EFFECTS OF POPULATION SIZE AND SELECTION INTENSITY ON SHORT-TERM RESPONSE TO SELECTION FOR
POSTWEANING GAIN I N MICE1
J. P. HANRAHANZ, E. J. EISEN A N D J. E. LEGATES
Department of Animal Science, North Carolina State Uniuersity, Raleigh, N . C.
Manuscript received August 21, 1972 Revised copy received November 1, 1972
Transmitted by ROBERT W. ALLARD
ABSTRACT
The effects of population size and selection intensity on the mean response was examined after 14 generations of within full-sib family selection for post- weaning gain in mice. Population sizes of 1, 2, 4, 8 and 16 pair matings were each evaluated at selection intensities of 100% (control), 50% and 25% in a replicated experiment. Selection response per generation increased as selection intensity increased. Selection response and realized heritability tended to in- crease with increasing population size. Replicate variability in realized herit- ability was large at population sizes of 1 , 2 and 4 pairs, Genetic drift was impli- cated as the primary factor causing the reduced response and lowered repeatability at the smaller population sizes. Lines with intended effective population sizes of 62 yielded larger selection responses per unit selection differential than lines with effective population sizes of 30 or less.
T H E usefulness of selection theory is measured by its ability to predict accu- rately the outcome of selection studies. Consequences of artificial selection based on infinite population size have been described adequately
(FALCONER
1960). In actuality selection for a quantitative character often is conducted with relatively small effective population sizes. I n such cases genetic drift may lead to fixation of a n allele even though a more desirable one is present in the popula- tion (ROBERTSON 1960). Thus, the cumulative effects of selection and genetic drift over several generations may yield a mean response less than that predicted from selection alone. The influence of drift on selection has been demonstrated for traits controlled by single genes (KERR and WRIGHT 1954a,b) but has received little experimental consideration for quantitative characters. A smaller effective population size also results in larger variance in gene frequency among replicate populations (WRIGHT 1938). Although it has been recognized that knowledge of the repeatability of selection response is necessary to strengthen general conclu- sions on the nature of selection response (FALCONER 1953), there is a paucity of experimental data dealing with this problem. An additional area which meritsPaper No. 3853 of the Journal Series of the Nonh Carolina State University hgriculiural Experiment Station, Raleigh, North Carolina. Supported in part by Public Health Service research grant GM1154F from the National Institutes of Health.
Present address: The Agricultural Institute, Creagh, Ballinrobe, County Mayo, Ireland.
514 J . P . H A N R A H A N , E. J . E I S E N A N D J . E . LEGATES
experimental investigation is the influence of selection intensity on rate of re- sponse. DEMPSTER (1955) and ROBERTSON (1960) have shown that for a fixed number of individuals measured the largest cumulative genetic advance is ex- pected when one-half of the population is selected each generation.
MATERIALS A N D M E T H O D S
The present experiment was designed to examine the effects of population size and selection intensity on short-term response and repeatability of response to selection for increased post- weaning gain (6- minus 3-week body weight) in mice.
Experimental design: Mice used t o initiate the study were sampled at random from an out- bred ICR albino population described by HANRAHAN and EISEN (1972). The experimental design and line designations are shown in Table 1. Population sizes used were 1 (full-sib mating), 2, 4, 8 and 16 pairs of parents. For each population size there were selection intensities of 25% (maximum), 50% (intermediate) and 100% (unselected controls). The various combinations of these factors were replicated to provide information on the repeatability of the selection response. All selection intensity lines at a given population size were replicated equally. Results are pre- sented for the first 14 generations of selection. The C16 and MI6 lines were examined previously for differences in growth rate, feed efficiency and body composition (TIMON and EISEN 1969, 1970; TIMON, EISEN and LEATHERWOOD 1970).
Selection and mating procedures: Within full-sib family selection was practiced throughout. In control lines a male and female were selected at random from each litter to provide parents of the next generation. I n order to achieve different selection intensities at the same population size it was necessary to vary the number of individuals considered for selection. Since all litters were standardized at five days of age to four males and four females, wherever possible, maxi- mum (M) selection intensity, in the context of this study, means selecting within each litter the individual of each sex with the largest postweaning gain. An intermediate (I) intensity of selection was achieved by randomly choosing two males and two females within each litter and selecting the member of each sex with the larger postweaning gain.
Pair-matings of selected mice were made at random except that full-sib matings were avoided where possible. The effective population size under the planned mating scheme is approximately 4N - 2 (ROBINSON and BRAY 1965). Selected parents were mated between eight and ten weeks of age on the same day each generation so that all lines were contemporaneous. Selections were made from first litters only, and all mice were weaned at three weeks of age, Mice were weighed to the nearest 0.1 g at 3, 6 and 8 weeks of age. In addition, number born and 12-day litter weight were recorded. Results of the correlated responses to selection for postweaning gain will be reported elsewhere.
TABLE 1
Design of the selection experiment
Population size'
1 2 4 8 16
Number of Selection intensity codes
replicates for each
selection intensity? Control (C) Intermediate (I) Maximum (M)
16
c1
I1 M I8 c2 I2 M2
4 0% I4 M 4
2 C8 I8 M8
2 C16 I16 M I 6
* Population size is expressed as the number of pairs of selected parents mated each generation.
