D
E
V
E
LO
P
M
E
N
T
INTRODUCTION
Gastrulation of the mouse embryo culminates in the formation of three primary germ layers: ectoderm, mesoderm and endoderm. These layers contain the precursors of all the tissues of the fetal body (Tam and Behringer, 1997; Lawson, 1999; Tam and Gad, 2004). Fate-mapping studies have revealed a body plan of the embryo that can be discerned from the regionalization of the germ-layer progenitors in the epiblast (Lawson et al., 1991; Lawson and Pedersen, 1992; Quinlan et al., 1995; Parameswaran and Tam, 1995) and the primitive streak (Tam and Beddington, 1987; Kinder et al., 1999; Kinder et al., 2001). Subsequently, the precursors of major tissue types of the body can be discerned at specific locations, with reference to the orientation and polarity of the primary embryonic axes, in the ectoderm (Beddington, 1981; Beddington, 1982; Tam, 1989), the mesoderm (Lawson and Pedersen, 1992; Parameswaran and Tam, 1995; Tam et al., 1997) and the endoderm (Lawson and Pederson, 1987; Lawson et al., 1986; Tam and Beddington, 1992; Tam et al., 2004). The endoderm, which is the focus of the present study, contains the precursors that give rise to the epithelium of both the gut and the associated organs, such as the liver, pancreas and the respiratory tract.
Analysis of the developmental fates of cells in the endoderm layer of the pre-streak mouse embryo has shown that they contribute almost exclusively to the extraembryonic endoderm. However, after gastrulation commences, some cells in the endoderm in the vicinity of the newly formed primitive streak are fated to become the gut endoderm of the early-somite embryo (Lawson and Pedersen, 1987). By quantifying the epiblast-derived
cells in the endoderm, it was estimated that approximately 5% (about 400 cells) of the total population of the endoderm is recruited in the first 4 hours after the onset of gastrulation and that this incoming population rises to 10% of the total population by the mid-streak (MS) stage (Tam and Beddington, 1992). The majority of the newly recruited cells were localized in the endoderm in the vicinity of the anterior end of the primitive streak where trafficking of epiblast-derived cells to the endoderm takes place (Tam and Beddington, 1992). At the MS stage, endoderm cells in the distal region of the embryo, as well those overlying the primitive streak, are fated to become foregut and ‘posterior’ endoderm (Lawson et al., 1986), but there seems to be no precursors for the ‘mid-gut’ endoderm. This raises the issue of whether the allocation of the endoderm to different segments of the gut follows the same anterior-posterior order of allocation as that of mesodermal derivatives (Tam and Tan, 1992; Kinder et al., 1999).
In the present study, the contribution of cells from different regions of the endoderm of mid-gastrula-stage embryos to different segments of the embryonic gut of early-somite-stage embryos was mapped. Previous analyses on gastrula embryos mainly focused on endodermal cells along the anterior-posterior body axis (Lawson et al., 1986; Lawson and Pedersen, 1987) and did not encompass cells in the non-axial regions of the endoderm layer. Our goal is to elucidate the developmental fates of the entire endoderm population of the MS embryo to achieve a comprehensive fate map of the endoderm that is comparable in its coverage to that of the no- to early-bud-stage embryo (Tam et al., 2004). We have tracked the localization of the descendants of a group of cells that can be visualized by the expression of fluorescent protein tags (achieved by electroporation of expression vectors) or by the emission signal of lipophilic flurochrome (by painting the surface of the cells). The use of these vital cell markers also enables the elucidation of the overall pattern of cell movement in the endoderm of live embryos. Our results reveal that the allocation of definitive endoderm begins with precursors of the rostral-most foregut and the most-posterior segment of the embryonic gut, followed by the rest of the foregut and the mid- and hind-gut. The newly recruited definitive endoderm
Sequential allocation and global pattern of movement of
the definitive endoderm in the mouse embryo during
gastrulation
Patrick P. L. Tam1,2,*, Poh-Lynn Khoo1, Samara L. Lewis1, Heidi Bildsoe1, Nicole Wong1, Tania E. Tsang1, Jacqueline M. Gad1,‡and Lorraine Robb3
During mouse gastrulation, endoderm cells of the dorsal foregut are recruited ahead of the ventral foregut and move to the anterior region of the embryo via different routes. Precursors of the anterior-most part of the foregut and those of the mid- and hind-gut are allocated to the endoderm of the mid-streak-stage embryo, whereas the precursors of the rest of the foregut are recruited at later stages of gastrulation. Loss of Mixl1function results in reduced recruitment of the definitive endoderm, and causes cells in the endoderm to remain stationary during gastrulation. The observation that the endoderm cells are inherently unable to move despite the expansion of the mesoderm in the Mixl1-null mutant suggests that the movement of the endoderm and the mesoderm is driven independently of one another.
KEY WORDS: Definitive endoderm, Allocation, Movement, Mixl1, Gastrulation, Mouse embryo Development 134, 251-260 (2007) doi:10.1242/dev.02724
1Embryology Unit, Children’s Medical Research Institute and 2Faculty of Medicine, University of Sydney, Locked bag 23, Wentworthville, New South Wales 2145, Australia. 3The Walter and Eliza Hall Institute of Medical Research, 1G, Royal Parade, Parkville, Victoria 3050, Australia.
*Author for correspondence (e-mail: [email protected])
‡Present address: Kolling Institute of Medical Research, Royal North Shore Hospital, St Leonards, NSW 2065, Australia
D
E
V
E
LO
P
M
E
N
T
rapidly expands to displace the pre-existing visceral endoderm to extraembryonic sites. In addition, we studied the movement of cells in the Mixl1-null embryo, which is deficient of definitive endoderm. Based on the results of this analysis, we postulate that the accretion of cells via recruitment from the epiblast and the primitive streak may produce the propulsive force that drives the anterior expansion of the definitive endoderm during gastrulation.
MATERIALS AND METHODS
Mice
Two strains of mice were used for gastrula-stage endoderm-fate analysis: out-bred albino ARC/s mice, which provided the embryos for the fate-mapping studies, and Mixl1+/GFPmice generated by targeting an inframe
GFP transgene to the Mixl1locus, which resulted in the deletion of exon 1 of the gene (Hart et al., 2002). A similar strategy was used to generate a lacZ
knock-in Mixl1allele (L.R., unpublished), which provides another reporter of Mixl1activity to compare with the GFP reporter. Mixl1+/GFPmice were
inter-crossed to obtain embryos of Mixl1+/GFPand Mixl1GFP/GFPgenotypes.
