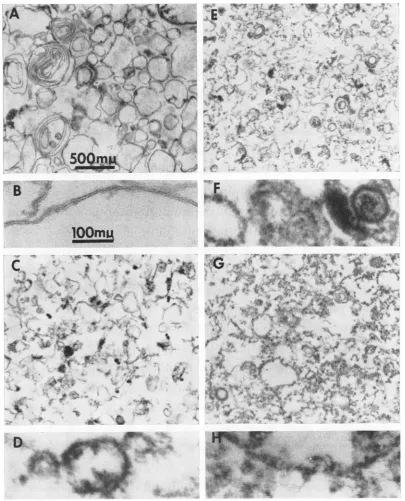
Purityof membrane preparation. Thefractions obtainedduring thecourseof membrane purifica-cationwere verysimilar in composition, with
re-spect to cellular constituents, to those obtained from HeLa cellsby Bosmannetal. (2). Electron micrographs of thin sections of the various frac-tions from infected cells are shown in Fig. 2. Note that 4-P and 5-P contained considerable amountsof virus (see also Table 1). In addition, fractions 3-P, 4-P, and 5-P consisted of mixtures of various kinds of membranes contaminated with other cytoplasmic constituents. However, the fraction ofgreatest interest, designated 2-P, containedonly smooth membranes. The degree of purity of this preparation isindicated by the fol-lowing observations. (i) Electron microscopy of preparations from both infected and uninfected cells failed to reveal contamination with ribo-somes or other cellular constituents. (ii) As shown in Table 1, less than 0.05% of the infectious virus put in the gradient was found in fraction 2-P. (iii) Soluble proteins (i.e., proteinsnot sedi-mentableat64,000 Xgfor 1hr) werenotfound infraction 2 or2-P (Table 1); these proteins re-mained in thesample layer (45% sucrose) during centrifugation.
Proteins incorporated into membranes of frac-tion 2-P after infection. To determine whether proteins synthesized afterinfection become bound
to the smooth membranes of fraction 2-P, ap-proximately 108 cells were incubated in medium containing 3H-glucosamine (2/,c/ml) and 14C_ leucine, -isoleucine, and -valine (0.5 ,uc/ml) from 4to22hr andfrom 6to22hr postinfection, re-spectively. Radioactive glucosamine was used as wellasaminoacids, sincemany membrane-asso-ciated proteins have been showntobe glycopro-teins. Membrane fraction 2-P was prepared, solubilized, and analyzed by gel electrophoresis.
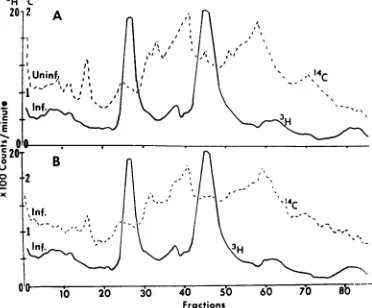
Theprofiles in Fig. 3 indicatethatboth
glucosa-mine and amino acids become incorporated into
proteins forming two major bands and possibly
two minor bands in the gel. Preliminary results indicate that the 3H-glucosamine isincorporated
into the membranes as glucosamine and
galac-tosamine. We concludethat three to four
glyco-proteins synthesized after infection become in-corporated into smooth membranes of thecell.
