Viral-Human Chimeric Transcript Predisposes Risk
to Liver Cancer Development and Progression
Chi-Chiu Lau,1Tingting Sun,1Arthur K.K. Ching,1Mian He,1Jing-Woei Li,1,2Alissa M. Wong,1Ngai Na Co,1
Anthony W.H. Chan,1Pik-Shan Li,3Raymond W.M. Lung,1Joanna H.M. Tong,1Paul B.S. Lai,4Henry L.Y. Chan,5
Ka-Fai To,1,6,7Ting-Fung Chan,2,*and Nathalie Wong1,6,7,*
1Department of Anatomical and Cellular Pathology, The Chinese University of Hong Kong, Shatin, Hong Kong, China 2School of Life Sciences, The Chinese University of Hong Kong, Shatin, Hong Kong, China
3Transgenic Facility, School of Biomedical Sciences, The Chinese University of Hong Kong, Shatin, Hong Kong, China 4Department of Surgery, The Chinese University of Hong Kong, Shatin, Hong Kong, China
5Department of Medicine and Therapeutics, The Chinese University of Hong Kong, Shatin, Hong Kong, China 6State Key Laboratory in Oncology in South China, The Chinese University of Hong Kong, Shatin, Hong Kong, China 7State Key Laboratory of Digestive Disease, The Chinese University of Hong Kong, Shatin, Hong Kong, China
*Correspondence:[email protected](T.-F.C.),[email protected](N.W.)
http://dx.doi.org/10.1016/j.ccr.2014.01.030
SUMMARY
The mutagenic effect of hepatitis B (HBV) integration in predisposing risk to hepatocellular carcinoma (HCC)
remains elusive. In this study, we performed transcriptome sequencing of HBV-positive HCC cell lines and
showed transcription of viral-human gene fusions from the site of genome integrations. We discovered
tumor-promoting properties of a chimeric LINE1 that, intriguingly, functions as a hybrid RNA.
HBx-LINE1 can be detected in 23.3% of HBV-associated HCC tumors and correlates with poorer patient survival.
HBx-LINE1 transgenic mice showed heightened susceptibility to diethylnitrosamine-induced tumor
forma-tion. We further show that HBx-LINE1 expression affects
b
-catenin transactivity, which underlines a role in
activating Wnt signaling. Thus, this study identifies a viral-human chimeric fusion transcript that functions
like a long noncoding RNA to promote HCC.
INTRODUCTION
Chronic hepatitis B (HBV) infection is a major risk factor for the development of hepatocellular carcinoma (HCC). Chronic HBV carriers are believed to be at 100–200 times increased risk of developing HCC compared to noncarriers (El-Serag and Ru-dolph, 2007). Several direct and indirect mechanisms of the viral induction toward HCC formation have hence been proposed. Long-lasting viral multiplications and expression of HBV proteins are known to stimulate host immune response, leading to liver inflammation, cirrhosis, and ultimately cancer (Chisari and Fer-rari, 1995). On the other hand, a more direct effect of HBV inte-grations on hepatic carcinogenesis has been postulated based on the fact that 85%–90% of HBV-associated HCC tumors
har-bor one or more sites of HBV insertion (Bre´chot et al., 2000). The majority of HCC tissue also exhibits clonal expansion of tumor cells carrying the same integration sites (Gozuacik et al., 2001). Implication of viral integrants in promoting HCC development has thus long been suggested, although details of the induced carcinogenetic changes remain elusive.
Earlier reports of isolating HBV integrants using PCR-based methods suggested that the insertional sites occurred randomly throughout the whole genome, leading to the presumption that there were no preferential sites of integration (Matsubara and Tokino, 1990). More recent high-throughput next-generation sequencing studies have begun to reveal a few recurrent sites of insertion and showed that many HBV integrants actually occur within or near repetitive sequences, such as long interspersed
Significance
The biological consequences of HBV integration incisactivation of gene expression and the functional implications of pro-moting HCC development are still largely unclear. Our whole-transcriptome study describes key features of readthrough transcription of chimera viral-human sequences as a consequence of genome integration. Specifically, we provide evidence of functionality of a long noncoding RNA-like transcript, HBx-LINE1, which confers cancer-promoting properties through Wnt/b-catenin signaling pathway activation. Reports on HBV integrants have often failed to address their biological impacts; our findings establish a clear functional role by which viral insertion can support cancer development. In sum, our data provide pathogenic insights into a hybrid RNA in HCC development and highlight an additional layer of molecular complexity in viral oncogenesis.
nuclear elements (LINEs), short interspersed nuclear elements (SINEs), and Alu (Ding et al., 2012; Fujimoto et al., 2012; Sung et al., 2012). The most common cellular gene associated with viral integration to date is the telomerase reverse transcriptase gene (TERT), where insertions within the gene or upstream of the promoter have been described. The humanTERTencodes the telomerase catalytic subunit, which is an essential compo-nent for maintenance of telomere length. Because aberrant overexpression ofTERTis associated with tumor development (Cohen et al., 2007), it is hypothesized that viral integration in theTERTlocus may confer early clonal advantage during chronic HBV infection.
Notably, recent deep-sequencing studies of HBV integrations in HCC have illustrated findings mostly from whole-genome sequencing (Fujimoto et al., 2012; Jiang et al., 2012) or targeted sequencing of enriched HBV integrants (Ding et al., 2012). These studies all focused on viral genomic integration sites and occa-sionally the effect of such integration on host cellular gene expression. The functional consequences of genome disruption by viral insertions, particularly those associated with repetitive sequences, however, have been minimally addressed and remain to be determined. In this study, we attempted to decon-volute HBV junctures at the whole-transcriptome level with an aim toward characterizing the transcribed viral-human fusions and theciseffect on cellular gene expression from the juxtapo-sition of HBV. To aid our analysis, we developed an algorithm, ViralFusionSeq (VFS) (Li et al., 2013), that allows accurate and unbiased detection of fusions between viral and human se-quences as well as complex fusion events. We examined whether direct transcription of HBV-human sequences could result from the site of genome insertions, and whether viral inte-gration could activate transcription of normally silent sequences such as LINEs and SINEs. Here we also provide detailed charac-terization of the biological functions of a specific chimeric tran-script, HBx-LINE1, that we discovered through the VFS screen. A major goal of this study was to provide evidence for tumori-genic activity of HBx-LINE1 and to identify the mode of action responsible for its oncogenicity.
RESULTS
HBV-Human Chimeric Transcripts
We performed transcriptome sequencing to survey six HBV-positive HCC cell lines for viral integrants. Chimeric viral-human transcripts were identified in four cell lines: HKCI-4, -5B, -7, and -11. A total of 11 flanking cellular sequences was isolated, where more than one chimeric transcript could be detected in each specimen, except HKCI-11 (Table 1). Many of the flanking sequences were from the human intergenic regions that con-tained a portion of a repetitive element such as a LINE or SINE (Table S1available online). Other fusion sequences containing part of cellular genes, including CHIC2, UPF3A, ZNF48,
METAP1, and TPCN2, were detected. Transcript fusion from
the proximal insertion upstream ofTERTcould also be found. Interrupted HBV open reading frames (ORFs) were common in the surface gene (HBs), X gene (HBx), and between X and core (HBc) regions (Table 1).
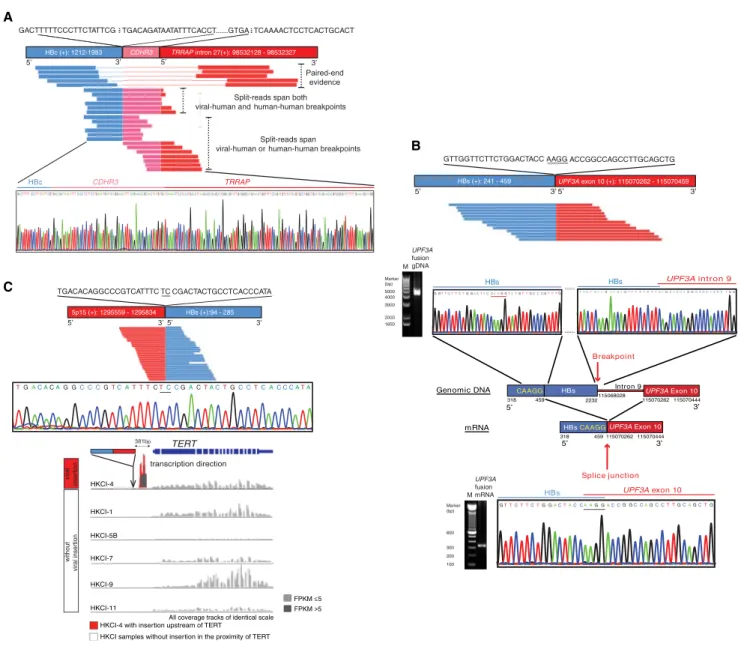
We attempted validation of chimeric sequences and also investigated for their corresponding genomic sites that might underlie the chimeric transcriptions. By Sanger sequencing, we confirmed all VFS-identified viral-human transcripts. Moreover, RNA sequences obtained in the chimeras matched the flanking virus-host juncture of genomic sites with identical overlapping microhomologies (Figure S1A). In addition to directing viral integrations in cellular genes, an integration event in genomic re-gions that have undergone possible complex genomic rear-rangement was found in two cases: a CDHR3-TRRAP fusion and a chr.5q14-RALGPS1fusion (Figure 1A;Figure S1A).