EFFECTS OF POPULATION SIZE AND SELECTION INTENSITY 515
The lines were kept in one laboratory where temperature and relative humidity were main- tained at 22
*
2°C and 50 t 5%, respectively, with a 12 hr lighk12 hr dark photoperiod. Mice were fed Purina Laboratory Chow ad libitum except during lactation when a high energy ration (Emory Morse Co.) was fed ad libitum. Further details on laboratory procedures have been given by H A N R 4 H A N and EISEN (1972).Each line was represented by the designated number of matings together with reserves to be used as replacements, should any of the selected matings fail. Mice for reserve matings in the control (C) lines were chosen at random. I n the M lines the reserve mouse chosen from any littsr was the second highest ranked individual of the particular sex. For reserves in the I lines, the unselected member of the random pair was the only individual qualified to be mated as a reserve. Equal numbers of male and female reserves were chosen, and reserves in any line were pair-mated inter se only.
Two classification errors occurred during the course of the experiment and were not detected until inbreeding coefficients were computed. In generation 10 offspring of a n M16-1 mating were classified as being MIG-2 mice, and the same error occurred again at generation 12 involv- ing one litter. As a result of both migrations, 10 out of a total of 12 M16-2 litters at generation 14 had ancestry in M16-1. A consequence of this inadvertent migration between replicate lines
is that any comparison of these replicates to assess similarity of response pattern is compromised.
Ezpected response: Following the genetic model of HANRAHAN and EISEN (1972) which allows for maternal effects and a genetic correlation of less than unity between genotypic effects in males and females, the expected response per generation to within full-sib family selection is
*Gull = ?4 il (hi
+
hOl hml r0,pn1+
h O l ho2 r0,o:+
ho2 h m l U&AGw2 = 34 i, (h20
+
ho2hZL
r o * n t >+
4,
ho2 +0102+
hOl hm2 ro,m2) U P 2 K 21 for males and
for females, where
= heritability of the s effects in the i t h sex; s = direct (0) or maternal ( m ) additive
1
genetic effects, respectively, and j = males (1) or females (2),
r g = genetic correlation between the s effects in the i t h sex and the s' effects in the kth sex; e.g , rOlo2 is the genetic correlation between additive direct genetic effects in males and females,
I C
u p , = phenotypic standard deviation of the j t h sex, i, = standardized selection differential in the i t h sex,
ni = number of progeny of the j t h sex,
t1 = intraclass correlation among full sibs of the j t h sex.
I n the present experiment it is reasonable to assume that i, = I,, K , = K , , n, = n2, t , = t,. The above formulas clearly show that within full-sib family selection does not obviate contributions of correlated maternal genetic effects to the response.
Estimates of parameters for postweaning gain in the ICR base population (H~NRAHAN and EISEN 1972) are summarized in Table 2. Substituting these values into the precedkg formulas, and assuming ro = -.31 for all i and k, yielded genetic gain predictions of 0.22 and 0.33 g per generation for males and females, respectively, at maximum selection intensity. The cor- responding predictions for intermediate selection intensity were 0.12 and 0.18 g, respectively.
Measuring response and selection differential: Male and female responses were estimated by the generation means of all progeny with complete records regressed o n generations of selection. Since male and female responses were treated separately, realized selection differentials were computed for each sex. Thus, the realized selection differential for males was the mean of the male and female selection differentials weighted by the number of male progeny contributed to
516 J . P. H A N R A H A N , E . J . E I SE N A N D J . E. LEGATES
TABLE 2
Base population statistics for postweaning gain
Statistic Males Females
Mean*
Phenotypic variance ( u Z ) Coefficient of variation
Additive genetic variance (uZA )f P j
17.89 13.02 6.22 4.96 13.94 17.11 1.82 1.91
j
Heritability of direct effects (hz,, ) 0.29 0.38 Heritability of maternal effects (hz ) 0.38 0.08 Phenotypic correlation among full sibs (tj) 0.36 0.39 Genetic correlation between the sexes (ro
Genetic correlation between direct and maternal effects (ro .m = -0.31)
3
m j
= 0.71)
1 2
1 k
~~~
* 2568 males, 2562 females.
+
354 degrees of freedom for paternal half-sib families.the next generation. Realized selection differentials for females were computed similarly, except that the number of female offspring was used as the weighting factor.
Statistical analysis: The analysis followed the procedure given by RICHARDSON, KOJIMA and LUCAS ( 1 968). Data from generations zero through fourteen were summarized by computing male and female generation means for each line together with their appropriate cumulative selection differentials. The model used to describe a generation mean for either sex was
where:
I'iikl = mean of the lth replicate-kth population size-jth selection intensity combination in the ith generation (i = 0,1,.., 14; j = 1,2,3; k = 1,2 ,.., 5; I = 1,2 ,.., nk, the number of replicate lines at the kth population size),
O i = environmental effect common to all observations in the it* generation with the restriction that 0, = 0,
ailcl = intercept of i k l t h line (accounts for the sampling process used in deriving individuals for generation zero from the base population),
P j k Z = regression coefficient (realized heritability) of response on cumulative selection differen- tial ( Z i i k l ) for the iklth line,
E~~~~ = random error assumed to be NID ( 0 , u ' Z ) .