Are host membrane proteins displaced? It was of interest todetermine whether host proteins in themembranesaredisplacedbytheglycoproteins synthesized after infection. In an attempt to pro-vide information concerning this point, two ex-periments were done. In the first experiment,
three replicate sets of cultures (approximately
108 cellsin each set) were used. One set was incu-bated in medium containing '4C-leucine, -iso-leucine, and -valine (0.2 ,uc/ml) for 12 hr and then inmediumcontaining nonradioactive amino acids for anadditional 24hr. The second set was
incubated in medium containing the "4C-amino acids and in nonradioactive medium as above, exceptthatthe cultures were infected after 3 hr in
nonradioactive medium. The third set of cultures was infected and incubated with medium
con-taining 3H-leucine, -isoleucine, and -valine (5
Auc/
ml) from 8 to 21 hr postinfection. The cells in
each set of cultures were then harvested, mixed with approximately 3 x 108 uninfected or
in-fected unlabeled cells, and fractionated. Fraction 2-P from the 3H-labeled cultures (labeled after
infection) was then coelectrophoresed with each of the "4C-labeled fractions. Acomparison of the 14C profiles in Fig. 4 reveals that, by this
pro-cedure, one cannot demonstrate displacement of host membraneproteins synthesized during a 12-hr interval before infection by proteins
synthe-sized after infection. Since the host proteins in fraction 2-P are numerous and difficult to resolve,
in the second experiment, we traced thefate of
host glycoproteins only. In thisexperiment, two
replicate cultures, each
containing
3 x 107 cells,were incubated with 3H-glucosamine (3
Ac/ml)
for 23 hr. One culture was then incubated in me-dium containing nonradioactive glucosamine
(6,ug/ml) foranadditional 23hr; theotherwas
incubated for 1 hr in the presence of
nonradio-active glucosamine and then infected and incu-bated until 22 hrpostinfectionin the absence of
glucosamine. A third culture was labeled with
3H-glucosamine (3 ,uc/ml) from 4 to 22 hr
post-infection.Theelectrophoretic profilesofproteins containing 3H-glucosaminefrom fractions2-P of thecultureslabeledasdescribedarepresentedin
Fig.5.Again, there was noevidence ofa
displace-ment of membrane-bound proteins synthesized
before infectionby theglycoproteins synthesized
126 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
VOL.5,1970PROTEINS SPECIFIED BY HERPES SIMPLEX VIRUS12
fl
.E.¾
.
V
It
r N
)
I
2'
lOOmuA
CAW
.ttr
~~~P..
WXI
[image:5.496.42.447.68.569.2]Avs
v4v
...
%
FtAl
1~~~~ 4.~~~
0D
a6
FIG.2. Electronmicrographsofthinsectionsofmaterialfoundinfractions2-PthroughS-P.AanzdB,fraction 2-F; CandD,fraction3-F;EandF,fraction4-F; G andH,fractionS-P.A,C,E,anid G,magnzificationasshownt inA.B,D,F,andH,magnificationasshowninB.
after infection. It appears that the
proteins
made after infection are either added topreexisting
membranes or become
incorporated
intonewly
synthesized
membranes.Finally,
these twoex-periments demonstrate,
aswepreviously reported
(P.
G.Spear,
S.B.Spring,
and B.Roizman,
Bac-teriol.Proc.,
p.170,
1969),
that there is nore-semblance between the
profile
of hostproteins
VoL.
5,
1970127
.,X:
.k..
A.-,4:::
on November 11, 2019 by guest
http://jvi.asm.org/
SPEAR, KELLER, AND ROIZMAN
TABLE 1. Assays of infectious virus and protein during the courseof membrane purification
Fraction PFU Per cent of Protein Per cent of
initialPFU (mg) initialprotein
Cell homogenatepluswashings(S1 and 1,090X 106 100 85.4 100
SO)
Discontinuous sucrosegradient
1 0.04 X 106 0.004 0.42 0.5
2 0.54 X 106 0.05 1.63 1.9
3 18.5 X 106 1.7 1.69 2.0
4 292 X 106 26.8 16.00 18.7
5 (pellet) 41 X 106 3.8 4.45 5.2
Remainder ofgradient 153 X 106 14.0 49.80 58.3
Totals forsucrose gradient 505 X 106 46.4 74.0 86.6
Pellets offractions fromsucrose gradi-ent
1-P NDa 0.30 0.4
2-P ND 1.50 1.8
3-P ND 1.38 1.6
4-P ND 4.32 5.1
Washedpellet fromsucrosegradient
5-P 22 X 106 2.0 1.32 1.5
aAssay not done.