Of particular interest was the finding of a cryptic splicing event resulting from a viral insertion intoUPF3A. In HKCI-4, truncated HBs were transcribed together withUPF3Aexon 10. However, the genomic breakpoints of viral and human sequences that flanked the junctures were dissimilar to the chimeric transcript; instead, HBs DNA (nt 318–2232) was shown to be inserted into intron 9 ofUPF3A. The HBs gene contains a CAAGG sequence (nt 455–459), which is identical to the last 5 nt of UPF3A exon 9. It is plausible that during RNA splicing a cryptic splice
Table 1. Detection of HBV-Human Chimeric Transcripts from Transcriptome Analysis
Cell Lines
Interrupted HBV ORF
Integrated Human Gene or
Sequence Location Description
HKCI-4 surface CHIC2intron 3 cysteine-rich hydrophobic domain 2 protein surface TERTupstream telomerase reverse transcriptase
X LINE1 on chr8p11 long-interspersed nuclear element 1 sequence core chr.5q14a-RALGPS1intron 3
(complex fusion)
Ral guanine nucleotide exchange factor with pleckstrin homology domain and avian sarcoma viral oncogene homology 3-binding motif 1 surface UPF3Aexon 10 UPF3 regulator of nonsense transcripts homolog A
HKCI-5B core ZNF48intron 2 zinc finger protein 48
core CDHR3exon 19-TRRAPintron 27 (complex fusion)
Cadherin-related family member 3; transformation/transcription domain-associated protein
X METAP1intron 1 methionyl aminopeptidase 1
HKCI-7 core LINE2 on chr11q13 long interspersed nuclear element 2 sequence polymerase TPCN2intron 21 two-pore segment channel 2
HKCI-11 X SINE on chr4q12 short interspersed nuclear element sequence
site was generated from HBs that led to the skipping of HBs (nt 459–2232) and the formation of an in-frame chimeric fusion of HBs-UPF3Aexon 10 (Figure 1B).
Expression of Silent Genomic Regions from Viral Integration
A possible oncogenic contribution of HBV DNA integration to cellular genes is the activation of transcriptional activities. To study thiscis-acting effect, we examined the expression levels based on FPKM values (fragments per kb of exon per million fragments mapped) of nearby protein-coding genes with viral
insertion. Although direct disruption of gene structure from viral DNA integration was found, an effect on cellular gene expression was not apparent. Cellular genes with or without viral insertion showed similar expression levels, suggesting that an effect on gene expression may not be evident (Figure S1B). We also sur-veyed the expression levels (in terms of FPKM) of genes located within 100 kbp upstream or downstream of all VFS-identified viral insertion sites. A direct correlation between expression level of a gene and its physical distance from the viral integration site was not observed (Figure S1C). The possible cis-regulatory effect of a viral insertion event on nearby genes would seem A
B
C
Figure 1. Illustration of Viral-Human Chimeric Transcripts and Sanger Sequencing Validation
(A) Viral insertion into genomic rearrangement ofCDHR3-TRRAPin HKCI-5B resulted in complex fusion transcription of HBc-CDHR3-TRRAP.
(B) Cryptic splice site from viral integration. VFS identified an in-frame fusion transcript of HBs-UPF3Aexon 10, whereas Sanger sequencing showed genomic insertion of HBs intoUPF3Aintron 9. An identical sequence motif, CAAGG (nt 455–459), could be found in the HBs gene and the splice junction ofUPF3Aexon 9. (C) Chimeric fusion of interrupted HBs ORFs and the human sequence upstream ofTERTwas detected by VFS in HKCI-4 and validated by Sanger sequencing. The figure highlights unique clipped-seq fragments of flanking viral and human regions with the microhomology sequence underlined. Theciseffect onTERTgene expression from proximal viral insertion was examined in HKCI-4 and other cell lines without nearby insertion. The arrow denotes the site of integration. See alsoFigure S1andTables S1andS2.
A B
C
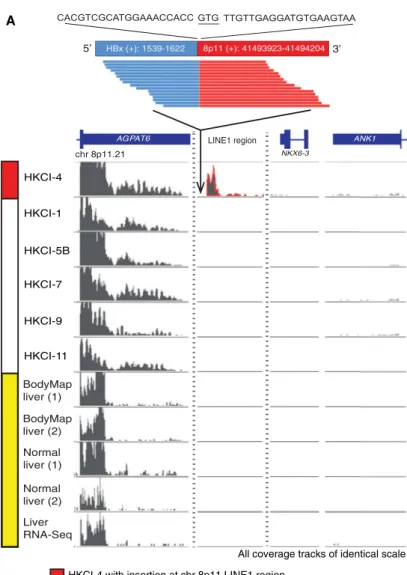
Figure 2. HBV Integration Can Activate Transcription of the Silent LINE1 Region
(A) Readthrough transcription of HBx-LINE1 from juxtaposed viral insertion into the chr.8p11.21 LINE1 region. RNA-seq data on liver tissue (Cabili et al., 2011), BodyMap 2.0 project on healthy liver samples (Sequence Read Archive accession number GSE30611), and transcriptome array on normal livers (Xu et al., 2011) signify the chr.8p11.21 LINE1 region as a transcriptionally silent heterochromatin. Cell lines without viral insertion in the region do not display any LINE1 expression. The partial HBx-LINE1 transcript sequence reconstructed using only the RNA-seq data is shown.
(B) Cell line HKCI-4 was established by our laboratory (Pang et al., 2002), and the corresponding patient’s HCC tumor (PHCC-4) was examined for the presence of the chimeric HBx-LINE1 transcript. RT-PCR analysis for HBx-LINE1 expression in HKCI-4 and the parental tumor PHCC-4 is shown. The cell line HKCI-9 and (legend continued on next page)
not to be apparent, although the involvement of trans-acting mechanisms cannot be ruled out.
Interestingly, we found active transcription of readthrough hu-man sequences from regions that are normally silent in the genome. For instance, an HBV insertion upstream ofTERTdid not elevateTERTexpression compared to other samples, but the transcribed sequence was derived from a region of nontran-scribing DNA (Figure 1C). Remarkably, a LINE1 sequence was cotranscribed from the HBV insertion into chr.8p11 in HKCI-4 (Figure 2A). This LINE1-rich region lies within the transcriptionally silent heterochromatin of chr.8p11.21 with no transcription factor binding sites as indicated by the ENCODE data set
(https://genome.ucsc.edu/ENCODE). Both RNA sequencing
(RNA-seq) data of multiple tissue types in the BodyMap 2.0 project (http://www.ebi.ac.uk/arrayexpress/experiments/
E-MTAB-513) (Sequence Read Archive accession number
GSE30611) and healthy liver samples (Cabili et al., 2011; Xu et al., 2011) also denote chr8, 41485000–41500000 nt, as a non-transcribing region in the human genome (Figure 2A). The dis-covery of a profound virus-driven LINE1 expression prompted our further investigations of the chimeric transcript HBx-LINE1. Expression of other silenced genomic regions from juxtaposition of viral insertion could also be found for HBc-LINE2 (average read depth 10.69 FPKM) and HBx-SINE (average read depth 11.11 FPKM) in HKCI-7 and HKCI-11, respectively, but their expression levels are lower than HBx-LINE1 (average read depth 60.35 FPKM).
Expression of HBx-LINE1 in HCC Tumors and Cloning of Full-Length Transcripts
We first confirmed HBx-LINE1 expression in HKCI-4 by RT-PCR and Sanger sequencing (Figure 2B). To rule out the possibility that HBV insertion was introduced as a result of prolonged in vitro culture, we examined the parental HCC tumor of HKCI-4 for the hybrid transcription. The primary tumors were accessible because the HKCI series of cell lines was established by our laboratory (Pang et al., 2002; Chan et al., 2006). Cell line HKCI-4 and its parental HCC tumor (PHCC-4) showed the consistent presence of the HBx-LINE1 hybrid transcript. In conjunction, we also examined HKCI-9, which served as a negative control. The expression of HBx-LINE1 was further demonstrated by in situ hybridization. Using a probe comple-mentary to the viral-human juncture, intense cytoplasmic signal of the HBx-LINE1 transcript was detected in HKCI-4 but not HKCI-9 (Figure 2B).
We proceeded to clone the full-length HBx-LINE1 transcript in HKCI-4. By 50and 30rapid amplification of cDNA ends (RACE) clonings, we isolated a 30 transcript of LINE1 that contained the polyadenylation signal AATAAA and a poly(A) tail and a 50 transcript entailing a C-terminal truncated HBx (Figure S2). The combined 30and 50end RACE enabled a full-length HBx-LINE1
transcript of 674 bp to be achieved (Figure 2C). The fusion tran-script entailed 297 bp from HBx that ranged from 1,326 to 1,622 nt of the HBV genome and 380 bp of LINE1 from chr.8p11 that spanned position 41493923–41494302 nt. There was a 3 bp microhomology at both breakpoint junctions, where the 50end of the HBx sequence shared a GTG sequence with the 30 end of the LINE1 sequence. We further examined the up-stream viral sequence of the genomic integration site. By primer walking, we determined the promoter of HBx 50upstream of the truncated HBx (Figure 2C), suggesting a possible viral promoter-driven transcription of the HBx-LINE1 fusion.
