i
I1Ulian J. Plant PhYliol., Vol. XXIX, No.4, pp. 335-344 (December.1986)
,.
with each other. DCMU and atrazine can replace DNOC from its binding site of the chloroplast membrane.
4. In the susceptible biotype tbe main TL band which is related to the secondary quinone electron acceptor Q. (Q.-band) apeared at +30·C. This band is totally absent in the resistant biotype.
5. In the resistant biotype a TL band at +IO·C can be observed Which is assigned to the primary acceptor Q. (QA-band).
INTRODUCTION
Differential response to triazine herbicides has been observed within certain
weedspecies for example, Ryan (1970) and Radosevich and Devilliers (1976)
Department of Botany, University of Chittagong. Chittagong, Bln gladesh.
TRIAZINE RESISTANCE IN ERIGERON CANADENSIS L. IV
INVESTIGATED BY THERMOLUMINESCENCE
MEASUREMENT
A. RASHID· AND S. DEMETBR
Biological Research Centre, Institute of Plant Physiology, Hungarian Academy of Sciences, H-6701 Szeged. Hungary.
SUMMARY
Thermoluminescence (TL) after continuous illumination was studied in untreated and herbicide treated (atrazine.2-chloro-4-etbyla mino-6-isopropylamino-s-triazine; DCMU, N-(3-4-dichlorophenyl)-N-N dimethyl-urea; DNOC. dinitro-o-cresol) chlosoplasts of triazine-suscepti ble and -resistant biotypes of Eri,eron canmieNlil L. The results obtained from the above experiment can be summarized in the following mannel'$ :
1. TL can be applied in the approximate determination of 16a value of photosytltem II (PS II) herbicides in the treated plants.
2. In the susceptible chloroplasts herbicide binding infiuences the redox state of the primary quinone electrone acceptor QA, which is reflected on the peak position of the main TL band.
336
A. RASHID AND S. DEMETERreported in common grounsel
(Senecio vulgaris),
Radosevich (1977) reported in
redroot pigweed
(Amaranthus retroflexus)
and Machado
et al.
(1978) reported in
common lambsquarter
(Chenopodium album).
The difference is not associated
with uptake or metabolism of the inhibitor but of difforential chloroplast function
as related to the photosynthetic electro transport from
QA
to
QB
in the reducing
side of PS
[I(Pfister and Arntzen, 1979).
It
has been stated that TL provides
information about the redox state of the primary and the secondary acceptors of
PS II (Inoue and Shibata, 1978, 1979 and Droppa
et al.
1981). Therefore, the
present investigation was undertaken to characterizc the modifications occurring
in the redox state of the primary acceptor
QA
and the secondary acceptor
QB
due
to mutation induced alteration in the
QA-QB
herbicide binding complex of the
I
chloroplast membrane of
E. canadensis
L.by using TL measurement technique.
The variation in electron transfer rates from
QA
to
QB
was also one of the
objectives of this investigation.
MATERIALS AND METHODS
The supply of seedlings, the growing of seedlings and the isolation of
chloroplasts were described
elsewhere (Rashid and Demeter,
1985a).
Measurement of thermoluminescence :
The preparation of samples and the
technique of TL measurements have also been described previously (Rashid and
Demeter, 1985e). Samples were illuminatea with white light from a NARV A
halogen lamp of 650 W for 2 min at-60°C. The exciting light was passed through
a heat absorbing water filter (10 cm thickness) and a Balzer neutral density filter
giving an illumination intensity of 10 Wm-
2•The samples were heated in darkness
at a rate of :lO°C/min in order to get best resolution of the peaks (Sane
et al.
1977). Urea-type (atrazine and DCMU) and phenolic (DNOC) herbicides of
different concentrations were mixed with the samples before putting into the
sample holder.
RESULTS
TL originates from PS II and the bands of the glow curve can be related
to the charge recombination between positively charged donors and negatively
charged acceptors (Inoue and Shibata, 1978; Demeter
etal.
1979). In our
experiment the main TL band has been found to appear at + 30°C during conti
nuous heating from -60°C to + 80
a337
et al.