3H4c 16T4
*1213
182
0
~~~ ~ ~ ~ ~ ~ ~ ~ ~ aminoacids
3Hglucosamin
10 20 30 40 50 60 70 80
Fractions
FIG.3. Electrophoretic profiles ofthe proteins
synthe-sizedafter infection andincorporated into the smooth
membranes of fraction 2-P. Cells were incubated in
medium containing 3H-glucosamine and 14C-leucine,
-isoleucine, and-valinefrom4 to22hr andfrom 6 to 22 hrafter infection, respectively. Symbols: solidline, 8H-glucosamine; dashedline, IC-amino acids. In this
and allofthefollowingfigures,migrationofproteins is
from lefttorighttoward theanode.
from fraction 2-P and that of the glycoproteins synthesizedafterinfection.
Comparisonofglycoproteinsin thevarious bands obtained after flotation of membranes. To de-termine whether the glycoproteins found associ-ated with themembranes offraction2-Pare also associated with the other membranes from in-fectedcells,thefollowing experimentwasdone.A
culture containing 3 X 107 cells was incubated from 4 to 22 hrpostinfectioninmedium contain-ing 3H-glucosamine (3 ,uc/ml); the infected cells
were then fractionated as described above, and thevariousfractionswereanalyzed by gel electro-phoresis. Figure6 shows that thesame glycopro-tein bands are present in all membranes
frac-tionatedaccordingtodensity.
3H14c
Fractions
FIG.4. Pulse-chase experimenttodetermine whether host proteinsinthesmooth membranesaredisplacedby the proteins synthesized after infection. Fraction 2-P
from cells labeled with 3H-leucine, -isoleucine, and -valinefrom8 to21hrpostinfectionwassolubilizedand
coelectrophoresed with A, the proteinsoffraction2-P
from uninfectedcells labeled with thesame 14C-amino acidsfor12hr and then incubated with nonradioactive
mediumforanadditional24hr, and B, the proteins of
fraction2-Pfromcellslabeledas in A with 14C-amino acids and then infected after 3 hr in nonradioactive
medium. Symbols: solid line, 3H-leucine, -isoleucine, and -valine; dashed line, 14C-leucine, -isoleucine, and -valine.
128 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.496.56.256.100.442.2] [image:6.496.266.452.367.521.2]SIMPLEX VIRUS
4
4-3
0 0
K4 1
3- 0
2-°.
~1020
30
40
50 90 70 80Fractions
FiG.5. Pulse-chaseexperimenttodetermine whether host glycoproteins in the smooth membranes are dis-placedby glycoproteinssynthesizedafterinfection. The electrophoretic profiles are shown of8H-glucosamine incorporated into the proteinsoffraction2-Pfromcells treated asfollows. (A) Uninfectedcells were incubated withmedium containing 8H-glucosaminefor23 hr and with medium containing nonradioactive glucosamine for an additional 23 hr. (B) Cells were incubated with mediumcontaining WH-glucosaminefor23 hr andthen infected afterincubationfor1hr in medium containing nonradioactive glucosamine. (C) Cells were labeled with'H-glucosaminefrom 4 to 22 hr postinfection.
5
A
4
I
]Bond
Xit
0
Bond4
°1 10 20 30 40 50 6b 70 80
[image:7.496.47.238.70.263.2]Fractions
FIG. 6. Electrophoretic profiles of glycoproteins in
fractionsI-Pto4-P from cellsincubated with medium containingWH-glucosaminefrom 4 to 22 hrpostinfection.
Comparison of membrane glycoproteins with structural proteins of the
virus.