To obtain donor cells for the fate-mapping study, a new stock of
Mixl1+/GFP;lacZmice was produced by crossing Mixl1+/GFPmice with
transgenic mice that express the Hmgcr-lacZtransgene (Tam and Tan, 1992).
Mixl1+/GFP;lacZ mice were inter-crossed to produce Mixl1+/+,Mixl1+/GFP
and Mixl1GFP/GFPembryos that also widely express the lacZtransgene in all
embryonic cell types. The Mixl1+/GFPand Mixl1GFP/GFPembryos were the
donors of cells for the transplantation experiment for testing the endodermal potential of primitive-streak cells. ARC/s strain embryos were used as recipients of cell transplantation.
Embryo culture
Embryos were harvested from pregnant mice at 7.0 days post coitum (E7.0). They were dissected from the uterus and the decidua. Following the removal of the Reichert’s membrane, embryos were sorted into the MS and mid- to late-streak (M-LS) stages based on their morphology (Downs and Davies, 1993). Prior to experimental manipulation, embryos were kept in DR75 culture medium (Sturm and Tam, 1993) comprised of 75% heat-inactivated rat serum and 25% Dulbecco’s modified Eagle medium. The culture medium was kept at 37°C under 5% CO2, 5%O2, 90% N2in a bottle rotating at 30
RPM. After manipulation (electroporation, painting or cell transplantation), embryos were cultured in the DR75 medium for up to 48 hours.
Fate mapping by whole-embryo electroporation
The fates of the endoderm of MS and M-LS embryos were mapped by tracing the distribution of EGFP-tagged cells after development to the early-head-fold (EHF) stage (24 hours of culture) and early-somite stage (40-46 hours of culture). Cells in the endoderm of gastrula-stage mouse embryos were marked by introducing either a CMV-EGFPor CMV-lacZ-IRES2-EGFPexpression vector into the embryos via electroporation. The embryos were soaked for 5 minutes in an aqueous plasmid solution (1-1.5
g/l DNA) and were then washed in pH 7.5 Tyrode Ringer solution to remove the excess DNA that had not been adsorbed on the apical surface of the endoderm cells. The embryos were then positioned between a plate and a needle platinum electrode (Davidson et al., 2003). Electroporation was performed using a BTX Electro Square Porator T820 that delivers with a 15 V charging voltage 5 square-wave pulses for 50 milliseconds each with a 1-second inter-pulse gap at a low voltage mode. The fluorescent EGFP-expressing cells were visualized using a Leica MZ16 FA fluorescence stereomicroscope. The image data were captured by a SPOT 2 Slider digital camera and editing was performed using SPOT 32 application software and Adobe Photoshop CS. The lacZ-expressing cells were visualized by X-gal staining of specimens fixed in 4% paraformaldehyde for 5 minutes at the end of the culture experiment. The stained embryos were processed for histology to localize the X-gal-stained cells in the embryonic tissue.
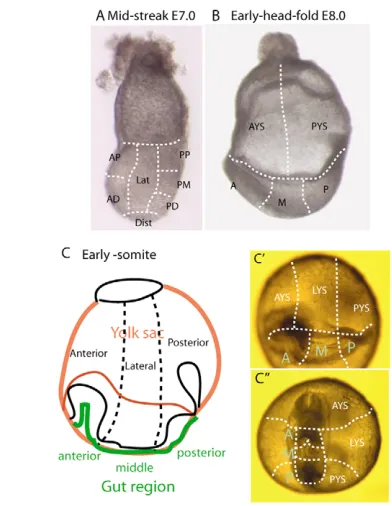
Cells at seven sites of the endoderm were studied (Fig. 1A). A total of six sites were localized along the anterior-posterior axis: two in the anterior region (anterior-proximal and anterior-distal domains), one in the distal region and three in the posterior endoderm (posterior-distal, posterior-middle and posterior-proximal domains). The area between the anterior and
[image:2.612.325.521.58.311.2]posterior regions was designated as the lateral site. The site of electroporation was ascertained by the localization of EGFP-expressing cells after 3 hours of in vitro development. The distribution of EGFP-expressing endoderm cells was monitored after 24 hours of in vitro culture when the embryo had developed to the EHF (equivalent to E8.0) stage (Fig. 1B). For scoring the location and number of EGFP-expressing cells, the yolk sac was partitioned into the anterior and posterior halves, and the embryonic endoderm was subdivided into anterior (underneath the anterior half of the head folds), middle (the posterior half of the head folds) and posterior (from the posterior margin of the head folds to the posterior end of the primitive streak) regions (Fig. 1B). After fluorescence imaging and digital photography, embryos were cultured for another 24 hours, during which time they developed to the early-somite stage, which is equivalent to about E8.75 in vivo. The distribution of EGFP-expressing cells was scored in three regions (anterior-, lateral- and posterior-third) of the yolk sac, and in the anterior (in the foregut portal and ventral to the heart), middle (the somites) and posterior (the presomitic mesoderm) regions of the embryo (Fig. 1C⬘ -C⬙). The number of EGFP-expressing cells in these regions was scored (for details, see Tam et al., 2004). For estimating the relative contribution of cells that were derived from the endoderm in different regions of the MS and M-LS embryo, the data were presented as percentage of the total population (Table 1).
Fig. 1. Sites of electroporation and domains in the endoderm for fate mapping experiments. (A) Regions of the endoderm for testing cell fates by electroporation: anterior-proximal (AP), anterior-distal (AD), distal (Dist), lateral (Lat), posterior-distal (PD), posterior-middle (PM) and posterior-proximal (PP). (B) Regions of the endoderm for scoring the distribution of GFP-expressing cells in early-head-fold-stage embryos. (C-C⬙) Early-somite-stage embryo. The sub-division of the endoderm is shown as a schematic lateral view (C), an oblique view (C⬘) and a ventral view (C⬙). In the early-somite-stage embryo, although the domain of embryonic foregut may be approximately delineated by the foregut portal, the precise sub-division of gut segments is not
D
E
V
E
LO
P
M
E
N
T
Tracking the endoderm by painting cells
Cells in the endoderm of MS- and M-LS-stage ARC/s, Mixl1+/GFPand
Mixl1GFP/GFPembryos were labeled by painting the with carbocyanine dyes: DiO (D275), CM-DiI (C-7001) or DiI (D282, D3911, Molecular Probes). While holding the embryo by suction against a wide-bored pipette with polished tip, the endoderm was painted by touching the cells with a bolus of dye that was partially extruded from the tip of another micropipette. A broad area of the endoderm of the ARC/s embryos was painted in single color with either DiI (red) or DiO (green), or in both of these colors by painting consecutively with DiI and DiO. Painting was performed using DiI on the
Mixl1+/GFPand Mixl1GFP/GFPembryos to contrast labeled cells with the green fluorescent host tissues. Painted embryos were imaged by fluorescence microscopy and digital photography 1 hour after labeling to determine the paint pattern; and at 6-, 12- or 24-hours in vitro to visualize the distribution of labeled endoderm cells in the embryonic gut and the yolk sac.
