Copyright 0 1993 by the Genetics Society of America
The Drosophila Sex Determination Gene
snf
Is Utilized for the
Establishment
of the Female-Specific Splicing Pattern
of Sex-lethal
Elizabeth Brown Albrecht and Helen K. Salz
Department of Genetics, Case Western Reserve University, Cleveland, Ohio 44106-4955
Manuscript received January 1 1 , 1993 Accepted for publication March 17, 1993
ABSTRACT
The Drosophila snf gene is a positive regulator of the sex determination gene Sex-lethal in both the germline and the soma. Its role in the soma is only evident when the probability of Sex-lethal activation has been reduced. For instance, in an otherwise wild-type background, females homozygous for a weak snf mutation produce both male and female progeny; however, when mated to males hemizygous for a null allele of Sex-lethal, they produce only male progeny. We demonstrate that the lack of female progeny is due to aberrant Sex-lethal regulation in late embryogenesis. In these mutant embryos, there is little accumulation of the late female-specific spliced RNAs and proteins. In contrast, in early embryogenesis, Sex-lethal regulation is not affected. The accumulation of both the early Sex-lethal transcripts and proteins is normal. These results suggest that the wild-type product of snf plays an important role in establishing the female-specific RNA splicing pattern of Sex-lethal. Whether snf influences the female-specific splice site choice directly or indirectly remains to be determined.
I
N Drosophila melanogaster, the decision between male and female development is regulated by the binary switch gene Sex-lethal, (Sxl; recently reviewed in BELOTE 1992). Normal female development re- quires that Sxl be active in all tissues throughout development, whereas normal male development re- quires that Sxl remain inactive. Because Sxl is also required for the vital process of dosage compensation, the lack of SxZ expression in females results not only in sex transformations but also in sex-specific lethality. Recent molecular studies have shown that female- specific activation of Sxl is a complex process. In the soma, the initial expression of Sxl is under female- specific transcriptional regulation (KEYES, CLINE and SCHEDL 1992). Expression from a female-specific pro- moter generates a unique set of early RNAs which are only present very early in embryogenesis (SALZ et al.1989; KEYES, CLINE and SCHEDL 1992). By gastrula- tion and throughout adulthood, the female-specific promoter is no longer utilized. Instead, expression from a non-sex-specific promoter generates a new set of transcripts (the late transcripts) which are differ- entially spliced in males and females (BELL et a l . 1988; SALZ et al. 1989; SAMUELS, SCHEDL and CLINE 1991). Once Sxl activity is established, it maintains its own expression by establishing an autoregulatory feedback loop (CLINE 1984; BELL et al. 1991). Consequently, Sxl expression remains female-specific throughout de- velopment because only the female-specific transcripts encode functional proteins (BELL et al. 1988; BOPP et
a l . 1991; SAMUELS, SCHEDL and CLINE 1991). T h e predicted protein products of both the early and the late female-specific RNAs are highly similar to each
Genetics 134: 801-807 uuly, 1993)
other (BELL et al. 1988; SAMUELS, SCHEDL and CLINE 1991 ; KEYES, CLINE and SCHEDL 1992). T h e finding that these proteins share homology with a family of RNA binding proteins suggests that they direct the female-specific splicing pattern of Sxl by directly in- teracting with the RNAs (BELL et al. 1988; INOUE et
a l . 1990; SAMUELS, SCHEDL and CLINE 1991; KEYES, CLINE and SCHEDL 1992).
In this paper, we investigate the mechanism by which snfregulates Sxl. Females homozygous for the original snfmutation, snf 16*’, are sterile because the
germline components of the ovary do not differentiate properly (OLIVER, PERRIMON and MAHOWALD 1988; STEINMANN-ZWICKY 1988; SALZ 1992). Instead, the germarial cells continue to divide resulting in the formation of “ovarian tumors.” Two lines of evidence suggest that snf is required to activate Sxl in the germline. First, the snf mutant phenotype is sup- pressed by either of two constitutive alleles of Sxl
(STEINMANN-ZWICKY 1988; SALZ 1992). Thus unre-
gulated expression of Sxl can bypass the requirement for snf + function. Second, the Sxl transcription pat-
tern is abnormal in snf mutant females (SALZ 1992). T h e two germline-dependent RNAs are missing whereas the remaining female-specific RNAs appear to be processed normally. Since the regulation of Sxl is not well characterized in the germline, it is difficult to assess where in the process of Sxl regulation snf is likely to function (SALZ, CLINE and SCHEDL 1987; SALZ et al. 1989; BOPP et a l . 1991; SAMUELS, SCHEDL and CLINE 199 1).