The model is of full rank due to the restriction that 0, = 0. To obtain the best linear unbiased estimates of the parameters it is necessary to know the variance-covariance structure of the residuals.
It was assumed that the variance of the residual effects ( E ~ was ~ ~u2/nijkl ~ ) where uz is the within-generation phenotypic variance and niik2 is the number of observations in the corre- sponding generation mean. It was also assumed that there was n o correlation between residuals in different generations of the same line. This assumption is violated to a degree by genetic drift. In addition, genetic drift causes the variance of a line mean to increase over time (HILL 1971 ) .
These effects of drift could not be determined from the data; thus, a weighted least squares analy- sis was performed using merely the square root of the corresponding n i j k l as the weighting factor for each generation mean. The resulting estimates of the parameters should be unbiased.
The sums of squares due to the regression coefficients were partitioned to allow tests of the effects of population size, selection intensity, their interaction and replicate differences, as shown in Table 3. To reduce the number of parameters in the model to facilitate a least squares s o h - tion, the full-sib mating lines (Cl, I1 and MI) were not entered as replicate means. Instead, generation means were computed for each selection intensity level, and these were used in the
EFFECTS OF POPULATION SIZE AND SELECTION INTENSITY
TABLE 3
Analysis of u a r i m e of the response to selection.
517
Source Degrees of freedom
Generations Intercepts Slopes (linear)
Control lines Population size (P)
Linear (PL) Quadratic (PQ) Remainder
Selection intensity (I us. M) P
x
(I us. M)PL
x
(I us. M) PQ x (I us. M) RemainderHomogeneity of replicates I2 M2 I4 M4 I8 M8 I16 MI 6
Error
14 . . 51 . . 51 . .
. . 17
. . 4 . . . . . . . . . . . .
. . I . . 4 . . . .
. . . .
. . . .
. . 7 . . 7
. . 3
. . 3
. . I . . 1 . . 1 . . 1 563 . .
. . . . . . . . . . 1 1 2 . . . . 1 1 2 . . . . . . . . . . . . . . . . . .
analysis. Cumulative selection differentials also were included for control lines since many con- trol lines, especially at the smaller population sizes, accumulated considerable selection differen- tials. In partitioning the sums of squares due to population size and selection intensity, contra1 line regression coefficients were not included in the contrasts.
RESULTS AND DISCUSSION
Surviual of lines: Table 4 gives a summary of the line survival for population sizes of one and two pairs in terms of the number of replicates present at genera- tions
5 ,
10 and 14. Despite the fact that reserve matings were used, replicate lines became extinct due to infertility at these population sizes. Thus, any comparisons involving these lines are seriously biased and results are presented for complete- ness only.TABLE 4
Number of replicate lines present at generations 5,10 and 14 Line
Generation c1 I1 M1 c2 I2 M 2
5 10 10 10 7 7 7
10 2 5 5 6 5 4
518 J. P. HANRAHAN, E. J. EISEN A N D J. E. LEGATES
TABLE 5
Inbreeding coefficients and uuriunce effectiue sizes, averaged over replicates, at generation 14
Inbreeding coefficients' - FA Fa FI FM N.4 N, NI N M Effective size+
1 2 Population 4 size (pairs) 8 16
0.951 0.676 0.379 0.951 0.756 0.421 0.951 0.731 0.541 0.951 0.843 0.495 6.00 14.00 4.05 10.52 4.32 7.07 3.46 7.72
0.197 0.196 0.265 0.262 30.00 22.08 20.60 18.64 0.100 0.1 15 0.124 0.131 62.00 49.53 48.28 40.85 * FA is the expected inbreeding coefficient under the experimental design.
F,, F,, FM are the observed inbreeding coefficients for control, intermediate and maximum
+
NA, N,, NI, N, are corresponding effective sizes. selection intensities, respectively.Inbreeding levels: Mean inbreeding coefficients and variance effective sizes (CROW 1954) were computed through generation 14. The results, averaged over replicate lines, are given in Table
5
together with the inbreeding levels expectedif no reproductive failures or other deviations from the mating plan had occurred. Selected lines had slightly higher inbreeding coefficients than control lines at population sizes 2,4, 8 and 16, with the exception of 12. This result is a reflection of the increased frequency of sterile matings as selection progressed.