1975)_ In the present experiment the relative intensity of the main TL
band of the glow curve was measured by using different concentrations of PS
II herbicides (atrazine, DCMU and DNOC) in isolated triazine-susceptible and
-resistant chloroplasts of
E.canadensis
L.The QB-band at 30"C has been found
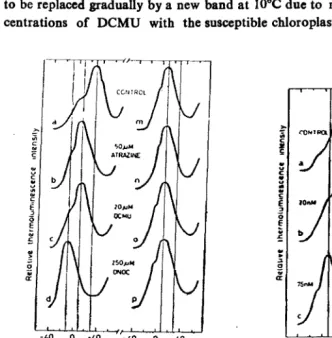
to be replaced gradually by a new band at IOoC due to mixing of different con
centrations of DCMU with the susceptible chloroplasts (Fig
1).
Atrazine and
..
..
,;1
"
..
or.40 O.lD -40 0 ·40 -40 0 '40 -40 0 .40
Temperature (-C) Temp,uotur. ro
Fig. 1: Gradual disappearance of Pig. 2: Comparison of the effect the Qs-band at+30·C and of atrazine, DCM and concomitant appearance DNOC on the peak posi of \theQ.cband aHI0·C tions of the glow bands in the glow curye of TL of TL in susceptible (a, due to the treatment of b, c and d) and resistant susceptible chloroplasts (m, n. 0 and p) chloro
with DCMU. plasts.
338
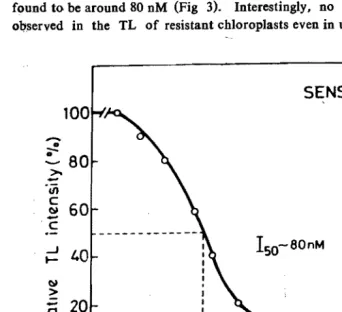
A. RASHID AND DEMETERfGund to be arGund 80 nM (Fig 3). Interestingly, no. QB-band at
+
30
0e was
G~erved
in the TL Gf resistant chlGrGplasts even in untreated cGntrGI sample.
SENSITIVE
-
...
•
-
:>. "Cijc:
toI
-
c:
~t
toI
>
-g
toI
a::
80
60
40
20
10-8
I
so
-80nM
10-
7DCMU(M~
Fig. 3: Estimtion of Ito concentration of DCMU in the susceptible chloro plasts by plotting the amplitude of the Qb-band as a function of
increasing concentrations of DCMU.
A band at
+
lOoe (QA-band) was fGund Gnly in this case. That band remained
unchanged (Fig 2) at high concentrations of atrazine, DeMU and DNOe 50 p.M,
20 p.M and 250 I'M respectively).
~Since DeMU and DNOC induced TL bands are well separated frGm each
Gther therefGre, we tried to follGW the displacement of these herbicides in the
chlGrGplast membranes. Fig 4 shGWS the concentration dependence Gf the dis
Temperatur. rcl
Fig. 4 : Displacement of bound DNOC from the susceptible chloroplast tbylakoids by DCMU as monitored by TL. After S min incubation of thechJoroplasts with 200 uM DNOC, different cancentrations of DCMU were added after 2min. a} no DCMU added b} 7S nM DCMU c) 200nM
DISCUSSION
Measuremeut of oxygen evolution (Rashid and Demeter, 1985a), flash
induced SIS nmabsorption change (Rashid and Demeter, 1985c) and fluorescence
induction (Rashid, 1983) could be applied in the determination of Iso concentra
tion ofPS IT herbicides (atrazine. DCMU and DNOC) of
Erigeron canadensis
L.
However, these techniques could not give any data about the changes occuring
at the acceptor side of PS II accompanying with the appearance of herbicide
resistance in this species. Therefore, we tried to introduce a new method, TL
for the investigation of the mechanism of herbicide resistance in plants. It has
been stated that TL provides information about the redox states of tbe primary
and the secondary acceptors of PS II (Inoue and Shibata, 1979; Droppa
et al.
1981). Thus TL seems to
be
an useful technique for studying the mechanism
and sites of action of herbicides that inhibit the electron transport of PS II at
level the of secondary acceptor QB.