examine its role in the embryo. In contrast to snf's
role in the germline, its role in the somatic sex deter- mination is only evident when the probability of Sxl activation has otherwise been reduced (CLINE 1988;
OLIVER, PERRIMON a n d MAHOWALD 1988; STEIN-
MANN-ZWICKY 1988; SALZ 1992). Here, we investigate the basis for the strongest female-specific lethal inter- action, using the snfeBH allele (SALZ 1992). In this case, t h e female-specific lethality is solely d u e t o t h e lack of maternal snf function. We further demonstrate, by examining both the RNA and protein expression pat- tern of Sxl in the mutant embryos, that Sxl is not stably activated. Although the early female-specific
transcriptional pattern of Sxl and the early SXL pro- tein expression appear normal, there is little or n o accumulation of the female-specific spliced form of the late transcripts, and consequently little or n o SXL protein later in embryogenesis. These data indicate that this snf mutation blocks the transition between the transcriptional and post-transcriptional modes of Sxl regulation in animals that are heterozygous for S x l . Since this snf allele appears to be a weak loss of function mutation, we reason that the wild type func- tion of snf is t o establish t h e female-specific splicing pattern of S x l . Whether snf is directly involved in establishing the female-specific splicing pattern o r whether it indirectly influences the female-specific
splice site choice remains to be determined. Moreover, it remains to be determined whether snf is essential for the establishment of the female-specific splicing pattern or instead is a "facilitator" whose function is only critical when the probability of Sxl activation has otherwise been reduced.
MATERIALS AND METHODS
Fly stocks and culture conditions: D f l ) D E B 4 D , snfeBH and Dp(l;2)4FRDup are described in SALZ (1992). The re- maining mutations and balancer chromosomes are described in LINDSLEY and ZIMM (1992). snfwas initially identified in a screen for female-sterile mutations and namedfs(l)l621 (CANS, AUDIT and MASSON 1975). Since then it has been renamed sans-jille ( s n f ) (OLIVER, PERRIMON and MAHOW- ALD 1988) and Liz (STEINMANN-ZWICKY 1988). Although we use the name snf, the locus is still identified by others as
fs(1)1621 and Liz in the literature.
Flies were grown at room temperature, which ranged from 22" to 25", on a standard cornmeal, yeast, molasses, agar medium.
Western blot analysis: Staged embryos were collected and frozen on dry ice and stored at -80" until ready for use. Protein extracts were prepared in a similar fashion to the procedure described in DRIEVER and NUSSLEIN- VOLHARD ( 1 988). Approximately 200 pg eggs still in their chorions were homogenized while thawing in -300 /AI of 2
X loading buffer containing 8 M urea and 4% sodium dodecyl sulfate. After the samples were boiled for 5 min they were centrifuged for 1 min at 4000 X g to spin down the debris. A 1 5 - ~ 1 aliquot was applied to each slot of a 12% polyacrylamide gel. Western blot analysis was performed as described in BOPP et al. (1 99 1). The blot was incubated with
TABLE 1
Lack of maternal snf function is responsible for the lethal synergistic interaction between snf'" and Sxl
Relative via
hilit I of snf+/+Sxl; Relative via bility o f
Cross with respect to snf females (n) females (n) males n Genotype of mother snf+)+Sxl Dp(snf')/+ Keference
A snf/snf; Dp(snf')/+ 89.4% (84) 98.9% (93) 188
B snflsnf <0.4% (0) <0.4% (0) 549
C snf/snf <0. 1 % (0) N E 769
Relative viability is assessed by comparing the number ( n ) of females of a given genotype to the number of males recovered from the same cross. For crosses A and B, the number of females of each genotype is expected to be half the number of males recovered.