The incidence of the HBx-LINE1 chimeric transcript was examined in a cohort of 90 HBV-associated HCC tumors. Hemi-nested RT-PCR analysis indicated expression of HBx-LINE1 in 21 of 90 cases (23.3%) (Figure 3A). To corroborate the presence of this HBx-LINE1 fusion transcript, all PCR prod-ucts from primary tumors were sequence verified (Figure 3A; Fig-ures S3A and S3B). In all cases, the exact viral-human flanking sequence was found. We extended our study to investigate the association of HBx-LINE1 expression with prognosis of HCC patients. Kaplan-Meier analysis indicated that the presence of the HBx-LINE1 transcript could predict a shorter overall survival of patients (p = 0.0154;Figure 3B). Further univariate and multi-variate Cox regression analyses suggested that HBx-LINE1 expression remained a significant independent predictor of patient survival (p = 0.0165;Figure 3C) in addition to microvas-cular invasion. We also asserted the presence of HBx-LINE1 by an independent assay of real-time PCR using a custom probe that flanked the viral-human junction. In 20 HBV-associated tumors examined, HBx-LINE1 expression could be detected in 6 cases (30%) with estimated transcript levels ranging from 13,875 to 79,525 copies/mg RNA (Figures S3C–S3E).
We also examined the recurrence incidence of ten remaining viral-human chimeras discovered from the transcriptome (Table 1). Recurrence was not suggested for eight chimeric tran-scripts in 66 HBV-associated HCC tumors examined; only
HBs-UPE3Awas found in two cases (2/66; 3.03%) and HBx-METAP1
in one case (1/66; 1.52%) (Figure S3F).
HBx-LINE1 Promotes Cell Motility through Epithelial-to-Mesenchymal Transition
We postulated that the high prevalence of HBx-LINE1 expres-sion in HCC may hold biological significance. To elucidate the potential functional contributions from this chimeric transcript, knockdown experiments with two small interfering RNAs (siRNAs), siHBx-LINE1a and siHBx-LINE1b, were performed in HKCI-4. The knockdown efficiencies were confirmed by quanti-tative (q)RT-PCR (Figure 4A). Whereas siRNA-mediated target-ing of HBx-LINE1 did not seem to affect cell viability, more apparent effects were indicated from Transwell migration and Matrigel invasion (Figures 4B and 4C). Both siHBx-LINE1a and
parental tumor PHCC-9 pair served as negative control. In situ hybridization for HBx-LINE1 transcript in HKCI-4 and HKCI-9 is also presented. The scale bars represent 50mm.
(C) Schematic diagram of the HBx-LINE1 flanking sequence. RACE cloned a full-length transcript of 674 bp with an ATG start codon indicated in HBx and a TGA stop codon on chr.8p11. The viral DNA sequence upstream of the HBx ORF was examined by primer walking. The sequence of the 674 bp full-length HBx-LINE1 transcript is shown with numbers denoting nucleotide position. Blue highlights the viral sequence and black highlights the human sequence, and a 3 bp GTG microhomology at both breakpoint junctions is underlined.
siHBx-LINE1b could significantly reduce migratory and invasive properties of HKCI-4 cells compared to siMock control. Because the phenomenon of epithelial-to-mesenchymal transition (EMT) is a major molecular mechanism by which cancer cells acquire motile ability to invade the tumor microenvironment, we further investigated the effect of HBx-LINE1 on the modulation of EMT. Western analysis of siHBx-LINE1-treated HKCI-4 revealed increased expression of epithelial markers E-cadherin and g-catenin and diminution of the mesenchymal protein fibronectin (Figures 4D and 4E). These findings were further substantiated in immunofluorescence analysis, which corroborated the augmen-tation of E-cadherin andg-catenin expression and their promi-nent accumulation at the cell membrane in siHBx-LINE1-treated cells. In comparison, a more diffuse cytoplasmic localization of these markers was suggested in both siMock and Lipofect-amine-treated control cells. A decrease in the level of cyto-plasmic fibronectin was also observed in siHBx-LINE1-treated cells relative to control treatments (Figure 4F).
HBx-LINE1 Activates Wnt/b-Catenin Signaling
It is known thatb-catenin forms a membranous complex with the cell-adhesion protein E-cadherin and other catenin proteins. During the process of EMT,b-catenin is dissociated from the membrane-associated E-cadherin complex and then translo-cates to the nucleus, where it activates the Wnt signaling pathway (Polette et al., 2007). To examine whether HBx-LINE1 has an effect on Wnt/b-catenin signaling, we next examined the nuclear localization and transactivity ofb-catenin under the influence of HBx-LINE1. In HKCI-4 cells treated with siHBx-LINE1a and siHBx-siHBx-LINE1a, an effect on the total cellularb -cate-nin protein was not suggested but both siRNAs concurred to suggest a reduced level of active b-catenin by western blot
(Figure 4G). By TOP/FOPflash assay, we found a reduced
transactivation activity ofb-catenin following HBx-LINE1 knock-down, which is in line with the lowered activeb-catenin protein found (Figure 4H). Cellular distribution ofb-catenin as revealed from immunofluorescence staining also indicated diminution of A HBx-LINE1 GAPDH HBx-LINE1 GAPDH HBx-LINE1 GAPDH C HCC#14 HCC#26 HCC#31 HCC#39 *p=0.0154 HBx-LINE1 positive (n=21) HBx-LINE1 negative (n=69)
Follow-up Time (months)
Percent Survival B e t a i r a v i t l u M e t a i r a v i n U e u l a v p I C % 5 9 o i t a r d r a z a H e u l a v p I C % 5 9 o i t a r d r a z a H e l b a i r a V HBx-LINE1 Positive Negative 2.3529 1.2054-4.5928 0.0122 2.6491 1.1942-5.8763 0.0165 3 6 2 1 . 0 0 6 6 5 . 1 -5 6 2 0 . 0 8 3 0 2 . 0 5 1 7 0 . 0 7 3 7 1 . 1 -0 2 2 0 . 0 8 0 6 1 . 0 r e d n e G Age+ 0.6845 0.2854-1.6417 0.3957 1.1728 0.4147-3.3163 0.7638 Clinical stage* 2.0255 1.3642-3.0076 0.0005 1.0302 0.4474-2.3724 0.9443 Tumor grade# 1.6111 0.7837-3.3119 0.1946 1.1671 0.5306-2.5674 0.7008 Macro-vascular invasion^ 2.1211 0.6449-6.9758 0.2158 0.4701 0.0986-2.2409 0.3435 Micro-vascular invasion^ 3.3452 1.5924-7.0274 0.0014 4.3280 1.4535-12.8876 0.0085 8 7 8 6 . 0 0 1 0 2 . 3 -2 4 6 4 . 0 9 8 1 2 . 1 3 8 8 1 . 0 8 2 2 3 . 4 -8 9 4 7 . 0 4 0 0 8 . 1 ^ s i s o h r r i C Tumoral lesionsI 3.3135 1.7255-6.3627 0.0003 2.6368 0.9491-7.3256 0.0629 HBx LINE1 G GCATG GAAAC CAC C TGTTGTTGAG GATGT HBx LINE1 G GCATG GAAAC CAC C TGTTGTTGAG GATGT HBx LINE1 G GCATG GAAAC CAC C TGTTGTTGAG GATGT HBx LINE1 G GCATG GAAAC CAC C TGTTGTTGAG GATGT
Figure 3. Expression of HBx-LINE1 in Primary HCC
(A) Representative images of RT-PCR analysis of the HBx-LINE1 transcript in primary HCC tumors. NTC, no template control. All PCR bands were also subjected to sequence verification by Sanger sequencing.
(B) Kaplan-Meier curves depicting HCC patients with positive (dotted line) and negative (solid line) expression for HBx-LINE1. (C) Univariate and multivariate Cox regression analysis for poor prognostic factors affecting overall survival of HCC patients.+
Age: age %40 versus age >40. *Clinical stage: T1 versus T2 versus T3. Tumor stage is classified according to American Joint Committee on Cancer tumor-node-metastasis staging criteria (Greene et al., 2002).#
Tumor grade: poorly differentiated versus moderately differentiated versus well-differentiated. ^Macrovascular invasion, ^microvascular invasion, and ^cirrhosis: presence versus absence.I
Tumoral lesions: solitary versus multiple. See alsoFigure S3andTable S3.
Fibronectin E-cadherin γ-catenin GAPDH 0 0.1 0.2 0.3 0.4 0.5 0.6 1 2 3 4 5 6 7 Absorbance Days Lipo siMock siHBx-LINE1a siHBx-LINE1b C E 0.0 0.5 1.0 1.5 2.0 Normalized fold change
*
*
E-cadherin 0.0 0.5 1.0 1.5 2.0 Normalized fold change**
**
γ-catenin 0.0 0.3 0.6 0.9 1.2 Normalized fold change*
*
Fibronectin 0 0.2 0.4 0.6 0.8 1 1.2 Relativ e e x p ression of HBx -LINE1*
*
A 0% 40% 80% 120% Relativ e p ercentage of inv a ded c ells*
*
0% 40% 80% 120% Relativ e p ercentage of migrated cells*
*
B 0.0 0.4 0.8 1.2 TOP/FOPflash activ ity*
**
H F siHBx -LINE1a siMock LipoE-cadherin γ-catenin Fibronectin
s iHBx -LINE1b G total β-catenin GAPDH active β-catenin D I Lipo
β-catenin DAPI Merged
siHBx
-LINE1a
siMock
siHBx
-LINE1b
Figure 4. Knockdown of HBx-LINE1 Inhibits Cell Motility through Regression of EMT (A) Efficacy of siHBx-LINE1a and siHBx-LINE1b confirmed by qPCR.