Testing endoderm potential by cell transplantation
Pregnant Mixl1+/GFP;lacZmice were euthanized at E7.0 to harvest MS-stage
embryos, which provided the cells for the transplantation experiment. The embryos were examined under the fluorescence microscope and genotyped based on the intensity of GFP fluorescence: Mixl1+/+, no green signals; Mixl1+/GFP, moderate to weak green fluorescence; Mixl1GFP/GFP, strong
green fluorescence. In addition, because Mixl1is expressed specifically in the primitive streak (Robb et al., 2000; Hart et al., 2002), the relative size of the GFP or lacZ-expression domain (see Fig. S1A,B in the supplementary material) enables the staging of gastrulation and guides the identification of the anterior part of the primitive streak for harvesting cells (see Fig. S1C in the supplementary material). Donor embryos were dissected by polished alloy metal needles to isolate small fragments of tissues from the anterior segment of the primitive streak and the adjacent epiblast, from which any adherent mesoderm and endoderm were removed as completely as possible. The fragments were then dissociated into clumps of 10-15 cells using glass needles. These cell clumps were transplanted using Leica mechanical micromanipulators to the anterior segment of the primitive streak of stage-matched MS ARC/s embryos. Within 1 hour after transplantation, the
recipient embryo was examined to ascertain the presence and proper location of the GFP-expressing grafted cells (see Fig. S1D in the supplementary material).
After 24 hours of in vitro development to the early-somite stage, the recipient embryos were imaged by fluorescence microscopy to visualize the distribution of the GFP-expressing cells (see Fig. S1E in the supplementary material). They were then fixed in 4% paraformaldehyde and stained with X-gal reagent to visualize the Hmgcr-lacZ-expressing cells (see Fig. S1E in the supplementary material). Embryos containing positively stained graft-derived cells were processed and examined by histology. The number and distribution of the graft-derived cells in the tissues of the host embryo, especially in the endodermal derivatives, were scored in serial histological sections of the specimen.
RESULTS
Precursors of definitive endoderm are localized to the posterior and distal sites
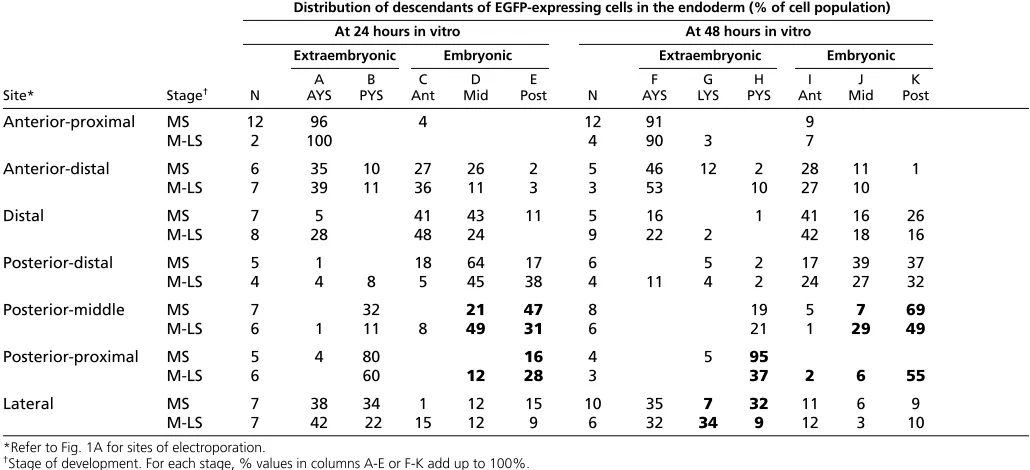
[image:3.612.48.563.84.319.2]To map the fate of endodermal cells in the MS embryo, whole embryos were electroporated with a plasmid encoding EGFP and the distribution of the fluorescent cells was monitored after development to EHF stage and early-somite stage (Table 1). In the MS embryo, precursors of the definitive endoderm of the early-somite embryo (48 hours in vitro) constituted approximately 47% of the endodermal population. Precursors of anterior (foregut) endoderm were found in the distal (anterior-distal, distal and posterior-distal) domains (Fig. 2C,F,G; Table 1, column I of MS embryo). Precursors of middle endoderm (at the somite level) were also localized to distal sites (anterior-distal, distal and posterior-distal; Fig. 2F; Table 1, column J of MS embryo). Precursors of the posterior endoderm of early-somite-stage embryos were found in the distal-to-posterior region (distal, posterior-distal and posterior-middle; Fig. 2E; Table 1, column K of MS embryo). Therefore, precursors of the different parts of the embryonic gut endoderm at the early-somite stage are found predominantly in the anterior-distal, distal, posterior-distal and Table 1. Distribution of GFP-expressing cells in the endoderm of embryos cultured for 24 hours or 40-44 hours after
electroporation
Distribution of descendants of EGFP-expressing cells in the endoderm (% of cell population)
At 24 hours in vitro At 48 hours in vitro
Extraembryonic Embryonic Extraembryonic Embryonic
A B C D E F G H I J K
Site* Stage† N AYS PYS Ant Mid Post N AYS LYS PYS Ant Mid Post
Anterior-proximal MS 12 96 4 12 91 9
M-LS 2 100 4 90 3 7
Anterior-distal MS 6 35 10 27 26 2 5 46 12 2 28 11 1
M-LS 7 39 11 36 11 3 3 53 10 27 10
Distal MS 7 5 41 43 11 5 16 1 41 16 26
M-LS 8 28 48 24 9 22 2 42 18 16
Posterior-distal MS 5 1 18 64 17 6 5 2 17 39 37
M-LS 4 4 8 5 45 38 4 11 4 2 24 27 32
Posterior-middle MS 7 32 21 47 8 19 5 7 69
M-LS 6 1 11 8 49 31 6 21 1 29 49
Posterior-proximal MS 5 4 80 16 4 5 95
M-LS 6 60 12 28 3 37 2 6 55
Lateral MS 7 38 34 1 12 15 10 35 7 32 11 6 9
M-LS 7 42 22 15 12 9 6 32 34 9 12 3 10
*Refer to Fig. 1A for sites of electroporation.
†Stage of development. For each stage, % values in columns A-E or F-K add up to 100%. For site of labeled cells see Fig. 1C.