For cross C:, the number of females is expected to be equal to the number of males. NE, not expected. T h e full genotypes of the crosses were as follows:
A . ut snf'"'//w myXH; Dp(l;2)4FRDup, w+ snf+f+9P X y U I S x l f ' rl
66.
B. w snyxHfw snf'"" 99 X y w S x l s ' c t ; Dp(l;P)4FRDup, UI+ snf'f+
66.
C . uj snf""'/w myuH OP X y w Sxi'' ct 66.
an anti-SXL monoclonal antibody (BOPP et al. 1991) and visualized by use of an alkaline phosphatase conjugated anti- mouse antibody (Cappel). As a loading control the blot was stripped and incubated with an anti-a-tubulin antibody (Sigma) and visualized with an alkaline phosphatase-conju- gated anti-mouse antibody (Cappel).
Immunolocalization in whole-mount embryos: After standard fixation, the embryos were stained for SxZ expres- sion with an anti-SXL monoclonal antibody (BOPP et al. 1991). The primary antibody was visualized by standard immunohistochemical procedures using an alkaline phos- phatase conjugated anti-mouse antibody (Cappel).
Northern blot analysis: Poly(A+) R N A was isolated as previously described (SALZ et al. 1989). Northern blot analy- sis was performed as in SALZ et al. (1 989) with the following modifications. The R N A was transferred to Nytran (Schleicher and Schuell) and hybridizations were done in CHURCH'S (1984) buffer. The Northern blot was probed with the S x l cDNAcF1 described in BELL et al. (1988).
RESULTS
Lack of maternal snf function is responsible for the lethal synergistic interaction between snf and
Sxlf': Females homozygous for the weak snfeBH allele do not exhibit the female sterility associated with the stronger snf mutation. snf e8H females are homo- zygous fertile and have equal numbers of male and female progeny (data not shown). However, like
snf 1 6 2 1 , snf e*H exhibits a female-lethal synergistic in-
teraction with Sxl mutations (SALZ 1992). When fe- males homozygous for snf e8H are crossed to a male carrying a null allele of S x l , n o females are recovered (Table 1, cross C). In the table, the null allele used is a n EMS induced mutation, S x l f l , which appears not to interfere with transcription of the locus (H. K . SALZ,
Sxl Female-Splicing Utilizes snf 803
T h e data presented in Table 1 further demonstrate that the female-specific lethality resulting from the snfeBH-Sxl interaction is due to the reduction of ma- ternal snf activity. T h e presence of a duplication for snf + in the mother is sufficient to allow all daughters
to survive, whether they carry the duplication o r not (cross A). However, the presence of the duplication in the daughters alone is not sufficient for survival (cross B).
snf interacts with Sxl f 9 but not Sxl fLs: To begin to characterize the nature of the interaction between snf and S x l , we tested for an interaction between snf with two complementing partial loss of function Sxl alleles. S x l p is primarily defective in the early Sxl regulation (MAINE et al. 1985; CLINE 1986). SxlfL", on the other hand, is defective in a later function re- quired for maintenance and/or expression (SANCHEZ and NOTHICER 1982; MAINE et al. 1985; CLINE 1986). Female progeny heterozygous for S x l p from snf mothers are recovered less frequently than expected (65% relative viability). In contrast, in the same mu- tant background, female progeny heterozygous for S x l f l s are fully viable. T h e lack of interaction is not unexpected, however, in light of the fact that the snf '8H-Sxl interaction is strictly maternal and S ~ l f l , ~ appears to affect Sxl function only later in develop- ment.
Sxl is not stably activated in a snf mutant back- ground: T o begin to examine the basis of this female- specific lethal synergistic interaction we examined the phenotype of the mutant embryos. Embryos were collected from females homozygous for snf crossed to males hemizygous for S x l f ' . We will refer to these mutant embryos as embryos from the "snf mutant background." Because this mutant effect is only ob- served in females with a single copy of S x l + , the control embryos are collected from females homozy- gous for snf c8H crossed to wild-type males.