(B) Relative cell migration and invasion in cells treated with siHBx-LINE1a and siHBx-LINE1b compared to Lipo and siMock controls. (C) Effect on cell viability by MTT assay.
(D) Western blot analysis of E-cadherin andg-catenin (epithelial markers) and fibronectin (mesenchymal marker) in siHBx-LINE1a and siHBx-LINE1b knockdown cells compared to siMock and Lipo controls.
(E) Densitometry measurements of western blots from three independent experiments.
(F) Immunofluorescence staining for the EMT process. Expression of adhesion proteins E-cadherin andg-catenin and cytoplasmic fibronectin in siHBx-LINE1a-and siHBx-LINE1b-treated cells is illustrated. Nuclei were counterstained with 40,6-diamidino-2-phenylindole (DAPI).
(G) Western blot for active and totalb-catenin levels. (H) TOP/FOPflash reporter assay forb-catenin transactivity.
(I) Immunofluorescence staining forb-catenin in HBx-LINE1 knockdown cells compared to siMock and Lipo controls. The scale bars represent 10mm. All data shown are mean ± SEM from three independent experiments (**p < 0.01, *p < 0.05).
b-catenin nuclear localization from the effect of siHBx-LINE1a and siHBx-LINE1b, whereas Lipofectamine-treated and siMock control cells showed more intense accumulation ofb-catenin within the nucleus (Figure 4I).
HBx-LINE1 Exerts Functional Effects as a Long Noncoding RNA-like Transcript
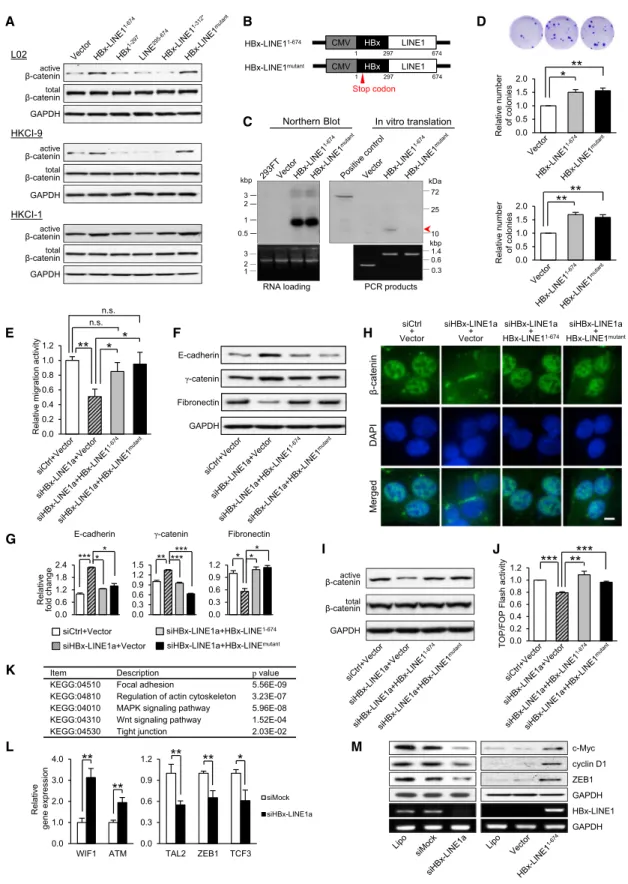
To determine whether the viral component, human sequence, or full-length HBx-LINE1 underlies the functional effects ob-served, we next cloned and expressed the full-length HBx-LINE11–674 and variants including HBx-LINE11–312*, HBx1–297, and LINE1295–674for translated proteins (Figure 5A). GFP-fused protein expressed from these clones showed protein sizes in accordance with prediction, where minimal differences could be detected between proteins translated from HBx-LINE11–674 and HBx-LINE11–312* (Figure 5B). Translated proteins from full-length HBx-LINE11–674showed a 48.4 kDa protein that is analo-gous to the HBx-LINE11–312* variant. Both constructs translated 82 amino acids from the truncated HBx region and an extra 5 amino acids from the LINE1 human region, which harbored a stop codon at position 310–312 nt (Figure 5A). Immunofluores-cence analysis showed a diffuse cytoplasmic distribution in cells transfected with HBx-LINE11–674that was similar to the other three constructs and GFP-vector alone (Figure S4A). Transcribed RNA of the cloned inserts HBx-LINE11–674, HBx-LINE11–312*, HBx1–297, and LINE1295–674 from GFP-deleted C2R(-) constructs was also assessed in transfected L02, and the molecular weights determined were in concordance with predicted size (Figure 5B).
In vitro functional investigations in immortalized human hepa-tocyte cell line L02 showing only cells expressing full-length HBx-LINE11–674 displayed noticeable colony growth, whereas expression of HBx-LINE11–312*, HBx1–297, and LINE1295–674 showed similar effects as the vector control (Figure 5C). The pos-sibility of functional augmentation being attributed to a viral-human fusion protein was eliminated by the fact that protein translated from HBx-LINE11–674 was analogous to HBx-LINE11–312*, and no growth advantage was conferred by HBx-LINE11–312*. In addition, transfection of full-length HBx-LINE11–674and truncated mutants in L02 suggested only HBx-LINE11–674could promote cell migration and invasion (Figure 5C). We further explored the changes in EMT markers, but it was noted that L02 failed to show detectable E-cadherin expression in repeated experiments. We hence replaced E-cadherin with
another epithelial marker, CK18, and included a hallmark marker, N-cadherin, for the mesenchymal change. Remarkably, marked expression and typical localization of N-cadherin to the cell membrane was evident in HBx-LINE11–674-transfected L02, signifying a mesenchymal change (Figures 5D and 5E). A corre-sponding increase in the expression of fibronectin was also found in HBx-LINE11–674-transfected cells. Expression of CK18 andg-catenin, on the other hand, was reduced following HBx-LINE11–674transfection, but not with other variant constructs and the vector control (Figures 5D and 5E). We also examined the functional effects of these expressing vectors in HKCI-9 and HKCI-1 and found a similar enhancement in cell growth and motility from the ectopic expression of HBx-LINE11–674( Fig-ure 5F;Figures S4B and S4C). This stimulation corresponded to an increased TOP/FOP transactivity, which was persistently higher in all three cell lines transfected with HBx-LINE11–674 but not HBx-LINE11–312* (Figure 5G). Image analysis showed nuclear localization of b-catenin with HBx-LINE11–674but not other variants (Figure 5H).
To affirm involvement of the EMT process in HKCI-9 and HKCI-1 as well from the transfection of HBx-LINE11–674, western analysis showed an increased expression of fibronectin and downregulation of E-cadherin and g-catenin (Figures 5I and 5J). The change in level of these protein markers also corre-sponded to immunofluorescence imaging, where marked reduction in the membranous localization of epithelial adhesion molecules was found and prominent fibronectin cytoplasmic localization was shown (Figure 5K;Figure S4D). More notably, western analysis also revealed the level of activatedb-catenin to be consistently elevated in all three cell lines when subjected to full-length HBx-LINE11–674induction, but this phenomenon was not found with other truncated variants (Figure 6A). Our data would imply that protein translated from HBx-LINE11–674 cannot account for the distinct functional phenotypes observed from ectopic expression. It is hence conceivable that HBx-LINE11–674 exerts its functional effects as a long noncoding RNA (lncRNA)-like transcript.
To assert that HBx-LINE11–674functions as a hybrid RNA, we further constructed a mutant with a stop codon introduced at the beginning of HBx, so that the protein translation was inhibited but RNA production was maintained (Figures 6B and 6C). Both HBx-LINE1mutantand HBx-LINE11–674were examined for func-tionality in colony-forming growth and migratory ability. We found that the HBx-LINE1 stop mutant expressed in L02 and
Figure 5. Full-Length HBx-LINE1-Mediatedb-Catenin Activation Promotes Cell Growth and Motility through EMT Progression (A) Schematic diagram of constructs of full-length HBx-LINE11–674
, HBx1–297
, LINE1295–674
, and HBx-LINE11–312
*.:andDdenote stop codons on LINE1 and vector, respectively. Each plasmid was cloned into two expression vectors, pEC2 and pEC2R(-). Plasmids cloned into a pEC2 vector, with a GFP-coding sequence, were examined for tag-fused protein translation and localization. The series of plasmids cloned into the pEGFP-C2R(-) vector, with a deleted GFP sequence, was used in all functional assays described. CMV, cytomegalovirus.
(B) GFP-conjugated protein translation and RT-PCR analysis in L02.
(C) Effect of full-length and truncated variants on cell growth, migration, and invasion analysis in L02. (D and E) Effect of HBx-LINE11–674
and truncated variants on the EMT process in L02. Results from western blot and immunofluorescence imaging are shown. (F) Effect of full-length and truncated variants on cell growth, migration, and invasion analysis in HKCl-9.
(G) TOP/FOPflash assay forb-catenin transactivity in L02, HKCI-9, and HKCI-1. (H) Immunofluorescence analysis forb-catenin localization in transfected L02 cells.
(I and J) Western analysis and densitometry measurements for changes in EMT markers in HKCI-9 and HKCI-1. (K) Immunofluorescence results in HKCI-9.
The scale bars represent 10mm. All data represent mean ± SEM values from three independent experiments (***p < 0.001, **p < 0.01, *p < 0.05; n.s., no sig-nificance). See alsoFigure S4.