In the current experiment, the
~-bandat + 30°C has been found to
be
340
A. RASHID AND S. DEMETER/
herbicides blocked the electron transport QA to QB (Trebst and Draber, 1979)
and as a results electrons accumulated on QA (Velthuys and Amesz, 1974).
Charge recombination of the reduced primary acceptor QA- with the donor side
of PS II generated the QA-band. Plotting the amplitude of the
QB~bandas a
function of DCMU concentrations the
150value was found to be 80 nm (Fig 3).
This finding is in good correlation with our data obtained from oxygen evolution
(Rashid and Demeter 1985a) and
SIS
nm absorption change measurements
(Rashid and Demeter, 1985c).
The differences in the peak positions of the
QA~bandsmay
beexplained
in the following ways:
(I)The structural modifications of the proteinaceous
components of QA and QB due to binding of DCMU, atrazine and DNOC
changes the mutual orientation and separation of QA and P680, so that probabi
lity of the reverse electron transport from QA to P680 might also change.
(2) TL originates from charge recombination occurring between positively
charged donors and negatively charged acceptors (Arnold, 1966: Lurie and
Bertsch, 1974; Ichikawa et al. 1975; Inoue and Shibata, 1979; Demeter et
01.
1979). The peak position ofTL band is determined by the redox span between
the donors and the acceptors molecules participating in the recombination (Vass
et al. 1981). The herbicide induced change in the redox state of QA is reflected
in shifts in the peak positions of the
QA~bands.The two different peak positions
(around
+
lOoC for DCMU and atrazine and -lOoC for DNOC) obtained in
our TL measurement can be supported by a classification done by Trebst and
Draber (1979), who classified urea-type and phenOlic herbicides into two different
groups on the basis of their chemical structure. We therefore, assume that
herbicides belonging to two different groups have different effects on the state of
QA which
is
evident from our results. Boger
eJ01. (1981) proposed binding sites
for urea-type and phenolic herbicides. Our results can also be supported by the
work of Pfister et al. (1981), who reported a 32 kDaIton polypeptide for binding
urea-type herbicides and Oettmeier and Masson (1980), who reported a 41
kDalton polypeptide for binding phenolic herbicides on the reducing side of
PS
n.
Since DCMU and DNOC induced TL bands are well separated from each
other, therefore, we tried to follow the displacement of these herbicides in the
susceptible chloroplast membrane. The results obtained from· the displacement
experiment suggest that DCMU, atrazine and DNOC have a common site of
action in chloroplast membrane. Our displacement experiment can be supported
by the work of Reimer et
01. (1979) and Trebst (1979). who demonstrated that
bromonitrothymol (BNT), a phenolic herbicide has replaced the bound radio
actively labelled metribuzin (C
IJL
3,
llt.l~..
I
J
TRIAZINE RESISTANCE 'IN ERIGERON CANADENSIS
341
concentration range of their
150value. Oettmeier
et al.
(1982) showed that the
ureas interfere noncompetitively but the phenolic herbicides interfere competi
tively with the specific binding sites of the phenolic herbicide. dinoseb.
The apparant contradictory results can be resolved
byexplaining that
there might have different but overlapping binding areas for DCMU, atrazine
and DNOC. The binding of one herbicide may influence the binding of the
other. Our interpretation can be supported
bya model proposed by Trebst
(1981).
In the above mentioned TL experiments it has been noticed that the' 'QB
'band appeared at
+
30°C in the case of susceptible biotype, Due to the treat
ment of DCMU, atrazine and DNOC, the electron transport is blocked between
QA to QB and consequently the QB-band has been found to be replaced by the
QA-band at
+
10DC,
+
8°C and -IODC respectively in susceptible chloroplasts.
Surprisingly, the QB-band did not appear at all at +30°C even in untreated
control sample of resistant chloroplasts. A band at + IODC (QA-band) has been
fonnd to appear in the glow curve of TL in this case. These observation suggest
that following light excitation of the resistant chloroplasts, the primary acceptor
pool remains in reduced state for a long period while in the susceptible biotype
Q-A is quickly reoxidized by the secondary acceptor QB.