To determine whether the glycoproteins present in themem-branes of infected cells are also structural com-ponents ofthe
virus,
partially purified
virus wasprepared asdescribedabove. This viruswas
pre-pared from the tissue culture fluid and saline washingsof the sameinfectedcells whichyielded
the 2-P preparation and its electropherogram
shownin Fig. 3. Thechoice of extracellularfluid as a source of virus for purification is based on
electron microscopic observations (unpublished
data) that it yields consistently cleaner
prepara-tionsof virus than celllysates.Theelectrophoretic profiles in Fig.7showthedistribution of glucosa-mine and amino acids in the proteins of the solubilized virus preparation. A comparison of Fig. 3 and 7 shows that the glucosamine profiles are practically indistinguishable; the amino acid profiles differ in that several additional protein bandsare presentinvirus.Weconclude that, with
respect totheFstrain of herpessimplex(a type 1
virus), the electrophoretic profiles of the
glyco-proteins in the membranesand virion are
identi-cal.Glycoproteins similar inelectrophoretic
mo-bilitiesto those presentin the F virus have also
beenfound inpartially purified virus prepared by Olshevsky and Becker (Virology, in press). The
choice of the F strain for these studies may have been fortuitous, since studies of glycopro-teins made in cells infected with several other
variants (Keller,Spear,andRoizman,manuscript in preparation) have shown that purified virus contains glycoproteins inaddition to those
con-tained in the membranes.
DISCUSSION
Characteristicsof the membranes infraction2-P.
Biochemical
data furnished by Bosmann et al (2) showed that the membranes recovered by theirprocedure,
in a fraction equivalent to 2-P, aresmooth endoplasmic reticulum. Some of the
FIG.7. Electropherogramofproteins presentin par-tiallypurifiedherpesvirus. Theviruswaspreparedfrom the extracellularfluid ofcells incubated in medium containing 8H-glucosamineand14C-leucine, -isoleucine, and-valinefrom4 to 22hr andfrom6 to 22hr post-infection, respectively. Symbols: solid line, 3H-gluco-samine; dashed line,14C-leucine, -isoleucine,and-valine.
5, 1970 129
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.496.46.235.397.552.2] [image:7.496.252.443.452.583.2]SPEAR, KELLER, AND ROIZMAN
characteristics of the membranes in our fraction 2-P, which was studied in greatest detail, are as follows. (i) The membranes are free from ribo-somes, virus, and soluble proteins. (ii) Prelimi-nary chemical analyses performed by E. D.
Kieff indicate thatthey consist of 50% lipid and are free from sphingomyelins. (iii) The spectrum ofhost glycoproteins in the smooth membranes appears unaltered after infection. It should be
pointed out that the absence of infectious virus in the 2-P membrane band is particularly
sig-nificant with respect to the purity of the
mem-branepreparation. Specifically,onlyenvelopedor
partially enveloped virus is infectious (30). The
buoyant density ofinfectious virus is lower than
that of unenveloped virus. However, even
en-velopedvirusappearsmore dense than the mem-branes inband 2-P.
Nature of the proteins incorporated into the membranes. Inthispaper, we have demonstrated
that three to four proteins are synthesized after infection and are subsequently incorporated into
cellular membranes. The following points
con-cerning these proteins should be noted. (i) The glycoproteins incorporated into the membranes
migrateat the samerate as proteins rangingfrom 50,000 to 100,000 daltons in molecular weight (29). (ii) The same glycoproteins appear to be incorporated into all membrane fractions
ex-amined. (iii) In theprevious paperin thisseries,
we cited evidence that theproteins made 4 to 6
hr after infectionare specified bythe virus (29). To this listwe may add two findings. First, the glycoproteins made after infection appear to be structural components of the herpesvirion. Sec-ond,recentstudies (Keller,Spear, and Roizman, manuscriptinpreparation) showthatthe electro-phoretic profiles oftheglycoproteins synthesized in infected HEp-2 cells vary with the strain of herpesvirusused.