Boldfont indicates the region where noticeable changes in the relative contribution of cells were found between MS- and M-LS-stage embryos.
D
E
V
E
LO
P
M
E
N
T
posterior-middle regions of the MS embryo (Fig. 3). These regions are within the Cer1-expression domain and partially overlap the expression domain of Sox17, a definitive endoderm marker (Fig. 3A). In the MS embryo, precursors of the anterior yolk sac (AYS) endoderm were derived from the anterior-to-distal sites (anterior-proximal, Fig. 2B; anterior-distal and distal, Table 1, column F of MS embryo). The precursor of the lateral yolk sac (LYS) endoderm was in the anterior-distal and lateral sites (Table 1, column G of MS embryo). The posterior yolk sac (PYS) endoderm was derived from lateral (Fig. 2A), posterior-middle and posterior-proximal (Fig. 2D) sites (Table 1, column H of MS embryo). Precursors of the extraembryonic (yolk sac) endoderm are therefore localized mainly in the anterior (proximal to distal) and posterior proximal regions of the endoderm (Fig. 3B⬙). There are, however, overlaps in the domains of precursors of the extraembryonic endoderm and the definitive endoderm. Specifically, precursors of AYS and anterior gut endoderm are co-localized in the anterior-distal and distal regions, as are those of the PYS and posterior gut endoderm in the posterior-middle region (Fig. 3B⬘,B⬙; Table 1, columns F-K for the anterior-distal, distal and posterior-middle sites).
Histological examination of the MS embryo (n=8) electroporated with the CMV-EGFP-IRES2-lacZ expression vector (Fig. 2H) confirmed that only endodermal cells were labeled (Fig. 2I). At the end of in vitro culture, lacZ-expressing cells (Fig. 2J) were localized in only the endoderm in the early-somite-stage embryo (n=8; foregut: Fig. 2K; midgut: Fig. 2L).
Expanding occupancy of the gut-endoderm precursors during gastrulation
[image:4.612.52.383.296.730.2]To test whether the localization of the precursor population changes during gastrulation, another snapshot of the regionalization of cell fates in the endoderm was taken at the M-LS stage, which is around 6 hours more advanced than the MS stage. The most noticeable difference was the relative contribution of cells in the posterior-proximal and lateral endoderm to the yolk sac. There was a reduced contribution to the PYS of cells at these two sites in the M-LS embryo (Table 1, column H, bold font), whereas the contribution to the LYS from the lateral site was increased (Table 1, column G, bold font). These changes in the relative abundance of the yolk sac precursors at specific endoderm sites suggest that, during
D
E
V
E
LO
P
M
E
N
T
gastrulation from the MS to the M-LS stage, the PYS precursors exit the posterior-proximal site, and that the AYS and PYS precursors that previously occupied the lateral site have departed and their place has been taken up by the precursor of the LYS.
The departure of the yolk sac-endoderm precursors led to a slight increase (by 5% to 52.7%) in the relative amount of gut-endoderm precursors in the M-LS embryo. The most obvious changes were the increase in the abundance of gut endoderm precursors in the posterior-proximal site (Table 1, column I-K of M-LS embryo, bold font). This was accompanied by a decrease in the precursor of the posterior endoderm and an increase in the middle (somite-level) endoderm in the posterior-middle site (Table 1, column J, K of MS and M-LS embryo, bold font). This pattern of cell movement was already evident from the changes in the relative distribution of the embryonic-gut-endoderm precursors in the posterior-middle and posterior-proximal regions of the embryos examined after 24 hours of in vitro development (Table 1, column D, E of MS and M-LS embryos, bold font). Based on the changes in the relative abundance of precursor types (Table 1, columns F-K), a pattern of displacement can be constructed to reflect the movement and regional expansion of the precursor populations from the seven sites (Fig. 3C) to the extraembryonic endoderm during the development from the MS to the M-LS stage (Fig. 3D), and to the EHF stage (Fig. 3E,E⬘). Similarly,
corresponding maps of the morphogenetic movement of precursors of the gut endoderm are shown in Fig. 3F-G⬘. During gastrulation, the pre-existing cell populations moved from the anterior, lateral and posterior proximal sites to the extraembryonic endoderm. This was accompanied by an expansion of the abundance of precursors of the definitive endoderm, predominantly towards the anterior and distal regions. This coincides with the distal extension of the primitive streak (see Fig. S1A,B in the supplementary material).
Cellular recruitment to the endoderm may drive cell movement
[image:5.612.202.562.327.704.2]The re-construction of the pattern of cell movement in the endoderm based on the regionalization of cell fate implies that cell movement may be associated with the continuous accretion of cells to the endoderm from the primitive streak during gastrulation. To test this hypothesis, we studied the Mixl1-null-mutant embryos, which were shown to be deficient of Sox17- and Cer1-expressing gut endoderm, presumably resulting from an inability to direct the allocation of mesendodermal progenitors to the definitive endoderm (Hart et al., 2002). We first tested whether Mixl1-null-mutant cells may have an impaired potential to form endoderm by cell-transplantation experiments. Cells from the anterior region of the primitive streak (see Fig. S1C in the supplementary material) of Mixl1+/GFP
Fig. 3. Fate maps of the precursors of embryonic and extraembryonic endoderm and the pattern of cell movement in the endoderm of the mid-streak stage embryo.(A) The expression domain of Cer1 and Sox17in the
endoderm of a mid-streak embryo (anterior to the left). Cer1expression overlaps in the distal, posterior-distal and posterior-middle regions, which contribute to the gut endoderm, but is also expressed in the endoderm immediately distal and lateral to the primitive streak, and in the anterior proximal endoderm. Sox17expression overlaps in the posterior-middle region and is also expressed in the extraembryonic visceral endoderm. Both genes are expressed in the anterior visceral endoderm (asterisk, anterior proximal region). (B) Fate maps of the endoderm of mid-streak-stage gastrula (left figures in B⬘and B⬙) showing the localization of the precursors of embryonic (gut) endoderm (B⬘) and extraembryonic (yolk sac) endoderm (B⬙) of the early-somite-stage embryo (right figures in B⬘and B⬙). (C) Color coding of the seven sites in the mid-streak-stage embryo for testing endodermal cell fates (see Fig. 1A). (D-G’) The trajectories of the progenitors of the yolk sac endoderm (D-E’) and
embryonic endoderm (F-G’) during development from the mid-streak to the mid- to late-streak stage (D,F) and then to the early-head-fold stage (E,E’,G,G’), showing the distribution of cells arising from (E,G) anterior and distal sites and (E’,G’) posterior sites separately. Cells originated from each of the seven sites are
D
E
V
E
LO
P
M
E
N
T
(Mixl1+/–, n=15) andMixl1GFP/GFP(Mixl1–/–, n=47) embryos were transplanted orthotopically to the ARC/s (wild type) embryos (see Fig. S1C in the supplementary material). Mixl1+/–and Mixl1–/–cells were similar in their ability to multiply and colonize the host tissues (Table 2, and see Fig. S1F,G in the supplementary material). However, histological examination and cell-count analysis revealed that Mixl1–/–primitive-streak cells contributed less to the endoderm than Mixl1+/–cells and more to the mesoderm of the host embryo than Mixl1+/–cells (Table 2). This strongly suggests that the loss of Mixl1function has an impact on the endoderm potential of the mesendoderm progenitors, which are presumably found in the primitive streak (D’Amour et al., 2005; Yasunaga et al., 2005; Tada et al., 2005; Ng et al., 2005; Loebel and Tam, 2005). The reduced contribution to the endoderm was apparent even when the Mixl1 -deficient cells had been in an environment conducive to endoderm formation. The deficiency of definitive endoderm is therefore likely to be caused by the impaired recruitment of cells to the endoderm during gastrulation (Hart et al., 2002).