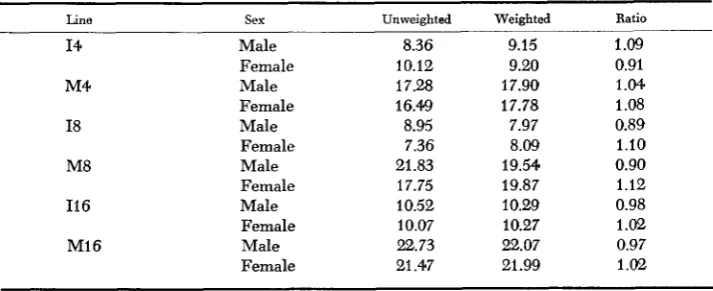
Selection diflerentials: Unweighted and weighted cumulative selection differ-
entials, averaged over replicates, are shown in Table 6 together with the ratio of weighted to unweighted selection differentials. When both sexes are averaged
TABLE 6
Unweighted and weighted cumulative selection differentials ( g ) and the ratio of weighted to unweighted cumulatiue selection differentials, averaged over replicates*
~~~
Line Sex Unweighted Weighted
I4 M4 I8
M 8
I16 MI 6
Male Female Male Female Male Female Male Female Male Female Male Female 8.36 10.12 17.28 16.49 8.95 7.36 21.83 17.75 10.52 10.07 22.73 21.47 9.15 9.20 17.90 17.78 7.97 8.09 19.54 19.87 10.29 10.27 22.07 21.99 Ratio 1.09 0.91 1.04 1.08 0.89 1.10 0.90 1.12 0.98 1.02 0.97 1.02
E F F E C T S O F P O P U L A T I O N SIZE A N D S E L E C T I O N I N T E N S I T Y 519
there is little difference between the two methods of calculation. This result is not surprising since the selection differentials were computed on a within-family basis, averaged over families, and only one male and one female were selected per family. Realized selection differentials were analyzed for generation trends. The results showed little evidence of any major changes.
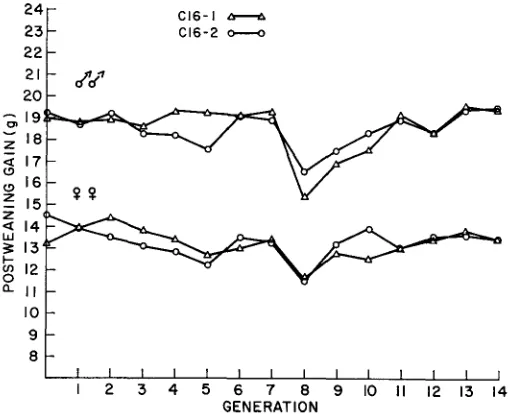
Selection response: Generation means for postweaning gain are plotted in Figure 1 for the C16-1 and C16-2 replicate lines. The decline at generation eight is a reflection of a reduction in 6-week body weight because of a failure of the temperature control system when the mice were about five weeks old, which re- sulted in a temperature of 13°C. There were extremely small cumulative selec- tion differentials of -.62 and 0.98 g in C16-1 and C16-2, respectively, indicat- ing that the random within-family selection effectively prevented any appreci- able selection for postweaning gain. The regressions of generation means on gen- eration number were non-significant in both replicates for postweaning gain (-.03
*
.07, -.04 f .04, 0.00* .05
and -.02*
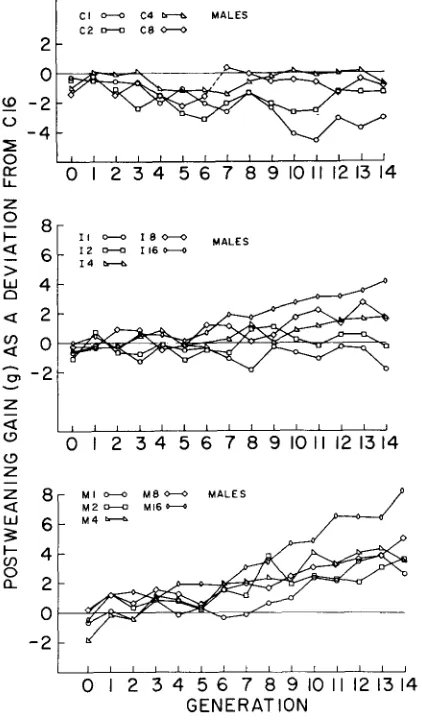
.04 f o r C16-1 males and fe- males and for C16-2 males and females, respectively). No significant differences between replicates were present for intercepts or slopes. It seems reasonable to assume that no appreciable genetic drift occurred in the C16 line for loci in- fluencing postweaning gain. It was therefore used as the reference line against which all other lines were compared to evaluate the effects of genetic drift and selection.Generation means for male postweaning gain, pooled over replicate lines and expressed as a deviation from C16, are shown in Figure 2. I t is evident that in- breeding in the smaller population size control lines has caused some depression in postweaning gain, especially in the C1 and C2 lines. The regressions of re-
C 1 6 - I
-
C16-2 o---o24
22
19
17
18
z
w 16
5 15
$
1413
cn 12
I 1 0
1 : 1 1 I I I , I , I I I I , I ,
8
I 2 3 4 5 6 7 8 9 1011 1 2 1 3 1 4 GEN E RAT ION
520 J. P. H A N R A H A N , E. J. EISEN A N D J. E. LEGATES
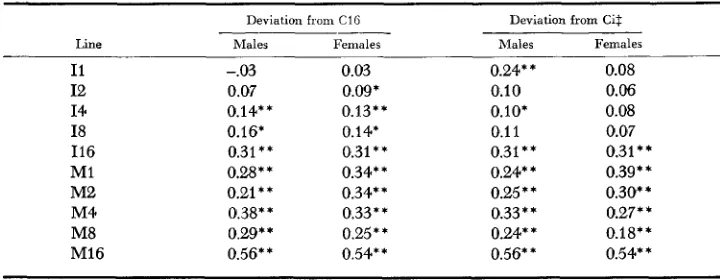
sponse on generations of selection are presented in Table 7. Regression coefficients are given as a deviation from C16 and as a deviation from the control of the same population size (Ci)
.