Significance of this work. Recent interest in
structure and biosynthesis ofmembranes of in-fected cellsstemsfromthree considerations. First,
in thecourse of infection,
all
viruses mustneces-sarilypassthroughatleastonecellular membrane
whenthey enter and exit from the cell. The struc-tureof cellmembranes isadeterminant of
infec-tion, and information concerning membranes is
essential for the understanding of the
mecha-nisms of entry and egress of the virus. Second,
nearly all viruses studied to date have been shown to affect the biosynthesis and structure of membranes. Theinteractionsofsome viruses with cell membranes are highly specific; these are
usually viruses containing lipid structural
con-stituents. With the apparent exception of
vac-cinia (6), viruses containing lipids acquire an
envelope from a membrane of the cell. In the
case of some viruses, viral proteins associated
withlipidandmakingup theenvelopehavebeen
identifiedandcharacterized (1, 11, 13,32, 33).It isthought that these viruses are enveloped
by
a membrane modified by prior insertion of viralproteins, butproofislackingsincepurified
mem-branes containing these proteins have not been
furnished.Third, cellsinfectedwithsome viruses
surviveinfectionbutexpressnewsurface
antigens
and exhibit altered behaviorwith respect to inter-action with other cells, the surfacesonwhichthey grow, and transplantability into a nonimmune
host.Someofthealtered propertiesofthe trans-formed cells arenecessarily mediated bythe sur-faceofthecells. At least in the caseofcells trans-formed by papovaviruses, thevirus-induced sur-faceantigens that have been identified have been
shown to result from a reorganization of the
plasma membrane whichresults inhostantigens coming accessible atthe surface(3, 12, 16, 18).
Inrelation to the basicproblems cited above,
the significance of our findings is as follows. (i)
The isolation from infected cells ofa membrane fraction containing only the glycoprotein constitu-entsof thevirion, coupled with the presence of the same glycoproteins in all membrane fractions,
suggests very strongly that the glycoproteins bind tomembranes and modifythembefore envelop-ment. The experimental data presented in this paper are inagreement with conclusions based on purely immunological grounds, namely, thatthe newantigenson theplasmamembraneof
herpes-virus-infected cells areidenticaltothoseon viral
envelopes (19, 21). (ii) Several herpesviruses are associated with tumors in man and animals. In one, the Burkittlymphoma of African children, it has been shown that tumor cells exhibit new
surface antigens, that there isagood correlation between antibodiestosurface
antigens
and to in-tracellular viral antigens (14, 15), and that re-gression ofthe tumor correlateswithanincreaseintiters ofantibody tosurface antigens (5).The
demonstration thatherpes simplex virus specifies glycoproteins which bind to membranes gives
credence to thehypothesis that the new surface antigensin these tumors arespecified bythevirus. (iii)To ourknowledge,this isthe firstisolation of cellular membranes which contain viral glyco-proteinsand are freefrom all otherprotein
con-stituentsof thevirus. Such amembrane
prepara-tionshouldbeusefulfor thestudy of membrane structure. Studies ofthe properties ofthe
glyco-proteins and of the membranes to which they bindarecontinuing.
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service grantCA08494 from the National CancerInstitute, American CancerSocietygrant E314E,andNational Science Foundation
130 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
PROTEINS SPECIFIED BY HERPES SIMPLEX VIRUS
grantGB 8242.P.G.S. is aPublic Health Servicepostdoctoral trainee(TO1HD00297-01). J.M.K. isafellowof theLeukemia Society.
Weacknowledgetheadvice of JeromeSchwartz and Robert Friis and the technical assistance of Patricia Wiedner in the preparation ofthe electron photomicrographs. A preliminary
accountofthispaper waspresentedatthemeetings of the Amer-ican SocietyforMicrobiologyinMayof1969.
LIERATURE CITED
1. Bikel, I., andP.H. Duesberg. 1969. Proteins of Newcastle disease virus andof theviralnucleocapsid. J. Virol. 4:388-393.