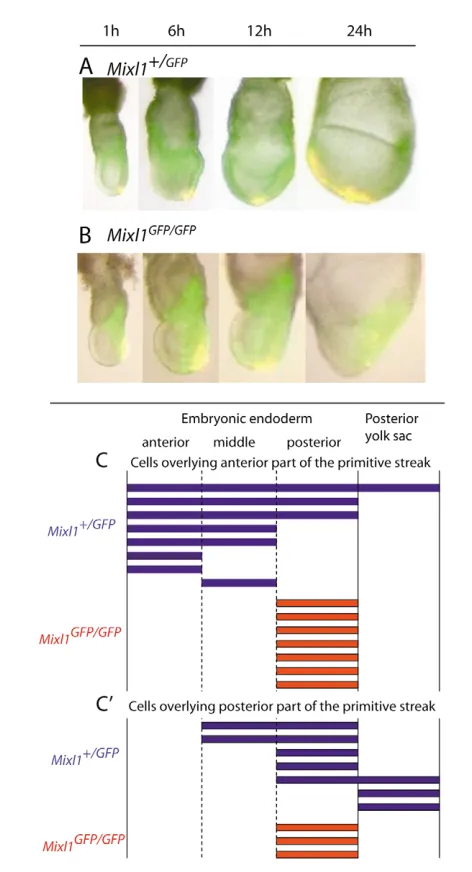
In Mixl1GFP/GFPembryos, the allocation and differentiation of the mesoderm were unaffected. Although the anterior expansion of the mesoderm appeared to be delayed, a complete mesodermal layer was formed eventually (see Fig. S2A,B in the supplementary material). To test whether the movement of the endoderm was also retarded in the Mixl1GFP/GFPembryo, endoderm cells overlying the anterior half of the primitive streak were painted with DiI and their position was determined after 24 hours of in vitro culture. In the Mixl1+/GFPembryo (n=14), labeled cells were distributed along the anterior-posterior axis whereas, in the Mixl1GFP/GFP embryo (n=11), labeled cells stayed in the posterior region (see Fig. S2C,D in the supplementary material). By tracking the
movement of GFP-positive mesoderm and DiI-labeled endoderm in the embryos at different time points of in vitro development, the endoderm cells in the Mixl1GFP/GFPwere found to remain stagnant while the mesoderm spread anteriorly (Fig. 4A,B). The impaired movement was not limited to the endoderm associated with the anterior half of the primitive streak, because cells overlying the posterior half of the primitive streak of Mixl1GFP/GFP embryos also remained stationary (Fig. 4C). Therefore, the endoderm in the Mixl1-null mutant is inherently unable to move and could not be mobilized by the expansion of the underlying mesoderm.
Allocation of the lateral and medial gut endoderm
[image:6.612.49.561.81.368.2]The transplantation experiment showed that cells derived from the primitive streak contribute to the gut endoderm in the anterior and middle region of the EHF-stage embryo (Table 2). To elucidate whether there was a sequential order of allocation of medial and lateral populations of the gut endoderm in these regions, the fate of cells in the endoderm at the anterior region of the primitive streak of embryos at the MS and M-LS stages was examined. Endoderm at the anterior region of the primitive streak of Mixl1+/GFPMS (Fig. 5A, 0 hour) and M-LS embryos (Fig. 5B, 0 hour) was painted with DiI, and the distribution of the labeled cells was examined at 12 hours (late-streak stage) and 24 hours (EHF stage) of in vitro culture. Labeled endoderm cells of MS-stage embryos were first present in the lateral proximal region of the embryo (Fig. 5A, 12 hours) and finally in the lateral region of anterior and middle endoderm (Fig. 5A, 24 hours). By contrast, the endoderm cells from a similar site in the M-LS-stage embryo (Fig. 5B, 0 hour) were Table 2. Contribution of the anterior primitive-streak cells of mid- to late-streak stage Mixl1+/–and Mixl–/–mutant embryos to the germ layer derivatives of the wild-type host embryo
Donor cells (genotype) Mixl1+/– (Mixl1+/GFP;Hmgcr-lacZ) Mixl1–/– (Mixl1GFP/GFP;Hmgcr-lacZ)
Donor cells: tissue of origin / stage APS / M-LS APS / M-LS
Site of transplantation APS APS
Number of recipient embryos analyzed 14 32
Tissue distribution Cell count % total Cell count % total
Gut endoderm
Anterior 48 8.3 45 2.8
Middle 48 8.3 14 0.9
Posterior 6 1.0 0 0
Axial mesoderm
Head process 5 0.9 0 0
Notochord 45 7.8 122 7.5
Neural tissues of the trunk
Neural tube 0 0 35 2.2
Floor plate 0 0 58 3.6
Paraxial mesoderm
Cranial 83 14.3 671 41.5
Somite 78 13.4 386 23.8
Presomitic 153 26.4 60 3.7
Lateral mesoderm 12 2.1 132 8.3
Primitive streak 102 17.6 93 5.7
Total number of cells scored 580 1616
Average number of graft-derived cells/embryo 41.4 50.5
Bold font indicates noticeable differences between the contribution of the Mixl–/–cells to specific types of host tissues and that of Mixl1+/–cells.