A mechanism which can explain our result would
be
that in the resistant
chloroplasts, the semiquinone QB- is stabilized (i.e. mid-point potential of
QB/QB- is raised) so that the center would be predominantly in QB- state after
dark adaptation. The more positive potential of Qs- explains why the singly
reduced secondary acceptor QB- does not take part in charge recombination
reaction and consequently the lack of the QB-band in the TL spectrnm.
It
has been shown by Bowes
et al.
(1980) that in succeptible chloroplasts
the rate of the reaction QA-Qs-+QAQS- is faster than the reaction QA-QB--+
QAQB
1-.According to the redox theory of qui nones (Clarke, 1960), a more
positive redox potential of QB- results in an equivalent lowering of the redox
potential of QB
lt- .
Thus in resistant chloroplasts the QA-Qs- -+ QAQB2-
reaction
342 A. RASHID AND S. DEMETER
biotype chloroplasts showed a much higher "intermediate"
(FI)level in the
resistant chloroplasts than in the susceptible types. This implies that the rate
of reoxidation of the primary electron acceptor by the secondary electron accep
tor pool is slower in the resistant biotype than in the susceptible one.
In the susceptible chloroplasts the QA-band is shifted upon herbicide
addition indicating that not only the state of QB but that of QA also is influenced
by herbicide binding to the chloroplast membrane. Opposite to this, in resistant
chloroplasts the QA-band remained unchanged even at high concentrations of
DCMU, atrazine and DNOC. This observation suggests that in the resistant
chloroplasts the
QA~QBcomplex has changed due to the mutation of chloroplast
DNA (Pfister and Arntzen,
1979)and herbicide binding at the level of the
secondary acceptor does not exert any effect on the redox state of the primary
acceptor.
ACKNOWLEDGEMENT
The first author gratefully acknowledges the Hungarian Academy of
Sciences and UNESCO for providing him financial support for tbis research
work.
REFERENCES
Arnold, W. (1966). Light reactions in green plant photosynthesis: A method of study. Science, 154: 1046~J049.
Boger, P., Sandmann, G. and Miller, R. (1981). Herbicide resistance in a mutant of the microalga Bumiller;oJllil fili/orm;l. Photol"n. ReI.• %: 61-74.
BoWes. J., Crofts. A. R. and Arntzen, C. J. (1980). Redox reactiODs on the reducing side of photosystem II in chloroplasts with altered herbicide binding properties. Archil'. Biochem. BiophYI., %00: 303-308.
Clarke. W. M. (1960). Oxidation reduction potential of organic system. Waverly Press. Baltimore, Md.
Demeter, S., Droppa. M., Vass, I. and Horvath. G. (1982). The theromoluminescence of chloroplasts in the presence of photosystem II herbicides. Photochem. Photobio phYI .• 4: 163-168.
Demeter, S., Herczeg, T., Droppa, M. and Horvath. G. (1979). Thermolum inescence charac teristics of granal and agranal chloroplasts of maize. FEBS fAlters, 100; 321-324.
Droppa, M.• Horvath. G. and Demeter. S. (1981). Effect of photosystem II herbicides in triazine-susceptible and resistant Amaronlhus tetroftexUI biotypes investigated by thermoluminescence. Z. Plonzenphysio/. Bd.• 105: 75-79.
b
.1.
U;;hTRIAZINE RESISTANCE IN ERIOERON CANADENSIS
343
Ichikawa, L, Inoue. Y. and Shibata, K. (1975). Characteristics of thermoluminescence .bands in relation to the water-splitting activity in photosynthesis. Biochim. Biophys. Acta., 408: 228-239.
Inoue, Y. and Shibata, K. (1978). Oscillation of thermoluminescence at medium-low tempe rature. FEBS Letters, 8S: 193-197.
Inoue, Y. and Shibata, K. (1979). Thermoluminescence from oxygen evolving system of photosynthesis. Trends Biochem. Sci., 4: 182-184.