As expected, response per generation was significantly (P
<
. O l ) larger at the 25% (M) selection intensity than at 50% (I) within each population size. CLAY-TON, MORRIS and ROBERTSON (1957) reported that short-term selection response
for chaetae number in
D.
melanogaster increased as selection intensity increased. The genetic gains in the I16 and M16 lines were significantly (P<
. O l ) larger than those found in the smaller population sizes at the corresponding selection intensities. There was little difference in response between M8 and M4 or be- tween I8 and 14. The larger response in M16 and I16 relative to the smaller population size lines could be attributed only in small part to greater cumulativeL
C I e-c C4 M MALES c 2 w C 8
-
2
0
2 - 2
- 4
0
.c;
1 1 C - - a * 8 - MALES I 2 W 116-
I 4 w
0 I 2 3 4 5 6 7 891011121314
GENERATION
EFFECTS O F POPULATION SIZE A N D SELECTION INTENSITY
TABLE 7
Regressions of response on generation number ( g ) for each populaiion size-selection intensity combination, pooled over replicated
521
Deviation from C1F Deviation from Cis
Line Males Females Males Females
I1 I2 I4 I8 I16 MI M2 M4 M8 MI6 -.03 0.07 0.14** 0.16* 0.31** 0.28"' 0.21 * * 0.38** 0.29** 0.56** 0.03 0.09* 0.13** 0.14* 0.31** 0.34** 0.34* * 0.33** 0.25** 0.54** 0.24** 0.10 0.10' 0.1 1
0.31** 0.24* * 0.25 * * 0.33 * * 0.24' * 0.56* *
0.08 0.06 0.08 0.07 0.31** 0.39* * 0.30** 0.27' * 0.18** 0.54**
+
Results for 11, MI, I2 and M2 are biased because of extinction of replicates. a P<
.05,Ci refers to C1, C2, C4, C8 and C16, respectively.
* * P
<
.01.selection differentials. Thus, these results suggest the effects of genetic drift in reducing selection response in populations of relatively small effective size, as predicted by ROBERTSON (1960). The data are in general agreement with the studies of LEWIS and
WARWICK
(1953),
T A N T A W Y (1956),
CHUNG and CHAP- MAN (1958) and FRANKHAM, JONES and BARKER (1968). Because selection waspracticed within full-sib families and single-pair matings were utilized, the in- fluence of genetic drift would be expected to be minimal. Nevertheless a sizeable effect of genetic drift was observed under this selection and mating procedure, even though the selection experiment was of a relatively short duration.
Since many one and two pair replicate lines became extinct, there were only two valid comparisons of genetic gain per generation between lines having the same expected number of individuals considered for selection
(57):
M8 us. I16( T = 64) and M4 us. I8 (2' = 32). The M8 and I16 lines showed no significant difference between selection response when averaged over replicates. Based on the cumulative selection differentials, the observed response in I16 was almost twice that in M8. Such an outcome has been predicted for the selection limit based on theoretical considerations (DEMPSTER 1955; ROBERTSON 1960). FRANK-
HAM, JONES and BARKER (1968) reported a similar result in selection for bristle number in
D.
melanogaster. In contrast, the selection response was significantly(P
<
.01) greater in M4 compared with I8 and could be explained on the basisof the difference in selection differentials. An explanation f o r this discrepancy at these particular population sizes must await an approach of the lines to the selec- tion limit.
522
I O
9 -
-
"0 2 4 6 8 IO 12 14 16 18 20 22
CUMULATIVE SELECTION DIFFERENTIAL
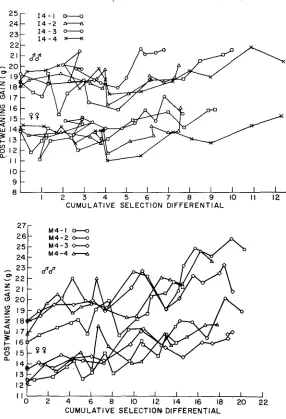
FIGURE 3.-Generation means for male and female postweaning gain in the individual repli- cates of the I4 and M4 populations, plotted against the cumulative selection differential.
differential in some generations which arose as a consequence of using reserve matings. At intermediate selection intensity a reserve had a probability of 0.25 of being the smallest individual in the litter. Thus, use of a reserve could cause a net negative selection differential in I4 lines since the selection differential is averaged over four litters, at most.