D
E
V
E
LO
P
M
E
N
T
distributed initially along the axial and paraxial region (Fig. 5B, 12 hours) and subsequently moved to the medial region of the anterior and middle endoderm (Fig. 5B, 24 hours). In summary, precursors of the lateral-anterior and middle endoderm, which constitute most of the foregut endoderm (Tremblay and Zaret, 2005), may be allocated separately from those of the medial endoderm, and these two endoderm populations reach their destination via different paths (Fig. 5C). During the morphogenesis of the foregut portal, the lateral and medial population will become the ventral and dorsal endoderm of embryonic gut, respectively (Fig. 5D). These results
reveal that the timing of recruitment to the endoderm influences the allocation of cells to the medial (dorsal) and lateral (ventral) endoderm of the gut.
Trajectories of gut-endoderm precursors to the embryonic gut
[image:7.612.50.286.49.485.2]The trajectory of gut-endoderm precursors of MS-stage embryos to the endoderm of different parts of the embryonic gut of EHF-stage embryos was tracked by mapping the distribution of cells originated from five areas (Fig. 6A) covering the whole of the posterior and Fig. 4. Impaired cell movement in the endoderm of Mixl1GFP/GFP
mutant embryos.(A,B) The displacement of DiI-labeled endoderm (yellow because of overlaying of red dye color on the green
fluorescence) in Mixl1+/GFP(A) and Mixl1GFP/GFP(B) mutant embryos after
1-, 6-, 12- and 24-hours of in vitro culture. (C,C⬘) Summary of the distribution of DiI-labeled endodermal cells along the anterior-posterior axis of the embryos and in the posterior yolk sac 24 hours after labeling. Each bar represents the pattern of distribution of labeled cells in a single representative embryo after the endoderm overlying the anterior (C) or the posterior (C⬘) region of the primitive streak was painted.
[image:7.612.307.559.58.414.2]D
E
V
E
LO
P
M
E
N
T
distal endoderm. Cells anterior to the primitive streak (distal and posterior distal) and those at anterior end of the primitive streak (APS) contributed to the endoderm from the prechordal to the somitic levels, but rarely to the posterior (presomitic mesoderm) endoderm (Fig. 6B,C, and see Table S1 in the supplementary material). Cells in the middle segment of the primitive streak (posterior middle) contributed exclusively to the endoderm from the hindbrain to the presomitic mesoderm level, but not to that in the prechordal to midbrain level. The endoderm in the posterior segment of the primitive streak (posterior distal) colonized only the posterior-most endoderm (see Table S1 in the supplementary material).
Although the endoderm associated with the primitive streak can contribute to the embryonic endoderm along the whole anterior-posterior body axis of the EHF embryo (see Table S1 in the supplementary material), the most preponderant contribution is to the anterior endoderm and the most-posterior endoderm. Furthermore, within the anterior endoderm, most labeled cells were found in the anterior-lateral and posterior endoderm, and fewer were found the middle endoderm (Fig. 6D). To reveal the source of anterior medial endoderm, double-color painting of the posterior-distal to the posterior-proximal endoderm was performed, with an overlapping area in the APS (yellow area in Fig. 6E). The result showed that the medial population of the anterior and middle endoderm was derived from cells in the posterior-distal (red cells in Fig. 6E) and APS (yellow cells in Fig. 6E), whereas the lateral
anterior endoderm and the posterior endoderm were derived from the APS and posterior-proximal cells (green cells in Fig. 6E). APS cells contributed to the medial population of both the anterior and middle endoderm of the embryo. Endoderm cells in a large area of the middle and posterior regions were not labeled (Fig. 6E), suggesting that they may be recruited to the endoderm after the labeling at the MS stage. The allocation of precursors of definitive endoderm during gastrulation therefore commences with the anterior and the most-posterior endoderm, followed by those of middle endoderm and the rest of posterior endoderm. Histological examination of the dye-labeled embryos revealed that labeling was limited to the superficial endoderm layer (Fig. 6F, three embryos examined 1 hour after labeling) and the gut endoderm (Fig. 6G-I, four out of six embryos examined after 24 hours of culture). However, in two of the six embryos examined at 24 hours, although the majority of labeled cells was in the endoderm, a few were also found in the mesoderm, as previously reported by Lawson and Pedersen (Lawson and Pedersen, 1987).
DISCUSSION
Progressive allocation of the definitive endoderm during gastrulation
[image:8.612.50.374.59.437.2]Fate mapping of the endoderm of MS embryos revealed that the fate of cells in the area distal to the anterior region of the primitive streak is to become the medial population of the anterior endoderm (the dorsal foregut). The location of some dorsal-foregut endoderm
D
E
V
E
LO
P
M
E
N
T
precursors distal to the anterior primitive streak (Lawson and Pedersen, 1987) (this study) may be due to the displacement of precursors that had been recruited since the early-streak stage. The preponderance of precursors of the lateral population of the anterior endoderm (the ventral foregut) in the vicinity of the primitive streak of the mid- to late-streak embryo suggests that these precursors are recruited after the medial population. Recruitment of the endoderm of the foregut therefore follows a temporal schedule of early recruitment of the medial (dorsal) endoderm and later recruitment of the lateral (ventral) endoderm.
Previous analyses of cell fates in the primitive streak of the chick embryo showed that, at Stage 3, cells emerging from the rostral region of the streak contribute to a wide mediolateral domain of the anterior endoderm, whereas those emerging later at Stage 3+ and Stage 4 contribute to the dorsal and ventral midline of the foregut (Lawson and Schoenwolf, 2003; Kirby et al., 2003). A slightly different order of appearance of endoderm types was revealed by mapping the fate of cells in the lower layer of the chick gastrula embryo. In Stage 2 to Stage 3+ (early- to mid-primitive-streak stage) gastrula, the lower layer contains the precursors of the mid-hindgut and dorsal foregut, and they are clustered around the anterior region of the primitive streak (Kimura et al., 2006). As gastrulation proceeds to Stage 4 (definitive-streak stage), precursors of the lateral foregut and more of those for the mid- and hind-gut appear in the lower layer. In the chick gastrula, in contrast to the mouse, precursors of the ventral foregut appear in the lower layer as late as Stage 5, presumably due to the extended retention of these endodermal cells in the mesodermal layer after their ingression at the primitive streak (Kimura et al., 2006).