E-cadherin γ-catenin Fibronectin GAPDH K L β-catenin DAPI Merged siHBx-LINE1a + HBx-LINE11-674 siHBx-LINE1a + Vector siCtrl + Vector siHBx-LINE1a + HBx-LINE1mutant 0.0 0.2 0.4 0.6 0.8 1.0 1.2 TOP/FOP Flash a ctiv ity
***
**
***
G J I active β-catenin total β-catenin GAPDH H F E M 0.0 0.2 0.4 0.6 0.8 1.0 1.2 Relativ e migration activ ity**
*
*
n.s. n.s. 0.0 0.3 0.6 0.9 1.2 0.0 0.6 1.2 1.8 2.4E-cadherin γ-catenin Fibronectin
0.0 0.3 0.6 0.9 1.2 1.5 siCtrl+Vector siHBx-LINE1a+HBx-LINEmutant siHBx-LINE1a+HBx-LINE1-674 siHBx-LINE1a+Vector ***** ** ****** * ** Relativ e fold change 0.0 1.0 2.0 3.0 4.0 WIF1 ATM 0.0 0.3 0.6 0.9 1.2 TAL2 ZEB1 TCF3 siMock siHBx-LINE1a Relativ e gene ex pression
**
**
**
**
*
HBx-LINE1 GAPDH cyclin D1 c-Myc GAPDH ZEB1Item Description pvalue
KEGG:04510 Focal adhesion 5.56E-09
KEGG:04810 Regulation of actin cytoskeleton 3.23E-07 KEGG:04010 MAPK signaling pathway 5.96E-08 KEGG:04310 Wnt signaling pathway 1.52E-04 2 0 -E 3 0 . 2 n o i t c n u j t h g i T 0 3 5 4 0 : G G E K A active β-catenin total β-catenin GAPDH total β-catenin active β-catenin GAPDH total β-catenin active β-catenin GAPDH L02 HKCI-9 HKCI-1 D B Relativ e number of coloni e s 0.0 0.5 1.0 1.5 2.0
**
**
0.0 0.5 1.0 1.5 2.0 Relativ e number of coloni es**
*
C PCR products RNA loadingNorthern Blot In vitro translation
0.3 0.6 1.4 10 25 72 kDa kbp 3 2 1 kbp 3 2 1 0.5 CMV HBx-LINE11-674 1 CMV 1 HBx-LINE1mutant 297 674 297 674 Stop codon LINE1 HBx LINE1 HBx
Figure 6. HBx-LINE1 Exerts Functional Effects as a Long Noncoding RNA-like Transcript (A) Western analysis of activeb-catenin in the presence of HBx-LINE11–674
, HBx-LINE1mutant
, and truncated variants in L02, HKCI-9, and HKCI-1. (B) Schematic diagram of HBx-LINE11–674
and HBx-LINE1mutant
; the latter harbors a stop codon (red arrowhead) on HBx.
(C) Northern blot shows HBx-LINE11–674and HBx-LINE1mutanttranscripts at the expected length of900 bp (674 bp from HBx-LINE1 to the vector frame poly[A] tail of220 bp). In vitro translation showed a protein band from HBx-LINE11–674
(red arrowhead) at the predicted size of 11.4 kDa. No evident band could be detected from HBx-LINE1mutant
.
HKCI-9 showed similar colony-formation ability as HBx-LINE11–674, which emphasized the activity being derived from the RNA transcript independent of its coding capacity ( Fig-ure 6D). To affirm the function of HBx-LINE1mutant in cancer cell motility, we further conducted rescue experiments in HKCI-4 by reintroducing HBx-LINE1mutantor HBx-LINE11–674in the presence of si knockdown of HBx-LINE1. We found that ectopic reexpression of both HBx-LINE1mutant and HBx-LINE11–674could readily restore the cell motile phenotype from the inhibitory effect of si-HBx-LINE1 (Figure 6E), which further enforced the oncogenicity of HBx-LINE1 as a hybrid RNA. The expression of epithelial and mesenchymal markers in siHBx-LINE1+HBx-LINE1mutant cells also confirms the lncRNA-like function of HBx-LINE1. Western blot showed that siHBx-LINE1-induced expression of E-cadherin and g-catenin was downregulated in the presence of HBx-LINE1mutant. Inversely, mesenchymal fibronectin regained upregulation in the rescue of siHBx-LINE1+HBx-LINE1mutant(Figures 6F and 6G). Immuno-fluorescence imaging showed that both full-length and stop mutant HBx-LINE1 could revert nuclear localization ofb-catenin that corresponded to a rescue of activeb-catenin levels (Figures 6H and 6I). In agreement with these findings, the TOP/FOPflash assay also suggested an increase inb-catenin activity in siHBx-LINE1+HBx-LINE1 and siHBx-siHBx-LINE1+HBx-LINE1mutant com-pared to siHBx-LINE1-treated cells (Figure 6J).
To rule out the possible involvement of microRNA in the func-tional phenotypes observed, we conducted in vitro Drosha/Dicer experiments for microRNA production from endogenous cleav-age of HBx-LINE11–674. From northern blotting, we did not find enzymatic cleavage of full-length HBx-LINE1 into small 18– 22 nt fragments, which suggested the lack of hairpin folding of HBx-LINE11–674for microRNA maturation (Figure S5). Our re-sults would hence support a functional role for HBx-LINE1 as an lncRNA-like transcript in modulating HCC cell motility and growth.
To gain insight into the action of HBx-LINE1, we performed expression profiling of siHBx-LINE1- and siMock-treated cells. KEGG analysis (http://www.genome.jp/kegg) of differentially ex-pressed genes indicated pathways under the modulation of HBx-LINE1 (Figure 6K). RT-PCR verified an effect of siHBx-LINE1 on the upregulation of Wnt inhibitorsWIF1andATMand downregulation ofZEB1,TAL2, andTCF3(Figure 6L). To further substantiate an effect on Wnt signaling, we examined expression of c-Myc and cyclin D1, two well-known direct targets ofb -cat-enin, andZEB1, which is a master regulator of EMT. Western blot showed reduced levels of c-Myc, cyclin D1, and ZEB1 following HBx-LINE1 knockdown in HKCI-4, whereas upregulation of these proteins was readily detected when HBx-LINE1 was over-expressed in L02 (Figure 6M).
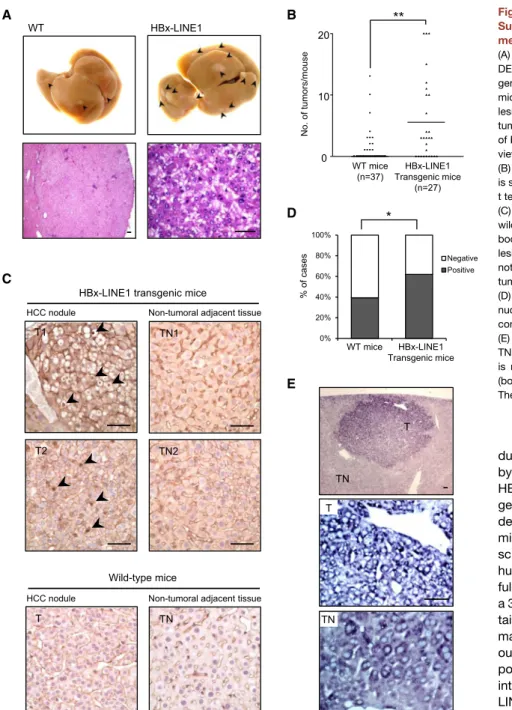
HBx-LINE1 Increases Risk of HCC Development
Upon diethylnitrosamine (DEN)-induced HCC formation, gross liver morphology of HBx-LINE1 and wild-type mice showed tumor development at 8 months.Figure 7A shows gross features and histology of the murine HCC tumors formed. HBx-LINE1 transgenic mice from three strains were found to be consistently more susceptible to DEN-induced tumor formation, where a significantly higher number of HCC tumors was found in mice with the transgene (mean 5.59, p = 0.0018) compared to wild-type (mean 1.38) (Figure 7B). Immunohistochemical staining further showedb-catenin nuclear localization in HCC tissue of transgenic mice (Figure 7C). A significant difference in the nu-clear positivity of b-catenin was further determined between transgenic (n = 31/50 tumor nodules; 62%) and wild-type ani-mals (n = 24/61 tumor nodules; 39%) (p=0.018), which would be suggestive of a more direct effect for HBx-LINE1 in modu-lating Wnt/b-catenin signaling (Figure 7D). In situ hybridization of tissues from HBx-LINE1 transgenic mice showed signals mainly localized within the tumor cytoplasm (Figure 7E). DISCUSSION
HBV is a hepadna virus that almost invariably integrates into the HCC genome. Whereas integrations of HBV were once thought to be random, recent genome resequencing of HCC has begun to highlight recurrent integrated regions and greatly expanded the list of targeted genes. In this study, using combined informa-tion on read-pair mapping and breakpoint-spanning reads from whole-transcriptome sequencing, we reconstructed viral and human flanking sites and junctures, which could be indepen-dently confirmed at both genomic DNA and transcribed RNA levels (Figure S1A). In line with published results on whole-genome sequencing of HBV-associated HCC, integration of HBV upstream ofTERT, withinRALGPS1, and in the intergenic region of chr.11q13 could also be found from our analysis (Jiang et al., 2012; Sung et al., 2012; Toh et al., 2013). More interest-ingly, we found active transcription of HBx-LINE1 from the viral insertion into a normally silent intergenic region, chr.8p11.21. In fact, the same integrated region chr.8p11.21 has also been re-ported from a whole-genome sequencing study bySung et al. (2012)and a captured sequencing study of enriched HBV inte-grants by Toh et al. (2013). In addition, previous PCR-based cloning studies, despite their inherent low resolution, coincided in suggesting regional chr.8p11 as a favorable viral integration site (Murakami et al., 2005). Nevertheless, information on the transcriptional effects resulting from viral insertion into this re-gion remains minimally studied. Here we show de novo tran-scription of LINE1 from chr.8p11.21. In addition to its mutagenic role as mobile genetic elements (or retrotransposons), we report (D) Effect of HBx-LINE11–674
and HBx-LINE1mutant
expression on the colony-forming ability of L02 and HKCI-9 cells. (E) Rescue of the migratory phenotype from ectopic transfection of HBx-LINE11–674
and HBx-LINE1mutant
in HKCI-4. (F and G) Western analysis for E-cadherin,g-catenin, and fibronectin, and densitometry measurements.