All embryos in the snf mutant background enter gastrulation and develop normally until stage 13 (- 1 1 hr). After this stage some of the embryos stop devel- oping. By stage 16 (-1 7 hr), 46% (1 75/382) of the embryos had degenerated as judged by their lack of internal structures and granular appearance. Simi- larly, in crosses that produce 25% Sxl homozygous females, embryogenesis proceeds normally through gastrulation. By stage 16, however, about one fourth of the embryos have begun to degenerate. T h e simi- larity in phenotypes suggests that the embryos in the snf mutant background are dying because they cannot properly activate or stably maintain Sxl function.
Two lines of evidence confirm that this hypothesis is correct. First, the presence of a constitutive allele of Sxl in the embryo can suppress the female-lethal synergistic interaction. T h e data in Table 2 demon- strate that the constitutive expression of Sxl is suffi-
TABLE 2
Suppression of the lethal synergistic interaction between snf"
and Sxl
Relative viability Relative viability
of snf+ f+ Sxl of snf SxlMf+ Sxl Reference
Maternal genotype females (n) females (n) males (n)
snf "" sxlM4/snf 0 100% (254) 202
Relative viability is determined by comparing the number (n) of females of a given genotype to the total number w snf males recovered from the cross y snf "" cm SxlM4/w
myBH
99 X y w cm Sxlf' c t 66. Since the recombinant and non recombinant females could not be distinguished, ally cm females were presumed to bey snf''"
cm Sxl"/y w cm Sxlf' ct. Two hundred and twenty of these females were recovered. Thirty-four w females were also recovered. This phenotype could be of the genotype w snf""/y w cm S x l f c t or could be the result of a recombination event in the w SxlM4 interval in the mother and thus be of the genotype wmysH
cm SxlM4/w Sxlf'ct. By individually test crossing each female, we determined that all the w females were of the genotype w snf'" cm SxlM4/w S x l f ' ct.
Each female had about half as many male progeny as daughters, suggesting that they all carried the male lethal SxlM4 allele. T h e recovery of these 34 recombinant females is about what we would have ex ected based on the recombination frequency between w and Sxl E 4 . To calculate the expected recombination frequency
between w and Sxl", the total number of w+ cm+ males was scored. cm is used as a marker for Sxl since cm is closely linked to S x l . Because SxlM4 is male lethal, only the w+ cm+ recombinants are recovered in males. Forty-two w+ cm+ males were recovered.
cient to bypass the lethal effect in those female prog- eny that are heterozygous for SxlM4. On the other hand, their sisters which are not heterozygous for
SxlM4 are not recovered. Similar results were also obtained when another constitutive allele of S x l , Sxl", was tested (data not shown).
Second, we demonstrate that the SXL protein is absent in embryos from the snfmutant background. Embryos (8-1 1 hr) of the snf mutant background were stained with an antibody to SXL (Figure 1). Normally, by 8 hr in development, Sxl activation is complete and SXL protein can easily be detected in one half of the embryos (Figure 1A). In contrast to control embryos, no SXL protein was observed in embryos from the snf mutant background (Figure 1 B).
Alteration of the Sxl transcription pattern in the
A
B
C
D
4 . 4 0 4.29
3.60
3 . 3 9 Embryo 3.1-
Embryo 1.7-
I
1
FIGURE I.-SXL protein expression i n control and snf mutant backgrounds. Expression of S X L protein detected in whole mount embryos using a monoclonal antibody against all the SXL proteins. (A) Typical field of embryos in the control background from the control cross: snf ""/snf females X Sxl+ males fixed 8-1 1 hr after egg deposition, illustrating that Sxl is expressed in only half the embryos. Similar results are obtained when embryos from a wild- type stock are used (data not shown). (B) Typical field of embryos in the snf mutant background from the cross: snf ""/snf
''"
females X Sxl'' males fixed 8-1 1 hr after egg deposition, illustrating the lack of Sxl expression in all embryos.experimental lanes probably due to the inherent in- accuracies in timed collections of embryos.