EFFECTS O F POPULATION SIZE AND SELECTION INTENSITY
116-1 bd
116-2 o-*
0 I 2 3 4 5 6
7
8 9 IO I I18-1
1 8 - 2 - 4
523
I I I I I I I I I I I J
0 I 2 3 4 5 6
7
8 9IO
IIC
U M U L AT I V ESE
L
ECT I ON DIF F
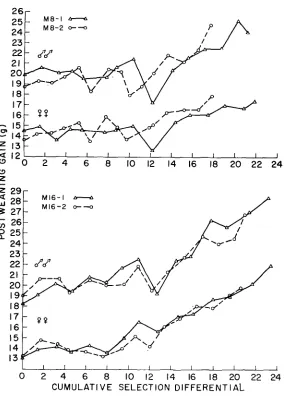
E R E N T I A L4.-Generation means for postweannig gain in the individual replicates of the I8 and I16 populations, plotted against the cumulative selection differential.
responding male component was negative but failed to reach significance. Exami- nation of Figure 3 shows that this line made no net advance after generation seven. Positive quadratic responses were observed in M16, MS, I16 and I8 lines. The quadratic component was significant for both sexes in M16-1 and M16-2 and for males in M8-1, M8-2 and 116-1. Possible causes of the non-linear response are discussed in the next subsection.
524 J . P. H A N R A H A N , E. J . E I S E N A N D J. E. LEGATES
(3 0 2 4 6 8 IO 12 14 16 18 20 22 24 (3
I I I I I I I I I I I I I 0 2 4 6 8 IO 12 14 16 18 20 22 24
CUM U LAT I V E SELECT ION D I FFE R E NT I AL
FIGURE 5.-Generation means for postweaning gain in the individual replicates of the M8 and MI6 populations, plotted against the cumulative selection differential.
EFFECTS O F POPULATION SIZE A N D SELECTION INTENSITY
TABLE 8
Weighted average realized heritabilities of postweaning gain estimated from complete model*
525
Selection intensity
Intermediate Maximum Difference
ropulation -
-size (pairs) Sex (1) (M) W-1)
1 2 4 8 16 Mean M F M F M F M F M F M F
0.052 t ,071
0.046 t .062 0.221 t .051 0.204 k .047 0.241 t .043 0.150 k ,037 0.320 f .049
0.196 k .042
0.486 f ,030 0.399 t .026 0.339 f .019 0.262 t .017 ..__...
0.283 f .039 0.287 t .035 0.225 k .MI 0.B1 f .034 0.320 f .022
0.241 f .019 0.235 rfr .021 0.163 f .018 0.376 k .014 0.329 f .012 0.322 f .OW 0.268 f .006 . . .
0.230 k .080
0.243 f .071 0.m f .OG
0.017 f ,056 0.079 rfr .W 0.091 t .041 -0.085 f .053 -0.033 f .045
-0.110 f .034 -0.070 f .028
* The realized heritabilities are denoted by pjk, in model given under Statistical analysis.
a lower realized heritability under inbreeding as compared with outbreeding when selection was for body weight in mice. TANTAWY (1956) reported similar findings for wing length in
D.
melanogaster.The analysis of variance (Table 9) showed a significant selection intensity X
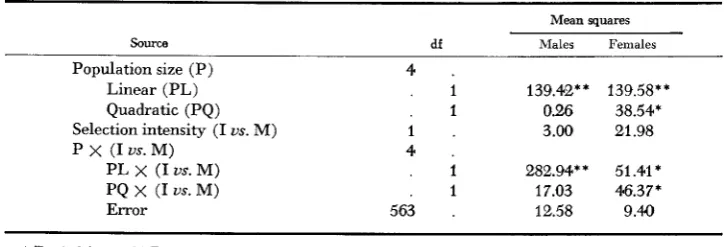
population size interaction but no overall significant difference between selection infensities. At population sizes of less than eight pairs, greater realized heritabil- ity was found at maximum intensity than at intermediate intensity. I n contrast, at population sizes of eight and sixteen pairs the larger realized heritabilities were achieved at the intermediate selection intensity.
At smaller population sizes the advantage of maximum selection intensity is
TABLE 9
The analysis of variance showing the effects of population size and selection intensity on realized heritability
Mean squares
Source df Males Females
Population size (P) Linear (PL) Quadratic (PQ) Selection intensity (I us. M) P x (I us. M)
PL x (I us. M) PQ x (I us. M) Error
4 .
. I 139.42** 139.58**
. 1 0.26 38.54*
1 . 3.00 21.98
4 .
. 1 282.94** 51.41 *
. I 17.03 46.37*
563 . 12.58 9.40
526 J. P. HANRAHAN, E. J. EISEN A N D J. E. LEGATES
to reduce the rate of genetic drift. The apparent advantage of intermediate selec- tion intensity at the largest population size is not readily explained. However, the significant result may be a consequence of the downward bias in the esti- mated standard errors (HILL 1971). Also, the regression coefficients were com puted ignoring the positive quadratic response in M16-1 and M16-2 which would cause the linear regression coefficient to underestimate response at the later gen. erations. Therefore, realized heritability in the I16 and M16 lines was calculated by taking the line means at generation
14,
as a deviation from C16, divided by the cumulative weighted selection differentials (Table IO). The values show that while I1 6-1 gave substantially higher realized heritabilities than any other line, the next highest line is M16-1 with 116-2 and M16-2 being about equal. There is, therefore, no justification for concluding that there is any difference in realized heritability between selection intensities at this population size.Causes of non-linear response: There are a number of possible genetic explana- tions for the positive quadratic response. The simplest explanation requires low initial frequency for the desired alleles. Other explanations depend on the pres- ence of dominance type genetic effects (GILL 1965; HILL 1969). However, in
view of the simulation results of GILL (1965) and the absence of evidence for much non-additive genetic variance in body weight of mice (ROBERTS 1965),
this is an unlikely explanation.