(H) Immunofluorescence staining forb-catenin localization. The scale bar represents 10mm. (I and J) Western blot and TOP/FOPflash assay for activeb-catenin.
(K) Analysis of gene expression profiling for affected pathways by HBx-LINE1. (L) Selected candidates were verified by qPCR.
(M) Western analysis ofb-catenin targets c-Myc, cyclin D1, and ZEB1 in knockdown and overexpressed HBx-LINE1.
All data shown represent mean ± SEM values from three independent experiments (***p < 0.001, **p < 0.01, *p < 0.05; n.s., no significance). See alsoFigure S5and Table S4.
here on oncogenic effects of LINE1 in the chimera with HBx. Based on our results, we believe that viral-induced transcription of fusion hybrids may potentially unveil an additional layer of transcript expression in HBV-associated pathogenesis.
The fact that HBV integrations have been mapped mostly to noncoding repetitive regions has dampened research interest in further investigation for many years. This was largely attributed to postulations that suggested viral DNA insertions merely allowed persistence of HBV in the host (Feitelson and Lee, 2007) or simply induced a more generalized phenomenon of fragile chromosomes (Kremsdorf et al., 2006). The significance of integrants in intronic or intergenic regions would seem to have been underestimated. In this study, our analysis suggests that HBx-LINE1 transcription can provide a selective advantage A WT HBx-LINE1
**
No. of tumors/mouse B WT mice (n=37) HBx-LINE1 Transgenic mice (n=27) 0 10 20 D 0% 20% 40% 60% 80% 100% Negative Positive WT mice HBx-LINE1 Transgenic mice % of cases*
Non-tumoral adjacent tissue HCC nodule
C
TN1
TN2 HBx-LINE1 transgenic mice
Non-tumoral adjacent tissue HCC nodule Wild-type mice T TN E T TN T TN T1 T2
Figure 7. HBx-LINE1 Transgene Increases Susceptibility to DEN-Induced HCC Develop-ment
(A) Typical gross morphology of liver tumors from DEN treatment at 8 months in HBx-LINE1 trans-genic (upper right) or wild-type (upper left) male mice. Arrowheads indicate macroscopic HCC lesions. Lower: histology of a representative HCC tumor nodule. Lower left: intermediate-power view of HCC tumor (H&E stain). Lower right: high-power view of HCC histology (H&E stain).
(B) The average number of HCC tumors per mouse is shown with the mean value highlighted (unpaired t test, **p = 0.0018).
(C) Immunohistochemical staining of transgenic and wild-type liver sections using anti-b-catenin anti-body. Clear nuclear staining ofb-catenin in tumor lesions was found in transgenic HCC tumors (de-noted by arrowheads) but not in adjacent non-tumoral liver (TN1 and TN2) and wild-type liver. (D) Frequency of tumor nodules harboring positive nuclear staining forb-catenin in transgenic mice compared to wild-type mice (c2
, *p = 0.018). (E) In situ hybridization for HBx-LINE1 in T and TN (top). Subcellular localization of HBx-LINE1 is mainly in the cytoplasm of T (middle) and TN (bottom).
The scale bars represent 50mm.
during tumorigenesis, rather than being a byproduct event from random insertion. HBx-LINE1 transcription occurred through genomic insertion of truncated HBx in tan-dem with the 50end HBx promoter, which might underscore the viral activated tran-scription of an otherwise nontranscribing human LINE1 repetitive sequence. The full-length HBx-LINE1 transcript contained a 30end polyadenylation signal and poly(A) tail, which would signify the process of mature RNA production. Whereas previ-ous studies of HBV integrants have re-ported a low frequency of recurring cellular integrations, we detected recurrent HBx-LINE1 expression in 23.3% of HBV-associ-ated HCC, suggesting a preferential site of insertion. Cox regression analysis further suggested HBx-LINE1 as an independent prognostic factor for predicting shorter survival of HCC patients. We also demon-strated from extensive in vitro functional investigations that HBx-LINE1 holds an important role in promoting tumor growth and as a mediator of cell motility through the EMT process. This behavioral change is likely through activation of Wnt/b -cat-enin signaling, of which we showed expression of c-Myc, cyclin D1, and ZEB1 can be spontaneously modulated by HBx-LINE1. In inducing an EMT,ZEB1is known not only to transcriptionally repress epithelial markers such as E-cadherin but also to acti-vate mesenchymal genes (Sa´nchez-Tillo´ et al., 2011). Of interest, nuclear translocated b-catenin was reported to induce ZEB1 expression by direct binding to the ZEB1 promoter region (Sa´nchez-Tillo´ et al., 2011). An effect of HBx-LINE1 on ZEB1
throughb-catenin activation is hence likely. In fact, aberrant acti-vation of Wnt/b-catenin signaling is a major pathway in the onco-genesis of HBV-associated HCC (Whittaker et al., 2010). Nuclear and/or cellularb-catenin accumulation, a hallmark of activated canonical Wnt signaling, has been observed in65% of HCC tumors (Devereux et al., 2001; Wong et al., 2001). However, mutations of the b-catenin gene are found in only 13% of HBV-associated HCCs (Huang et al., 2012; Wong et al., 2001), suggesting that regulatory mechanisms other than b-catenin mutations are likely involved in activating Wnt signaling. We hy-pothesize here that HBx-LINE1, because of its frequent expres-sion in HCC, may represent another upstream component in the activation of the Wnt/b-catenin cascade. Studies have also re-ported that HBV integration can be found during the course of chronic infection and hence be considered an early carcinoge-netic event (Bre´chot et al., 2000). In line with this notion, transfec-tion of HBx-LINE1 into the immortalized hepatocyte cell line L02 promoted colony growth. Moreover, functional involvement of HBx-LINE1 in promoting cancer susceptibility was substantiated in a transgenic model, where DEN-induced HCC tumor forma-tions were significantly higher in mice with the HBx-LINE1 trans-gene. Taken together, our data highlight the importance of the HBx-LINE1 transcript in conferring oncogenic advantages that may represent an elemental predisposing factor in the initiation and progression of HCC.
In addition to the aforementioned findings, this study further demonstrates that proteins translated from HBx-LINE1 cannot account for the functional phenotypes observed. Nevertheless, HBx-LINE1 translates an 11.4 kDa C-terminal truncated HBx. Many studies have shown that C-terminal truncated HBx protein holds oncogenic properties in HCC pathogenesis, although the degree of truncation varies with function. Given that the major functional breakpoints in primary HCCs are between 125 and 135 amino acids (Sze et al., 2013), the truncated HBx translated from our current HBx-LINE1 chimera, at a breakpoint of 99 amino acids, may well have minimized its tumor-promoting func-tion with an overall smaller HBx protein translated. Interestingly, results obtained with the HBx-LINE1 stop mutant showed similar functional phenotypes as HBx-LINE11–674, which accentuate the pathogenic role of HBx-LINE1 as an lncRNA-like transcript. Recent transcriptome studies have begun to highlight abundant expression of lncRNA in the mammalian genome. There is also increasing evidence highlighting vital roles for lncRNA in regu-lating cellular processes and their expression in contributing to disease phenotypes such as cancer (Braconi et al., 2011; Ørom et al., 2010). To date, only a few lncRNAs, namelyHOTAIR, HULC,H19, andTUC338, have been studied in HCC (Braconi et al., 2011; Matouk et al., 2007; Panzitt et al., 2007; Yang et al., 2011). The regulatory roles of most lncRNAs in liver carci-nogenesis are therefore still largely unexplored. Adding to this repertoire, our present study isolated a chimeric transcript that holds functional advantages. Because protein encoded by HBx-LINE1 cannot account for the tumorigenic effects, the oncogenicity instead is directly dependent on the hybrid RNA; this underlines the activity of HBx-LINE1 as an lncRNA-like tran-script. We hence report here that HBV insertion into a host genome can incite chimeric transcription, and possibly high-lights another level of intricacy in the production of functional hybrid RNAs in cancer.