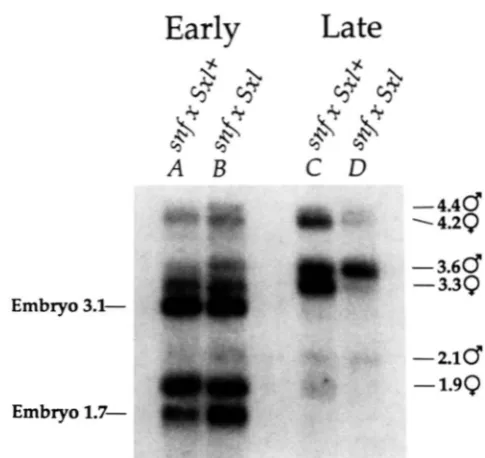
In sharp contrast, the transcription pattern detected in older embryos is markedly different in the mutant and control backgrounds. In the control embryos, t w o distinct sets of transcripts are detected, the 4.2- and 3.3-kb late female-specific RNAs and the 4.4- and 3.6- kb late male-specific RNAs. In contrast, in the snf
mutant background, the predominant RNAs are
male-specific and only a low level of the late female- specific RNAs are detectable.
To confirm that there is Sxl transcription in all 8-
FIGURE 2.-The early and late Sxl transcription pattern in con- trol and snfmutant backgrounds. Northern blot of 5 pg of poly(A+) RNA from embryos in the control background from the control cross: snf"/snf
''"
females X Sxl + males (lanes A and C) or the snfmutant background from the cross: snf'nH/snf'nH females X Sxl'' males (lanes B and D) hybridized with the probe Sxl cDNAcFl which detects all classes of Sxl RNAs. See text for a description of the different classes of Sxl RNAs. (A) Early embryos (0-5 hr) in the control background. (B) Early embryos (0-5 hr) in the snf mutant background. (C) Late embryos (8-1 1 hr) in the control background. (D) Late embryos (8-1 1 hr) in the snf mutant background. In contrast to our data, SALZ et al. (1989) observed that a 1.9-kb female-specific R N A is abundant until about 14-17 hr in develop ment. However, in similar experiments, KEYES. CLINE and SCHEDL (1992) find that the 1.9-kb R N A is barely detectable already by 5-
8 hr in development. Although it is not clear why these differences in abundance are observed, it is unlikely that they are relevant to the lethal synergistic interaction because these differences are ob- served in both the mutant and control backgrounds.
1 1-hr embryos, whole mount embryos were hybrid- ized, in situ, with a Sxl probe that detects all the different classes of Sxl RNAs (data not shown). T h e detection of signal in all the embryos suggests that transcription from the late Sxl promoter occurs in all of the embryos. This data together with the results of the RNA analysis suggest that the majority of late pre- mRNAs in both male and female embryos are proc- essed in the male mode.
Sxl Female-Splicing Utilizes snf 805
Early
1
2
180kd-
84
kd-
48.5
kd-
26.6
kd-1
anti-a-Tubulin-
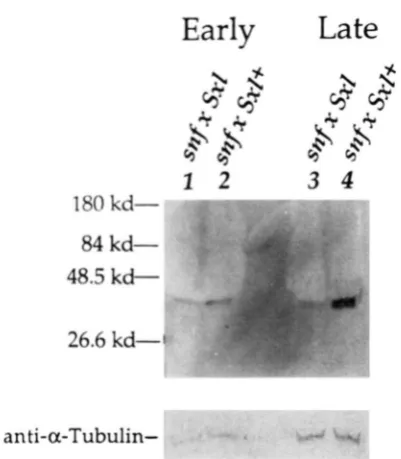
FIGURE 3.-The early and late SXL protein expression pattern in control and snf mutant backgrounds. Western blot o f protein extracts from either embryos in the control background from the control cross: snf ‘RH/snf females X Sxl + males (lanes 2 and 4) or
embryos in the snfmutant background from the cross: snf ‘RH/snf
females X S x l f ’ males (lanes 1 and 3) detected with a monoclonal antibody against SXL. (1) Early (0-5-hr embryos) in the snfmutant background: (2) early (0-5-hr embryos) in the control background; (3) late (8-1 I-hr embryos) in the snfmutant background; (4) late (8-1 I-hr embryos) in the control background. As a loading control, the blot was subsequently incubated with anti-a-tubulin to detect the 55-kD protein. A similar pattern o f SXL protein expression, as the control pattern, is obtained when embryos from a wild-type stock are used (data not shown).
ground by Western blot analysis (Figure 3). T h e early SXL proteins are as easily detectable in mutant em- bryos as in control embryos, suggesting that the early Sxl transcripts seen in Figure 3 are translated (Figure 3, lanes 1 and 2). Later in development, there is significantly less SXL protein compared to control embryos (Figure 3, lanes 3 and 4). Hence, the levels of SXL protein detected parallel the levels of the female-specific Sxl transcripts in the snfmutant back- ground.