Of particular interest is that the fate-mapping study in the chick revealed the presence of a small population of mid- and hind-gut precursors in the lower layer of the Stage 2-3 chick gastrula (Kimura et al., 2006). Similarly, in the endodermal layer of the early-streak mouse embryo, a small number of cells that contribute to trunk and posterior endoderm are found adjacent to the primitive streak (Lawson and Pedersen, 1987). Presently, the precise fate of these cells of the early-streak embryo is not known. In the MS embryo, in addition to precursors of the foregut endoderm, the endodermal layer also contains cells that contribute to the endoderm in the most-posterior region of the early-somite embryo. It has been shown that the descendants of these cells can contribute extensively to the gut from the upper-trunk level to the caudal end in 20- to 23-somite embryos (Tanaka et al., 2005) (Lewis and P.P.L.T., unpublished). If these cells are the descendants of the posterior endoderm precursors of the early- to mid-streak embryo, it would suggest that the endoderm of the mid- and hind-gut is allocated very early in gastrulation and ahead of the bulk of foregut precursors. Because the recruitment of cells to the endoderm appears to have ceased by the early-somite stage (Tam and Beddington, 1987) and the precursors for major segments of the gut are present in the no-to early-bud embryo (Lawson et al., 1986; Tam and Beddingno-ton, 1992; Tam et al., 2004), allocation of the full complement of definitive-endoderm precursor has to be accomplished before the morphogenesis of major parts of the mid- and hind-gut of the embryo takes place.
Together, these observations point to a probable sequence of allocation of the definitive endoderm proceeding with: (a) the most-posterior endoderm and the dorsal endoderm of the rostral segment of the foregut at early-streak stage; (b) the ventral endoderm of the rostral foregut and additional posterior endoderm at the MS stage; (c) the dorsal and then the ventral endoderm of the posterior segment of the foregut at the late-streak to late-bud stage; and, finally, (d) the
endoderm of the embryonic mid- and hind-gut at the late-bud- to EHF-stage. If no further recruitment of definitive endoderm takes place after the presomite stage (Tam and Beddington, 1987), the mid- and hind-gut (and presumably also the tail gut) would have to be generated by the expansion of all of the precursors that have been allocated to the definitive endoderm shortly after the completion of gastrulation.
Accretion of cells may drive cell movement in the endoderm
During the development from the MS stage to the M-LS stage, precursors of extraembryonic endoderm move proximally towards the ectoplacental pole of the conceptus from the anterior-proximal and posterior-proximal sites (Fig. 3D-E⬘) The precursors of embryonic endoderm display a concerted movement: cells in the posterior-middle to distal region of the MS-stage embryo are displaced anteriorly and proximally to occupy a wider domain in the lateral and posterior regions of the embryo (Fig. 3F-G⬘). It is worth noticing that, whereas the precursors of the anterior definitive endoderm appear to move in step with the mesodermal layer in the chick gastrula (Lawson and Schoenwolf, 2003; Kimura et al., 2006), the dorsal-foregut endoderm in the mouse embryo may have moved more anteriorly than that of the anterior mesoderm by the MS stage (Parameswaran and Tam, 1995; Kinder et al., 1999). This may suggest that the endoderm could move independently of the mesoderm.
The overall pattern of anterior and proximal displacement of the definitive endoderm is reminiscent of that of the visceral endoderm in the pre-gastrulation embryo (Thomas and Beddington, 1996; Rivera-Perez et al., 2003). Movement of the visceral endoderm has been attributed to active cell migration (Srinivas et al., 2004), to the propulsion generated by differential cell proliferation (Yamamoto et al., 2004) or to guidance-mediated Wnt signaling (Kimura-Yoshida et al., 2005). Results of the present study on Mixl1-mutant embryos reveal that a loss of Mixl1function reduces the endoderm potential of primitive-streak cells. This finding is consistent with the lack of contribution by Mixl1–/– ES cells to the gut endoderm of the chimaeric embryo, and the reduced population of Sox17and Cer1 -expressing cells in the mutant embryo (Hart et al., 2002). The inefficient recruitment of cells to the endoderm may lead to the lessening of the flow of cells into the endoderm immediately adjacent to the primitive streak. The finding that the endoderm cells remain stationary in Mixl1–/–-mutant embryos suggests that one of the forces driving endoderm movement might be the propulsion generated by the accretion of cells in the posterior region of the endoderm during gastrulation. A similar mechanism of driving cell movement by differential accretion of cells has been proposed for the visceral endoderm of the mouse embryo before gastrulation (Yamamoto et al., 2004). In Mixl1–/–embryos, endoderm cells overlying the primitive streak remain stagnant while the mesoderm expands. This finding further highlights the independence of the movement of the mesoderm and the endoderm, and that any traction force that might be exerted by the expanding mesoderm is insufficient to mobilize the endoderm cells.
We thank Peter Rowe and David Loebel for comments on the manuscript; and Kirsten Steiner for contributing Fig. 5D. Our work is supported by the National Health and Medical Research Council (NHMRC) of Australia and by James Fairfax. P.P.L.T. is a Senior Principal Research Fellow and L.R. is a Principal Research Fellow of the NHMRC.
Supplementary material
D
E
V
E
LO
P
M
E
N
T
References
Beddington, R. S. P.(1981). An autoradiograpic analysis of the potency of embryonic ectoderm in the 8th day postimplantation mouse embryo. J. Embryol. Exp. Morphol. 64, 87-104.
Beddington, R. S. P.(1982). An autoradiographic analysis of tissue potency in different regions of the embryonic ectoderm during gastrulation in the mouse. J. Embryol. Exp. Morphol. 69, 265-285.
D’Amour, K. A., Agulnick, A. D., Eliazer, S., Kelly, O. G., Kroon, E. and Baetge, E. E.(2005). Efficient differentiation of human embryonic stem cells to definitive endoderm. Nat. Biotechnol. 23, 1534-1541.
Davidson, B. P., Tsang, T. E., Khoo, P. L., Gad, J. M. and Tam, P. P. L.(2003). Introduction of cell markers into germ layer tissues of the mouse gastrula by whole embryo electroporation. Genesis35, 57-62.
Downs, K. M. and Davies, T.(1993). Staging of gastrulating mouse embryos by morphological landmarks in the dissecting microscope. Development118, 1255-1266.
Hart, A. H., Hartley, L., Sourris, K., Stadler, E. S., Li, R., Stanley, E. G., Tam, P. P., Elefanty, A. G. and Robb, L.(2002). Mixl1 is required for axial
mesendoderm morphogenesis and patterning in the murine embryo. Development129, 3597-3608.
Kimura, W., Yasugi, S., Stern, C. D. and Fukuda, K.(2006). Fate and plasticity of the endoderm in the early chick embryo. Dev. Biol. 289, 283-295. Kimura-Yoshida, C., Nakano, H., Okamura, D., Nakao, K., Yonemura, S.,
Belo, J. A., Aizawa, S., Matsui, Y. and Matsuo, I.(2005). Canonical Wnt signaling and its antagonist regulate anterior-posterior axis polarization by guiding cell migration in mouse visceral endoderm. Dev. Cell9, 639-650. Kinder, S. J., Tsang, T. E., Quinlan, G. A., Hadjantonakis, A. K., Nagy, A. and
Tam, P. P.(1999). The orderly allocation of mesodermal cells to the
extraembryonic structures and the anteroposterior axis during gastrulation of the mouse embryo. Development126, 4691-4701.