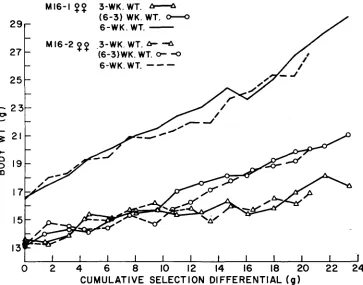
The response patterns for 3- and 6-week body weight suggest that maternal effects may have caused the initial lag in response. I t may be seen in Figure 2 that there was a reasonable response for postweaning gain in the first generation followed, in general, by a couple of generations during which little response OC-
curred. Figure 6 shows the response in females for 3- and 6-week body weight and postweaning gain, corrected for environmental effects. The most striking ob- servation is the linear response in 6-week weight, particularly during the early generations of selection. The pattern for 3-week weight shows no response in the first generation, followed by considerable response in the next three generations. The response in weaning weight is probably due to a maternal effect. It is pos- sible that this increase in weaning weight may have retarded the rate of post- weaning growth by a negative compensatory effect. That the observed increase in weaning weight was due to postnatal maternal effects rather than to direct
TABLE 10
Realized heritability measured as m a n response at generation 14, as a deviation from C16, diuided b y the cumulative selection differential
Line Male Female
116-1 11&2 Ml6-1 M I 6 2
0.467
*
.078* 0.454*
.072 0.331 t .a73 0.340 t .067 0.389 & .033 0.362 k .030 0.353 .036 0.313*
.034EFFECTS O F POPULATION SIZE AND SELECTION INTENSITY 527
M16-I
$9
3 - W K . WT. 6--6(6-3) WK. W T . M
6 - W K . WT.
-
M16-2 $ $ , 3 - W K . W T . 6- 4 27 (6-3)WK. WT. 0- -0
- . . . -
2 5 F
6 - W K . WT.
---
/
0
/ J '
A .
I
-
2 3 1D
u t
-"k;-
170 2 4 6 8 IO 12 14 16 18 20 22 24
CUMULATIVE SELECTION DIFFERENTIAL ( 9 )
FIGURE 6.-Generation means for female three- and six-week weights and postweaning gain
in M16-1 and Ml6-2, adjusted for environmental effects, plotted as a function of the cumulative selection differential.
genetic effects is supported by the results of TIMON and
EISEN
(1970). They found no differences in weaning weight when litters of M16 and C16 mice were nursed by C16 dams.Replicate variability: Analysis of differences among replicate lines in selection response is presented in Table 1 1 . Replicate differences were significant in one or both sexes for 12, M2, 14, M4 and M16 lines. Figure 3 clearly shows wide fluctuations in the I4 and M4 replicates. Examination of the response pattern in
M16 (Figure 5) suggests that there were no replicate differences of any conse- quence. It seems reasonable to conclude that for population sizes of eight and sixteen pairs there is good agreement between replicate lines. Differences between replicates decreased as the effective size increased, and at population sizes of less than eight pairs there was greater homogeneity among maximum intensity lines than among those being selected at intermediate intensity. This result is in agree- ment with that of FRANKHAM, JONES and BARKER (1968).
Observed vs. predicted response: The predicted genetic gain was much less than
528 J . P. H A N R A H A N , E. J . E I S E N A N D J. E . LEGATES
TABLE 11
Analysis of variance for testing homogeneity of realized heritability among replicate selected lines
Mean squares
Source df Males Females
I2 M2 I4 M4 I8 M8 I16 MI6
Error
7 7 3 3 1 1 1 1 563
31.46* 9.71 41.65* 32.76* 2.25 17.84 12.38 1 74.75 * *
12.58
12.65 22.78*
56.23**
24.20+ 0.31 20.21 8.15 36.58*
9.4.0
* P < . o 5 , * * P < . O l .
t
Approaching P<
.05.in Table 2 yielded predicted realized heritabilities of 0.13 and 0.21 for males and females, respectively. The observed values in Table 8 are much larger for the I16 and M16 populations. If the assumptions are imposed that rolOL = 1 .O and roIm = 0, then the predicted realized heritability becomes 0.29 and 0.26 for males and fe- males, respectively. These values are still less than those observed in the 16 pair lines. If one considers the 95
% confidence interval for the estimate of heritability
of direct effects, the observed realized heritabilities could not be declared signifi- cantly different from the predictions.Effect of selection on coejjicients of variation: Coefficients of variation (CV)
were estimated each generation using pooled variances over replicate lines. There was no evidence for any change in CV as a consequence of repeated truncation selection through 14 generations. FALCONER (1953) reported a similar result for &week body weight over 12 generations of selection.