2. Bosmann, H. B.,A.Hagopian, and E.H.Eylar. 1968. Cellu-lar membranes: theisolation andcharacterization ofthe plasma and smooth membranes of HeLa cells. Arch. Bio-chem.Biophys. 128:51-69.
3. Burger, M.M. 1969. Adifferenceinthearchitecture of the surfacemembrane ofnormalandvirally transformedcells. Proc. Nat.Acad. Sci.U.S.A. 62:994-1001.
4. Choules, G. L., and B. H. Zimm. 1965. Anacrylamide gel
soluble in scintillation fluid: itsapplicationtoelectrophoresis
atneutral andlowpH.Anal.Biochem.13:336-344. 5. Clifford, P.,S. Singh.,J. Stjemsward, and G. Klein. 1967.
Long-termsurvival ofpatients with Burkitt's lymphoma:
anassessmentoftreatment andotherfactorswhichmay
re-latetosurvival.Cancer Res.27:2578-2615.
6. Dales, S.,and E. H.Mosbach. 1968.Vacciniaas amodelfor
membranebiogenesis. Virology 35:564-583.
7. Darlington, R. W., and L. H. Moss III. 1968.Herpesvirus envelopment.J.Virology2:48-55.
8.Eagle, H. 1959.Amino acid metabolism in mammaliancell
cultures.Science(Washington) 130:432-437.
9.Ejercito,P.M.,E. D.Kieff,and B.Roizman. 1968.
Charac-terization ofherpes simplex virus strains differingintheir effectonsocial behavior ofinfectedcells. J. Gen. Virol. 3:357-364.
10. Epstein, M. A.1962.Observationsonthemode of release of
herpesvirus from infectedHeLacells. J. CellBiol. 12:589-597.
11. Evans, M. J.,and D. W.Kingsbury.1969.Separationof
New-castledisease virusproteins bypolyacrylamidegel
electro-phoresis. Virology 37:597-604.
12.Hakomori,S.,C.Teather,andH. Andrews. 1968. Organiza-tional differenceof cell surface "hematoside" in normal and
virally transformedcells.Biochem.Biophys.Res. Commun.
33:563-568.
13. Haslam,E.A.,I.M. Cheyne,andD.0.White. 1969.The
structural proteins ofNewcastle disease virus. Virology 39:118-129.
14.Klein,G.,G.Pearson,G.Henle,W.Henle,V.Diehl,and J. C.
Niederman. 1968.Relation betweenEpstein-Barrviraland
cellmembrane immunofluorescence in Burkitt tumor cells.
II. Comparisonofcells andserafrompatientswithBurkitt's lymphoma and infectious mononucleosis. J. Exp. Med.
128:1021-1030.
15. Klein, G., G. Pearson,J.S. Nadkarmi, J. J. Nadkarmi,E.
Klein,G.Henle,W.Henle,and P.Clifford. 1968. Relation
betweenEpstein-Barr viral and cellmembrane
immuno-fluorescenceof Burkitt tumor cells. I.Dependenceofcell
membrane immunofluorescenceonpresence of EB virus.J.
Exp. Med. 128:1011-1020.
16.Meezan, E.,H. C.Wu,P.H.Black, and P. W. Robbins.
1969.Comparative studiesonthecarbohydrate-containing membrane components ofnormal and virus-transformed
mouse fibroblasts. II. Separation ofglycoproteins and
glycopeptides bysephadexchromatography. Biochemistry 8:2518-2524.
17.Morgan, C., S. A. Ellison, H. M. Rose, andD. H. Moore.
1954. Structure and development of virusesasobserved in
the electron microscope. I. Herpes simplex virus.J. Exp.
Med. 100:195-202.
18. Pollack, R. E., and M. M. Burger. 1969. Surface-specific characteristicsofacontact-inhibitedcellline containing the SV40 viralgenome.Proc.Nat. Acad.Sci.U.S.A. 62:1074-1076.