DISCUSSION
snf’s role i n somatic sex determination as a positive regulator of Sxl was first suggested by OLIVER, PER-
ZWICKY ( 1 988) on t h e basis of a female-lethal syner- gistic interaction between snfand S x l . Further elabo- ration of snf’s role in the complex process of Sxl activation has been complicated by the lack of a snf
null allele; however, the isolation of additional snf
mutations by conventional genetic approaches has proven to be very difficult (SALZ 1992). Only one new allele was isolated in a screen of -25,000 chromo- somes using three different types of mutagens (SALZ
RIMON alld MAHOWALD (1988) and STEINMANN-
1992). In this report we take advantage of a this new weak allele of snJ snf “If, to examine further snf’s role in the embryo. This type of analysis parallels the analysis of daughterless and J(2)d (CRONMILLER and
CLINE 1987; CLINE 1989; GRANADINO, CAMPUZANO
and SANCHEZ 1990; GRANADINO, SAN
JUAN
and SAN-CHEZ 199 1 ; GRANADINO et al. 1 992). Since these genes have additional non-sex-specific functions, the analysis of weak alleles was essential to their identification as positive regulators of S x l .
Our results demonstrate that females heterozygous for Sxl from mothers homozygous for snfCRH do not survive because they fail to establish the autoregula- tory loop even though they appear to correctly initiate Sxl in early embryogenesis. In early embryogenesis, the pattern of accumulation of the early transcripts and the early proteins is similar in both mutant and control embryos; not only do the embryo-specific tran- scripts appear to be of the appropriate size, but the early presence of SXL protein suggests that the em- bryo-specific transcripts are also appropriately spliced. Furthermore, in both mutant and control embryos the early RNAs are no longer present after gastrula- tion, suggesting that the female-specific Sxl promoter is correctly down regulated. During late embryogen- esis, Sxl transcription occurs in all the embryos, sug- gesting that the non-sex-specific promoter has been activated. However, the mutant embryos show a lack of accumulation of the late female-specific spliced RNAs indicating that the switch to the autoregulatory mode of regulation is blocked.
Since snf
’*”
appears to be a partial loss of function mutation, these data strongly suggest that snf is a maternal component whose wild-type function is to establish the autoregulatory feedback loop, a process that is regulated by alternative RNA splicing (Figure4). Whether snfdirectly influences the female-specific splice site choice or whether it prevents the transition to the autoregulatory mode of regulation in a more indirect manner remains to be determined. For in- stance, it is possible that snf indirectly influences the establishment of the female-specific splicing pattern by reducing the amount and/or activity state of the early Sxl proteins.
INITIATION
TRANSITION
FEEDBACK
LOOP
ACTIVATION
AND
MAINTENANCE
Q
Sxl
ON
2X.:2A daughterless
\
d o n-
Fi p.early SXL protein
-
proteinlate SXL
0
SXI
OFF
1X:2A daughterless
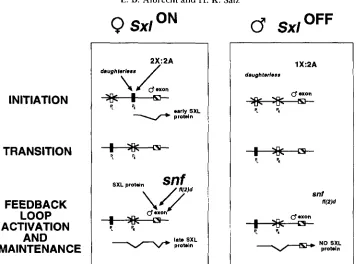
FIGURE 4.-Model for regulation of S x l . Both genetic and molecular experiments from several laboratories have suggested a detailed model for the sex-specific regulation of Sxl (see text for references). The regulation of S x l activity can be divided into a multistep process. Initiation: the early, female-specific promoter (PE) directs the transcription of an early class of Sxl RNAs in response to an X A ratio of 2:2. In addition to the components of the X A ratio, transcription from the female-specific promoter requires the presence of the maternally provided daughterless ( d a ) gene product. Transition: after cellular blastoderm, PE becomes inactive and the late non-sex-specific promoter
(PL) is activated. Feedback loop activation and maintenance: initially, the early S X L protein directs the female-specific splicing of the late Sxl transcripts from PL resulting in the production of the late S X L proteins. Thereafter, the presence of the late S X L proteins ensures that the primary transcripts will continue to be spliced in the female protein encoding mode. In males, on the other hand, the early S X L proteins are absent, and the primary transcripts from PL are therefore spliced in the male mode, resulting in no functional S X L proteins.