G E N E R A L DISCUSSION
EFFECTS O F POPULATION SIZE A N D SELECTION INTENSITY 529
In
selection experiments with farm animals the number of male parents is usually so much smaller than female parents that the effective population size is determined by the former. I n addition, the expected effective size will depend on the sampling procedure adopted for choosing replacements.If
both sires and dams are chosen at random, then the effective size can be approximately4
N m ,where
N,
is the number of male parents. If sampling is designed so that each male parent contributes equal numbers of progeny and each female is replaced by a daughter and either one or no son, then the effective size is approximately16
-
N,
(GOWE, ROBERTSON and LATTER 1959). Thus, between the two extremes3
possible with farm animals where female parents greatly outnumber males, between 12 and 16 sires would be necessary to achieve target effective population sizes in the region of 62.
The results on the effects of selection intensity indicate that at effective sizes likely to be used in practice there is no advantage to be gained by selecting at a n intensity less than the maximum possible. Future analysis of the present experi- ment hopefully will yield information on the long-term effects of selection in- tensity.
Thanks for many critical discussions of the problem are extended to DRS. A. F. PURSER,
C. C. COCKERHAM, J. RAWLINGS, T. MUKAI and 0. W. ROBISON. The valuable technical assistance of MRS. BILLIE JEAN EDWARDS is appreciated.
LITERATURE CITED
CHUNG, C. S . and A. B. CHAPMAN, 1958 Comparisons of the predicted with actual gains from selection of parents of inbred progeny of rats. Genetics 43: 594-600.
CLAYTON, G. A., J. A. MORRIS and A. RomR'rsoN, 1957 An experimental check on quantitative genetic theory. I. Short-term response to selection. J. Genetics 5 5 : 131-151.
CROW, 5. F., 1954 Breeding structure of populations. 11. Effective population number. Sta- tistics and Mathematic? in Biology. Edited by 0. KEMPTHORNE, Iowa State College Press, Ames.
DEMPSTER, E. R., 1955 Genetic models in relation to animal breeding problems. Biometrics 11: 535-536.
FALCONER, D. S., 1953 Selection for large and small size in mice. J. Genetics 5 1 : 470-501.
-
, 1960 Selection of mice for growth on high and low planes of nutrition. Genet. Res. 1: 91-113.The effects of population size and selec- tion intensity in selection for a quantitative character in Drosophila. I. Short-term response to selection. Genet. Res. 1 2 : 237-248.
Effects of finite size on selection advance in simulated genetic populations.
Environment and poultry breeding prob-
Sexual dimorphism and direct and maternal genetic
The rate of selection advance for non-additive loci. Genet. Res. 13: 165- FRANKHAM, R., L. P. JONES and J. S. F. BARKER, 1968
GILL, J. L., 1965
GOWE, R. S., A. ROBERTSON and B. D. H. LATTER, 1959 HANRAHAN, J. P. and E. J. EISEN, 1972
HILL, W. G., 1969
Aust. J. Biol. Sci. 18: 599-61 7.
533 J. P. H A N R A H A N , E. J. E I S E N A N D J. E. LEGATES
173. -, 1971 Design and efficiency of selection experiments for estimating genetic parameters. Biometrics 27: 293-31 1.
Experimental studies of the distribution of gene frequencies in very small populations of Drosophila melanogaster. I. Forked. Evolution 8 : 172-177. -, Experimental studies of the distribution of gene frequencies in very small populations of Drosophila melanogaster. 111. Aristapedia and spineless. Evolution 8 : 293- 301.
Effectiveness of selection for body weight in mice from inbred and outbred populations derived from common parent stocks. J. Hered. 44: 233-238.
An analysis of short-term selection ex- periments. Heredity 23 : 493-506.
Some contributions of the laboratory mouse to animal breeding research. Part I. Animal Breeding Abst. 33: 339-353.
A theory of limits in artificial selection. Proc. Roy. Soc. London, B 153:
234-249.
Expected effects on the inbreeding coefficient and rate of gene loss of four methods of reproducing finite diploid populations. Biometrics 21 : 447-458. Selection for long and short wing length in Drosophila melanogaster
with different systems of mating. Genetica 28: 231-262.
Comparison of growth curves of mice selected and un- selected for postweaning gain. Theor. Appl. Genetics 39: 345-351.
-
, 1970 Compari- sons of ad libitum and restricted feeding of mice selected and unselected for postweaning gain. I. Growth, feed consumption and feed efficiency. Genetics 64: 41-57.Comparisons of ad libitum and re- stricted feeding of mice selected and unselected for postweaning gain. 11. Carcass compo- sition and energetic efficiency. Genetics 65 : 145-155.
WRIGHT, S., 1938 Size of population and breeding structure in relation to evolution. Science 87: 430-431.
]<ERR, W. E. and S. WRTGHT, 1954a 1954b
LEWIS, W. L. and E. J. WARWICK, 1953
RICHARDSON, R. H., K. KOJIMA and H. L. LIJCAS, 1968
ROBERTS, C. R., 1965
ROBERTSON, A., 1960
ROBINSON, P. and D. F. BRAY, 1965 TANTAWY, A. O., 1956
TIMON, V. M. and E. J. EISEN, 1969