19. Roane, P. R.,Jr., and B. Roizman. 1964.Studies of the deter-minant antigens of viable cells. II. Demonstration of altered antigenic reactivity of HEp-2 cells infected with herpes sim-plex virus. Virology 22:1-8.
20.Roizman, B. 1962. Polykaryocytosis. Cold Spring Harbor Symp.Quant. Biol.27:327-342.
21. Roizman, B., and S. B. Spring. 1967. Alteration in immuno-logic specificity of cells infected with cytolytic viruses,
p.85-96. In J. J. Trentin (ed),Cross-reactingantigens and neoantigens, Williams & Wilkins Co.,Baltimore. 22. Rosan, R. C.,A. J.Nahmias, S.Kibrick,and J. A. Kerrigan.
1964. Studies inglycoprotein production:infection of
pri-marycultures of human amnionwith herpes simplex virus. Exp.Cell Res.36:611-624.
23. Schwartz,J., and B.Roizman. 1969. Concerning theegressof
herpessimplex virus from infected cells: electron and light microscopeobservations.Virology38:42-49.
24. Schwartz,J., and B. Roizman. 1969.Similaritiesand differences in thedevelopment of laboratorystrains and freshly isolated strains ofherpessimplex virus in HEp-2 cells. Electron mi-scopic studies. J. Virol. 4:879-889.
25. Shipkey, F. H., R. B. Erlandsen, R. B. Bailey, V. J.Babcock, and C. M. Southam. 1967.Virusbiographies. II. Growth of herpes simplex virus in tissue culture. Exp. Mol. Pathol. 6:39-67.
26. Siegert,R., and D. Falke. 1966. Elektronenmikroskopishe Untersuchungen uber dieEntruicklung des Herpes Virus hominus inKulturzellen.Arch. Ges.Virusforsch. 19:230-246.
27. Siminoff,P., and M. G. Menefee. 1966. Normal and 5-bromo-deoxyuridine-inhibiteddevelopment of herpes simplex virus. Anelectronmicroscopestudy.Exp. Cell Res.44:241-255. 28. Spear, P. G., and B. Roizman. 1968. Animprovedprocedure
for Hs and C4 counting in acrylamide gels witha non-aqueousscintillationsystem.Anal. Biochem. 26:197-200. 29. Spear, P. G., and B. Roizman. 1968. Theproteins specifiedby
herpes simplex virus. I.Time of synthesis, transfer into nuclei, andproperties of proteins made in productively in-fectedcells.Virology 36:545-555.
30. Spring, S. B., and B. Roizman. 1968. Herpes simplex virus products inproductive and abortive infection. III. Differ-entiation of infectious virus derived from nucleusand
cyto-plasm withrespect tostability and size. J. Virol. 2:979-985. 31.Spring, S. B., B. Roizman, and J. Schwartz. 1968. Herpes
simplex virusproductsinproductiveandabortive infection.
II.Electronmicroscopic andimmunological evidence for failure of virusenvelopmentas a causeofabortiveinfection. J. Virol. 2:384-392.
32. Strauss, J. H.,Jr., B.W.Burge, and J. E. Darnell. 1969. Sind-bisvirus infection ofchick andhamstercells;synthesisof virus-specific proteins. Virology 37:367-376.
33. Strauss, J.H., Jr.,B.W.Burge,and E. R. Pfefferkorn. 1968.
Identification of membraneprotein and"core" protein of Sindbis virus.Proc. Nat. Acad.Sci.U.S.A.50:533-537.
34. Summers,D.F.,J. V.Maizel, Jr.,andJ. E.Daruell,Jr.1965.
Evidenceforvirus-specific noncapsidproteinsin poliovirus-infectedHeLa cells. Proc. Nat. Acad.Sci.U.S.A.
54:505-513.
VOL. 5, 1970