Lurie, S. and Bertsch. W. (1974). Thermoluminescence studies in photosynthetic energy con version. I. Evidence for three types of energy storage by photoreaction II of higher plants. Biochim. Blophys. Acta.,357: 420-428.
Machado, S. V., Arntzen, C. J., Bandeen, J. D. and Stephensen, G. R. (1978). Comparative triazine effects upon photosystem II photochemistry in chloroplasts of two com mon lambs quarters (Chenopodium album) biotypes. Weed Science., 26: 318-322.
Oettmeier. W. and Masson, K. (1980). SynthCliis and tbylakoid membrane binding of the radioactively labelled herbicide dinoseb. Pest. Biochem. Pllysiol .• 1486-97.:
Oettmeier, W., Masson, K. and Johanningmeir, U. (1982). Evidence for two different herbi cide binding proteins at the reducing side of photosystem II. Biochlm. Biophys. Acta,. 679; 376-383.
Pfister. K. and Arntzen, C. J. (1979). The mode of action of photosystem II specifiic inhibi tors in herbicide resistant weed diotypes. Z. Natur/orsch., 34c: 996-1009.
Pfister, K., Steinback, K. E., Gardner, and Arntzen. C. J. (1981). Photoaffinity labelling of an herbicide receptor protein in chloroplast membranes. Proc. Natl. Acad. Sci., U. S. A•• 78: 981-98S.
Radosevich, S. R. (1977). Mechanism of atrazine resistance in lambsquarters and pigWeed. Weed Science., 25: 3Hi-318.
Radosevich, S. R. and Devilliers, O. T. (1976). Studies on the mechanisms of striazine resis tance in common groundsel. Weed ScieflCe., 14: 229·232.
Rashid. A. (1983). Comparative spectroscopic investigation of herbicidal effect upon photo system II photochemistry of chloroplasts isolated from triazine· susceptible and resistant biotypes of Eriger(Jn canadensis L. Ph. D thesis, Jozsef Attila Univ. Szeged, Hungary.
Rashid, A. and Demeter. S. (1985a). Triazine resistance in Erigeron canadensis L. I. Investi galed by photosynthetic oxygen evolution measurement. Bangladesh J. Bot., 14: 133·139.
Rashid. A. and Demeter. S. (1985c). Triazine resistance in E. canadensis L. III. Investigated by flash-induced 515 nm absorption change measurement. Nuclear Science & Applications. (Bangladesh Atomic Energy Commissions Journal), Series A : Bio. Sci., 16: In press.
Rashid, A. and Demeter, S. (l'85e). Differences in the oscillatory pattern of thermolumines cence in chloroplasts isolated form triazineresistant and susceptible biotypes of E. canadensis L. Chitlagong University Studies, Part II: SciefICe: In press. Reimer, S., Link, K. and Trebst. A. (1979). Comparison of the inhibition of photosynthetic
344
A. RASHID AND S. DEMETERRyan, G. E. (1970). Resistance of common groundsel to simazine and atrazine. Weed Science .• IS: 614-616.
Sane. P. V .• Desai. T. S., Tatake. U. G. and Govindjee (1977). On the origin of glow peaks ia Eu,lena cells, spinach chloroplasts and subchloroplast fragments enriched in system I or II. Photochem. Photohlol., 26: 33-39.
Trebst. A. (1979). Inhibition of photosynthetic electron flow by phenol and diphenyl ether herbicides in control and tripsin treated chloroplasts. Z. Natur/orsch., 34c: 986-991.
Trepst, A. (1981). Action mechanism of herbicides in photosynthetic electron transport. In: Proc. 5th Int. Congo on Photosynthesis. (G. Akoyunoglou ed.). Greece.
Trebst. A. and Draber. W. (1979). Structure-activity correlations of recent herbicides in phot
osynthetic reactions. In: Advances in Pest. Sci. Part 2. (Geissbuhler. H. ed.) : 223-234. Pergamon Press. Oxford.
Vasa. I., Horvath, G .• Herczeg. T. and Demeter. S. (1981). Photosynthetic energy conserva tion investigated by thermoluminescence. Biochim. Biophys. Acta., 634: 140-152.