It is known that human LINE-1 retrotransposons are a major source of endogenous mutagens (Burns and Boeke, 2012; Levin and Moran, 2011). In HCC, LINE1 has recently been shown to be a crucial source of mutations that can reduce the tumor-suppres-sive capacity of somatic cells (Shukla et al., 2013). Here we show oncogenic activity of LINE1 as a chimera with HBV, which poten-tially may signify a functionally distinct class of lncRNA-like transcripts. Our current study also provides evidence of the molecular complexity of HBV integrations, where virus inserted into the genome could create a cryptic splice site or be tran-scribed as a complex fusion when genomic rearrangements are involved. Many viral-human chimeras identified also showed a microhomology sequence at both breakpoint junctions. It is plausible that the mechanism of HBV insertion into the host genome was likely generated via the mediation of a microhomol-ogy during break-induced replication repair (Hastings et al., 2009). Moreover, the chimeric transcripts determined showed an identical sequence to the flanking viral-human DNA fragment, which suggests a direct readthrough transcription from the site of genomic integration. In summary, our results show a common transcription of a viral-human chimera in response to HBV genome integration. Our study, together with recent results from genome sequencing, provides a basis for investigative ana-lyses that may lead to further understanding of the effect by which HBV insertion supports cancer development.
EXPERIMENTAL PROCEDURES Transcriptome Sequencing
Six HCC cell lines, namely HKCI-1, HKCI-4, HKCI-5B, HKCI-7, HKCI-9, and HKCI-11, were subjected to whole-transcriptome sequencing. These cell lines were developed from HBV-infected HCC patients who underwent curative surgery at Prince of Wales Hospital, Hong Kong. Because these cell lines were established within our institute (Chan et al., 2006; Pang et al., 2002), the early culture passages were accessible and had been used for the sequencing described here. Briefly, TRIzol-extracted RNA was subjected to sequencing library preparation using an Illumina TruSeq RNA Sample Prepa-ration Kit according to the manufacturer’s instructions. RNA-seq was per-formed on an Illumina HiSeq 2000 platform, generating paired-end sequence reads of read length 101 bp with an average insertion size of 300 bp. On average, 127 M reads were obtained per sample with mappable reads at an average of 92.87%. Detailed sequencing statistics are shown inTable S2. Detection of Viral-Human Fusion Transcripts by ViralFusionSeq The analysis tool for discovery of viral-human fusion transcript in this study, ViralFusionSeq, was recently published by our group (Li et al., 2013). VFS pinpoints the locations of viral-human fusion events by integrating two comple-mentary approaches into the analysis: breakpoint-spanning reads and discor-dant read pairs. For paired-end sequencing, a majority of reads in a pair would be mapped, within the expected insertion size, together onto either the human or viral genome. A possible viral-human fusion is implied by (1) mapping of two reads in a pair separately onto the viral and human genomes; and/or (2) partial mapping of a sequence read within a given pair onto both the viral and human genomes. VFS exploits these two events: the read-pair (RP) analysis module to tackle the former, and utilizes the postalignment sequences in the clipped-seq (CS) analysis module to interrogate the latter case. Finally, a targeted de novo assembly using both the RP and CS reads was performed for each fusion candi-date to reconstruct the fusion transcript. Details of the VFS algorithm and analytical methods are described inSupplemental Experimental Procedures. Detection of the HBx-LINE1 Transcript in HCC Tumors
Tumorous liver tissues were collected from 90 patients who underwent cura-tive surgery for HCC at Prince of Wales Hospital, Hong Kong. Informed con-sent was obtained from each recruited patient and the study protocol was
approved by the Clinical Research Ethics Committee of the Chinese University of Hong Kong. Demographics of the patient cohort are shown inTable S3. The diagnosis of HCC was confirmed by histology examination. The patients re-cruited were all chronic carriers of HBV. The HBx-LINE1 transcript in HCC tumors was analyzed by RT-PCR. Reverse-transcribed cDNA was amplified by a hemi-nested PCR using specific primers designed for the juxtaposed positions of the viral-human flanking junction. Details are described in Supple-mental ExperiSupple-mental Procedures.
RNAi Knockdown of HBx-LINE1
Two oligonucleotides targeting the HBx-LINE1 transcript were designed: siHBx-LINE1a (289–313 bp, 50-ACCACCGUGUUGUUGAGGAUGUGAA-30) and siHBx-LINE1b (622–646 bp, 50-UCGUGGAUGGAAUCGAAGAACA UUA-30). HKCI-4 cultured in complete AIM-V medium (Life Technologies) was transfected with siHBx-LINE1a or siHBx-LINE1b using Lipofectamine 2000 (Invitrogen). Stealth RNAi siRNA negative control was used as siMock. The efficacy of transcript knockdown was confirmed and monitored by RT-PCR. Effects of HBx-LINE1 knockdown on function, including cell viability by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), Matri-gel cell invasion, and Transwell cell migration, were examined. Details are described inSupplemental Experimental Procedures.
Expression Vector Constructions and Ectopic Transfection
A number of plasmids were constructed based on the complete sequence ob-tained of HBx-LINE1 (674 bp). These included a full-length HBx-LINE11–674
and variants HBx1–297
and LINE1295–674
, each of which constituted the viral and human component along with the GTG microhomology, and HBx-LINE11–312
*, which contained the HBx fragment and a partial sequence of LINE1 prior to a stop codon on the human portion. Details of the cloning maps are shown inFigure 5A, and primers are listed inSupplemental Experi-mental Procedures. Each plasmid was cloned into a pEGFP-C2 expression vector (Clontech) with an EGFP sequence to examine tag-fused protein trans-lation. The series of plasmids was also cloned into another derivative, pEGFP-C2R(-), with a deleted EGFP-coding sequence for use in transfection assays. A mutant HBx-LINE1 with a stop codon introduced into the HBx region was also constructed and is shown inFigure 6A. This aimed at producing a full-length RNA transcript in the absence of encoded protein. The HBx-LINE1 mutant was cloned by site-directed mutagenesis using the Transformer Site-Directed Mutagenesis Kit (Clontech) according to the manufacturer’s instruc-tions. The HBx-LINE1 construct was used as a template for annealing of the mutagenic primer (50-GGCTGCTAGGGTGTGATGCCAACTGGATCCT-30; the stop codon is underlined). One base substitution was introduced into the HBx region, resulting in only a small peptide of 5 amino acids being expressed from the HBx-LINE1 construct. This construct is referred to as the HBx-LINE1 stop mutant or HBx-LINE1mutant
. All cloned plasmids in pEGFP-C2 and pEGFP-C2R(-) were sequence verified and their expression was confirmed by RT-PCR in transfected cells. Expression vectors were transfected into L02, HKCI-9, and HKCI-1 using Lipofectamine 2000 (Invitrogen).
Liver-Specific Transgenic HBx-LINE1 Mice
Full-length HBx-LINE1 was cloned into an expression vector containing a liver-specific transthyretin promoter to generate an HBx-LINE1 transgene. Full-length HBx-LINE1 was amplified using forward and reverse primers containing the NruI restriction site. Transgenic mice were made following the standard protocol outlined (Hogan et al., 1986), and three strains were developed for the purpose of hybrid HBx-LINE1 investigations. PCR of tail DNA was em-ployed to screen for positive transgenic mice. Mice of age 15 days received a single intraperitoneal injection of DEN (5 mg/kg body weight; Sigma). After 8 months, mice were sacrificed and liver RNA was extracted to confirm HBx-LINE1 expression. The presence of surface nodules in HBx-LINE1 and wild-type transgenic mice was evaluated and compared. Development of HCC was confirmed histologically. All experimentation was conducted in accordance with guidelines of the Animal Experimentation Ethics Committee of the Chinese University of Hong Kong.
Statistics
Data are presented as mean ± SEM from at least three independent experi-ments. Kaplan-Meier survival plot and log-rank test were performed, together
with univariate and multivariate Cox regression analysis, to establish prog-nostic significance of HBx-LINE1 in patients. Student’s t test compared experimental data obtained between groups. A p value less than 0.05 was considered statistically significant.
Other Experimental Assays
Other supporting verifications and functional assays are described in Supple-mental ExperiSupple-mental Procedures.
ACCESSION NUMBERS
Sequencing data on six HBV-positive HCC cell lines have been deposited in the Sequence Read Archive under accession number SRP023539. SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures, five figures, and four tables and can be found with this article online at http://dx.doi.org/10.1016/j.ccr.2014.01.030.
AUTHOR CONTRIBUTIONS
C.-C.L., T.S., A.K.K.C., M.H., A.M.W., and N.N.C. performed biological exper-iments, conducted immunofluorescence work, PCR, and Sanger sequencing, and analyzed and interpreted data. J.-W.L. and T.-F.C. carried out analysis on transcriptome and bioinformatic studies. A.W.H.C., P.B.S.L., and K.-F.T. secured surgical liver cancer materials and conducted pathological examina-tion of patient samples and histologic examinaexamina-tion of tissues from transgenic mice. A.K.K.C. and P.-S.L. generated transgenic mice. R.W.M.L. and J.H.M.T. carried out immunohistochemical staining and in situ hybridization of tissue samples. H.L.Y.C. helped design some experiments and provided technical advice. N.W. conceived the study, designed experiments, and analyzed and interpreted data. T.-F.C. and N.W. wrote the paper.
ACKNOWLEDGMENTS
The authors thank Wah Ng, Grace Song, and Dr. Alfa Bai for technical assis-tance. We are also thankful to Prof. Alice Wong for discussions, Dr. T.E. Van Dyke for providing the transthyretin expression vector, Prof. Susanna Lee for provision of the transgenic facility, and Prof. David I. Smith for inspiring the research. This work was supported by the Theme-Based Research Scheme of the Hong Kong Research Grants Council (T12-403/11) and General Research Fund of the Hong Kong Research Grants Council (CUHK 476308). Received: September 6, 2013
Revised: December 7, 2013 Accepted: January 31, 2014 Published: February 27, 2014 REFERENCES
Braconi, C., Valeri, N., Kogure, T., Gasparini, P., Huang, N., Nuovo, G.J., Terracciano, L., Croce, C.M., and Patel, T. (2011). Expression and functional role of a transcribed noncoding RNA with an ultraconserved element in hepa-tocellular carcinoma. Proc. Natl. Acad. Sci. USA108, 786–791.