(CLINE 1986). However, the male lethality can be suppressed by ~nf’~’’ (SALZ 1992). Taken together, these results suggest that the establishment of the autoregulatory loop can be sensitized to variations in snf activity. Perhaps snf interacts with the early Sxl protein, influencing the efficiency with which it directs female-specific splicing of the late Sxl RNAs.
snfalso has a similar zygotic function which is only detectable using the stronger snf 16” mutant allele (STEINMANN-ZWICKY 1988; OLIVER, PERRIMON and MAHOWALD 1988; SALZ 1992). Perhaps snf is also required throughout development for maintenance of Sxl activity. Indeed, the observation that the strongest snfmutation, snf 16*’, can suppress the male lethality associated with Sxl misexpression in Sxl” males could suggest that snf is required throughout development for the maintenance of Sxl activity (STEINMANN- ZWICKY 1988; SALZ 1992). Mutations inP(2)d, a gene required for maintenance of Sxl activity, also suppress the male lethality associated with Sxl” (GRANADINO et al. 1992).
Although our results clearly demonstrate that snf
participates in the establishment of the Sxl autoregu- latory loop, the observation that a female-lethal phe- notype is only observed in animals with reduced S x l + copy number raises the question of whether snf is
essential for this process. Indeed, the two extant snf
mutations have no effect on viability by themselves, further suggesting that snf is a “facilitator” whose product only becomes essential when the probability of activation of Sxl is reduced. On the other hand, since the existing alleles of snfare not null mutations, one cannot exclude the possibility that snf has addi- tional functions in sex determination and/or other processes.
We thank C. CRONMILLER and D. BOPP for antibodies and C. CRONMILLER, I. GREENWALD, D. SETZER, A. RIEDL and members of the SALZ, MORGAN and SEDENSKY laboratories for helpful dis- cussion and comments on the manuscript. This work was supported by a grant from the National Science Foundation (to H.K.S.), a Young Investigators Award from the Mathers Charitable Founda- tion (to H.K.S.) and a National Research Service Award 5T32 GM08056 (to E.B.A.).
LITERATURE CITED
BELL, L. R., E. M. MAINE, P. SCHEDL and T. W. CLINE, 1988 Sex-
lethal, a Drosophila sex determination switch gene, exhibits
sex-specific RNA splicing and sequence similarity to RNA binding proteins. Cell 55: 1037-1046.
BELL, L. R., J. 1. HORABIN, P. SCHEDL and T . W. CLINE, 199 1 Positive autoregulation of Sex-lethal by alternative splic- ing maintains the female determined state in Drosophila. Cell
BELOTE, J. M., 1992 Sex determination in Drosophila melanogaster:
Sxl Female-Splicing Utilizes snf 807
from the X:A ratio to doublesex. Semin. Dev. Biol. 3: 319-330.
BOPP, D., L. R. BELL, T. W . CLINE and P. SCHEDL, 1991
Developmental distribution of female-specific Sex-lethal pro- teins in Drosophila melanogaster. Genes Dev. 5: 403-41 5.
CHURCH, G. M., 1984 Genomic sequencing. Proc. Natl. Acad. Sci.
CLINE, T. W., 1984 Autoregulatory functioning of a Drosophila gene product that establishes and maintains the sexually deter- mined state. Genetics 107: 231-277.
CLINE, T. W., 1986 A female specific lethal lesion in an X-linked positive regulator of the Drosophila sex determination gene,
Sex-lethal. Genetics 113: 641-663.