Kinder, S. J., Tsang, T. E., Wakamiya, M., Sasaki, H., Behringer, R. R., Nagy, A. and Tam, P. P.(2001). The organizer of the mouse gastrula is composed of a dynamic population of progenitor cells for the axial mesoderm. Development 128, 3623-3634.
Kirby, M. L., Lawson, A., Stadt, H. A., Kumiski, D. H., Wallis, K. T., McCraney, E., Waldo, K. L., Li, Y. X. and Schoenwolf, G. C.(2003). Hensen’s node gives rise to the ventral midline of the foregut: implications for organizing head and heart development. Dev. Biol.253, 175-188.
Lawson, A. and Schoenwolf, G. C.(2003). Epiblast and primitive-streak origins of the endoderm in the gastrulating chick embryo. Development130, 3491-3501. Lawson, K. A.(1999). Fate mapping the mouse embryo. Int. J. Dev. Biol. 43,
773-775.
Lawson, K. A. and Pedersen, R. A.(1987). Cell fate, morphogenetic movement and population kinetics of embryonic endoderm at the time of germ layer formation in the mouse. Development101, 627-652.
Lawson, K. A. and Pedersen, R. A.(1992). Clonal analysis of cell fate during gastrulation and early neurulation in the mouse. Ciba Found. Symp. 165, 3-21. Lawson, K. A., Meneses, J. J. and Pedersen, R. A.(1986). Cell fate and cell
lineage in the endoderm of the presomite mouse embryo, studied with an intracellular tracer. Dev. Biol. 115, 325-339.
Lawson, K. A., Meneses, J. J. and Pedersen, R. A.(1991). Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development 113, 891-911.
Loebel, D. A. F. and Tam, P. P. L.(2005). A gut feeling. Nat. Biotechnol. 23, 1491-1492.
Ng, E. S., Davis, R. P., Azzola, L., Stanley, E. G. and Elefanty, A. G.(2005). The primitive streak gene Mixl1 is required for efficient haematopoiesis and BMP4-induced ventral mesoderm patterning in differentiating ES cells. Development 132, 873-884.
Parameswaran, M. and Tam, P. P. L.(1995). Regionalisation of cell fate and morphogenetic movement of the mesoderm during mouse gastrulation. Dev. Genet. 17, 16-28.
Quinlan, G. A., Williams, E. A., Tan, S. S. and Tam, P. P. L.(1995). Neuroectodermal fate of epiblast cells in the distal region of the mouse egg cylinder: implication for body plan organization during early embryogenesis. Development121, 87-98.
Rivera-Perez, J. A., Mager, J. and Magnuson, T.(2003). Dynamic
morphogenetic events characterize the mouse visceral endoderm. Dev. Biol. 261, 470-487.
Robb, L., Hartley, L., Begley, C. G., Brodnicki, T. C., Copeland, N. G., Gilbert, D. J., Jenkins, N. A. and Elefanty, A. G.(2000). Cloning, expression analysis, and chromosomal localization of murine and human homologues of a Xenopus mix gene. Dev. Dyn. 219, 497-504.
Srinivas, S., Rodriguez, T., Clements, M., Smith, J. C. and Beddington, R. S. P. (2004). Active cell migration drives the unilateral movements of the anterior visceral endoderm. Development131, 1157-1164.
Sturm, K. and Tam, P. P. L.(1993). Isolation and culture of whole
postimplantation embryos and germ layer derivatives. Methods Enzymol. 225, 164-190.
Tada, S., Era, T., Furusawa, C., Sakurai, H., Nishikawa, S., Kinoshita, M., Nakao, K. and Chiba, T.(2005). Characterization of mesendoderm: a diverging point of the definitive endoderm and mesoderm in embryonic stem cell differentiation culture. Development132, 4363-4374.
Tam, P. P. L.(1989). Regionalisation of the mouse embryonic ectoderm: allocation of prospective ectodermal tissues during gastrulation. Development107, 55-67.
Tam, P. P. L. and Beddington, R. S. P.(1987). The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis. Development99, 109-126.
Tam, P. P. L. and Beddington, R. S. P.(1992). Establishment and organization of germ layers in the gastrulating mouse embryo. Ciba Found. Symp. 165, 27-41. Tam, P. P. L. and Tan, S.-S.(1992). The somitogenetic potential of cells in the
primitive streak and the tail bud of the organogenesis-stage mouse embryo. Development115, 703-715.
Tam, P. P. L. and Behringer, R. R. (1997). Mouse gastrulation: the formation of a mammalian body plan. Mech. Dev.68, 3-25.
Tam, P. P. L. and Gad, J. M.(2004). Gastrulation of the mouse embryo. In Gastrulation(ed. C. D. Stern), pp. 223-262. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Tam, P. P. L., Parameswaran, M., Kinder, S. J. and Weinberger, R. P.(1997). The allocation of epiblast cells to the embryonic heart and other mesodermal lineages: the role of ingression and tissue movement during gastrulation. Development124, 1631-1642.
Tam, P. P. L., Khoo, P. L., Wong, N., Tsang, T. E. and Behringer, R. R.(2004). Regionalization of cell fates and cell movement in the endoderm of the mouse gastrula and the impact of loss of Lhx1(Lim1) function. Dev. Biol. 274, 171-187. Tanaka, S. S., Yamaguchi, Y. L., Tsoi, B., Lickert, H. and Tam, P. P. L.(2005).
IFITM/mil/fragilis family proteins, IFITM1 and IFITM3, play distinct roles in mouse primordial germ cell homing and repulsion.Dev. Cell9, 745-756.
Thomas, P. and Beddington, R. S. P.(1996). Anterior primitive endoderm may be responsible for patterning the anterior neural plate in the mouse embryo. Curr. Biol. 6, 1487-1496.
Tremblay, K. D. and Zaret, K. S.(2005). Distinct populations of endoderm cells converge to generate the embryonic liver bud and ventral foregut tissues. Dev. Biol. 280, 87-99.
Yamamoto, M., Saijoh, Y., Perea-Gomez, A., Shawlot, W., Behringer, R. R., Ang, S. L., Hamada, H. and Meno, C.(2004). Nodal antagonists regulate formation of the anteroposterior axis of the mouse embryo. Nature428, 387-392.