CLINE, T. W., 1988 Evidence that sisterless-a and sisterless-b are two of several discrete “numerator elements” of the X/A sex determination signal in Drosophila that switch Sxl between two alternative stable expression states. Genetics 1 1 9 829-862.
CLINE, T. W., 1989 T h e affairs of daughterlessand the promiscuity of developmental regulators. Cell 59: 231-234.
CRONMILLER, C., and T. W. CLINE, 1987 T h e Drosophila sex determination gene daughterless has different functions in the germline vs the soma. Cell 48: 479-487.
DRIEVER, W., and C. NUSSLEIN-VOLHARD, 1988 T h e bicoid pro- tein determines position in the Drosophila embryo in a concen- tration-dependent manner. Cell 54: 95-104.
CANS, M., C. AUDIT and M. MASSON, 1975 Isolation and charac- terization of sex-linked female-sterile mutants in Drosophila
melanogaster. Genetics 81: 683-704.
GRANADINO, B., S. CAMPUZANO and L. SANCHEZ, 1990 T h e Dro- sophila melanogasterfl(2)d gene is needed for the female-specific splicing of Sex-lethal RNA. EMBO J. 9: 2597-2602.
GRANADINO, B., A. B. SAN JUAN and L. SANCHEZ, 1991 T h e g e n e fl(2)d is required for various Sxl-controlled processes in Dro- sophila females. Wilhelm Roux’s Arch. Dev. Biol. 2 0 0 172- 176.
GRANADINO, B., A. B. SAN JUAN, P. SANTAMAR~A and L. SANCHEZ,
1992 Evidence of a dual function infl(2)d, a gene needed for
Sex-lethal expression in Drosophila melanogaster. Genetics 130:
USA 81: 1991-1995.
597-612.
INOUE, K., K. HOSHIJIMA, H. SAKAMOTO and Y. SHIMURA,
1990 Binding of the Drosophila Sex-lethal gene product to the alternative splice site of transformer primary transcript. Nature 3 4 4 461-463.
KEYES, L. N., T . W . CLINE and P. SCHEDL, 1992 T h e primary sex-determination signal of Drosophila acts at the level of transcription. Cell 68: 933-943.
LINDSLEY, D., and G. G. ZIMM, 1992 The Genome of Drosophila
melanogaster. Academic Press, New York.
MAINE, E. M., H. K. SALZ, P. SCHEDL and T. W. CLINE, 1985 Sex-
lethal, a link between sex determination and sexual differentia- tion in Drosophila melanogaster. Cold Spring Harbor Symp. Quant. Biol. 50: 595-604.
OLIVER, B., N. PERRIMON and A. P. MAHOWALD, 1988 Genetic evidence that the sans-flle locus is involved in Drosophila sex determination. Genetics 120: 159-1 7 1 .
SALZ, H. K., 1992 T h e genetic analysis of s n f : a Drosophila sex determination gene required for activation of Sex-lethal in both the germline and the soma. Genetics 130: 547-554.
SALZ, H. K., T . W. CLINE and P. SCHEDL, 1987 Functional changes associated with structural alterations induced by mo- bilization of a P element inserted in the Sex-lethal gene of Drosophila. Genetics 117: 221-23 1 .
SALZ, H. K., E. M. MAINE, L. N. KEYES, M. E. SAMUELS, T. W. CLINE and P. SCHEDL, 1989 T h e Drosophila sex determina- tion gene, Sex-lethal, has stage, tissue and sex-specific RNAs suggesting multiple modes of regulation. Genes Dev. 3: 708-
7 19.
SAMUELS, M. E., P. SCHEDL and T . W. CLINE, 1991 T h e complex set of late transcripts from the Drosophila sex determination gene Sex-lethal encodes multiple related polypeptides. Mol. Cell. Biol. 11: 3584-3602.
SANCHEZ, L., and R. NOTHIGER, 1982 Clonal analysis ofsex-lethal, a gene needed for female sexual development in Drosophila melanogaster. Wilhelm Roux’s Arch. Dev. Biol. 191: 21 1-214.
STEINMANN-ZWICKY, M., 1988 Sex determination in Drosophila: the X chromosomal gene liz is required for Sx1 activity. EMBO J. 7: 3889-3898.