Bre´chot, C., Gozuacik, D., Murakami, Y., and Paterlini-Bre´chot, P. (2000). Molecular bases for the development of hepatitis B virus (HBV)-related hepa-tocellular carcinoma (HCC). Semin. Cancer Biol.10, 211–231.
Burns, K.H., and Boeke, J.D. (2012). Human transposon tectonics. Cell149, 740–752.
Cabili, M.N., Trapnell, C., Goff, L., Koziol, M., Tazon-Vega, B., Regev, A., and Rinn, J.L. (2011). Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev.25, 1915–1927.
Chan, K.Y., Lai, P.B., Squire, J.A., Beheshti, B., Wong, N.L., Sy, S.M., and Wong, N. (2006). Positional expression profiling indicates candidate genes in deletion hotspots of hepatocellular carcinoma. Mod. Pathol.19, 1546–1554.
Chisari, F.V., and Ferrari, C. (1995). Hepatitis B virus immunopathogenesis. Annu. Rev. Immunol.13, 29–60.
Cohen, S.B., Graham, M.E., Lovrecz, G.O., Bache, N., Robinson, P.J., and Reddel, R.R. (2007). Protein composition of catalytically active human telome-rase from immortal cells. Science315, 1850–1853.
Devereux, T.R., Stern, M.C., Flake, G.P., Yu, M.C., Zhang, Z.Q., London, S.J., and Taylor, J.A. (2001). CTNNB1 mutations andb-catenin protein accumula-tion in human hepatocellular carcinomas associated with high exposure to aflatoxin B1. Mol. Carcinog.31, 68–73.
Ding, D., Lou, X., Hua, D., Yu, W., Li, L., Wang, J., Gao, F., Zhao, N., Ren, G., Li, L., and Lin, B. (2012). Recurrent targeted genes of hepatitis B virus in the liver cancer genomes identified by a next-generation sequencing-based approach. PLoS Genet.8, e1003065.
El-Serag, H.B., and Rudolph, K.L. (2007). Hepatocellular carcinoma: epidemi-ology and molecular carcinogenesis. Gastroenterepidemi-ology132, 2557–2576. Feitelson, M.A., and Lee, J. (2007). Hepatitis B virus integration, fragile sites, and hepatocarcinogenesis. Cancer Lett.252, 157–170.
Fujimoto, A., Totoki, Y., Abe, T., Boroevich, K.A., Hosoda, F., Nguyen, H.H., Aoki, M., Hosono, N., Kubo, M., Miya, F., et al. (2012). Whole-genome sequencing of liver cancers identifies etiological influences on mutation pat-terns and recurrent mutations in chromatin regulators. Nat. Genet. 44, 760–764.
Gozuacik, D., Murakami, Y., Saigo, K., Chami, M., Mugnier, C., Lagorce, D., Okanoue, T., Urashima, T., Bre´chot, C., and Paterlini-Bre´chot, P. (2001). Identification of human cancer-related genes by naturally occurring hepatitis B virus DNA tagging. Oncogene20, 6233–6240.
Greene, F.L., Page, D.L., Fleming, I.D., Fritz, A., Balch, C.M., Haller, D.G., and Morrow, M., eds. (2002). AJCC Cancer Staging Manual, Sixth Edition (New York: Springer-Verlag).
Hastings, P.J., Ira, G., and Lupski, J.R. (2009). A microhomology-mediated break-induced replication model for the origin of human copy number varia-tion. PLoS Genet.5, e1000327.
Hogan, B., Costantini, F., and Lacy, E. (1986). Manipulating the Mouse Embryo: A Laboratory Manual. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).
Huang, J., Deng, Q., Wang, Q., Li, K.Y., Dai, J.H., Li, N., Zhu, Z.D., Zhou, B., Liu, X.Y., Liu, R.F., et al. (2012). Exome sequencing of hepatitis B virus-asso-ciated hepatocellular carcinoma. Nat. Genet.44, 1117–1121.
Jiang, Z., Jhunjhunwala, S., Liu, J., Haverty, P.M., Kennemer, M.I., Guan, Y., Lee, W., Carnevali, P., Stinson, J., Johnson, S., et al. (2012). The effects of hepatitis B virus integration into the genomes of hepatocellular carcinoma patients. Genome Res.22, 593–601.
Kremsdorf, D., Soussan, P., Paterlini-Brechot, P., and Brechot, C. (2006). Hepatitis B virus-related hepatocellular carcinoma: paradigms for viral-related human carcinogenesis. Oncogene25, 3823–3833.
Levin, H.L., and Moran, J.V. (2011). Dynamic interactions between transpos-able elements and their hosts. Nat. Rev. Genet.12, 615–627.
Li, J.W., Wan, R., Yu, C.S., Co, N.N., Wong, N., and Chan, T.F. (2013). ViralFusionSeq: accurately discover viral integration events and reconstruct fusion transcripts at single-base resolution. Bioinformatics29, 649–651. Matouk, I.J., DeGroot, N., Mezan, S., Ayesh, S., Abu-lail, R., Hochberg, A., and Galun, E. (2007). The H19 non-coding RNA is essential for human tumor growth. PLoS ONE2, e845.
Matsubara, K., and Tokino, T. (1990). Integration of hepatitis B virus DNA and its implications for hepatocarcinogenesis. Mol. Biol. Med.7, 243–260.
Murakami, Y., Saigo, K., Takashima, H., Minami, M., Okanoue, T., Bre´chot, C., and Paterlini-Bre´chot, P. (2005). Large scaled analysis of hepatitis B virus (HBV) DNA integration in HBV related hepatocellular carcinomas. Gut54, 1162–1168.
Ørom, U.A., Derrien, T., Beringer, M., Gumireddy, K., Gardini, A., Bussotti, G., Lai, F., Zytnicki, M., Notredame, C., Huang, Q., et al. (2010). Long noncoding RNAs with enhancer-like function in human cells. Cell143, 46–58.
Pang, E., Wong, N., Lai, P.B., To, K.F., Lau, W.Y., and Johnson, P.J. (2002). Consistent chromosome 10 rearrangements in four newly established human hepatocellular carcinoma cell lines. Genes Chromosomes Cancer 33, 150–159.
Panzitt, K., Tschernatsch, M.M., Guelly, C., Moustafa, T., Stradner, M., Strohmaier, H.M., Buck, C.R., Denk, H., Schroeder, R., Trauner, M., and Zatloukal, K. (2007). Characterization of HULC, a novel gene with striking up-regulation in hepatocellular carcinoma, as noncoding RNA. Gastroenterology132, 330–342.
Polette, M., Mestdagt, M., Bindels, S., Nawrocki-Raby, B., Hunziker, W., Foidart, J.M., Birembaut, P., and Gilles, C. (2007).b-catenin and ZO-1: shuttle molecules involved in tumor invasion-associated epithelial-mesenchymal transition processes. Cells Tissues Organs185, 61–65.
Sa´nchez-Tillo´, E., de Barrios, O., Siles, L., Cuatrecasas, M., Castells, A., and Postigo, A. (2011).b-catenin/TCF4 complex induces the epithelial-to-mesen-chymal transition (EMT)-activator ZEB1 to regulate tumor invasiveness. Proc. Natl. Acad. Sci. USA108, 19204–19209.
Shukla, R., Upton, K.R., Mun˜oz-Lopez, M., Gerhardt, D.J., Fisher, M.E., Nguyen, T., Brennan, P.M., Baillie, J.K., Collino, A., Ghisletti, S., et al. (2013). Endogenous retrotransposition activates oncogenic pathways in hepatocellu-lar carcinoma. Cell153, 101–111.
Sung, W.K., Zheng, H., Li, S., Chen, R., Liu, X., Li, Y., Lee, N.P., Lee, W.H., Ariyaratne, P.N., Tennakoon, C., et al. (2012). Genome-wide survey of recur-rent HBV integration in hepatocellular carcinoma. Nat. Genet.44, 765–769. Sze, K.M., Chu, G.K., Lee, J.M., and Ng, I.O. (2013). C-terminal truncated hep-atitis B virus X protein is associated with metastasis and enhances invasive-ness by C-Jun/matrix metalloproteinase protein 10 activation in hepatocellular carcinoma. Hepatology57, 131–139.
Toh, S.T., Jin, Y., Liu, L., Wang, J., Babrzadeh, F., Gharizadeh, B., Ronaghi, M., Toh, H.C., Chow, P.K., Chung, A.Y., et al. (2013). Deep sequencing of the hep-atitis B virus in hepatocellular carcinoma patients reveals enriched integration events, structural alterations and sequence variations. Carcinogenesis34, 787–798.
Whittaker, S., Marais, R., and Zhu, A.X. (2010). The role of signaling pathways in the development and treatment of hepatocellular carcinoma. Oncogene29, 4989–5005.
Wong, C.M., Fan, S.T., and Ng, I.O. (2001).b-catenin mutation and overex-pression in hepatocellular carcinoma: clinicopathologic and prognostic signif-icance. Cancer92, 136–145.
Xu, W., Seok, J., Mindrinos, M.N., Schweitzer, A.C., Jiang, H., Wilhelmy, J., Clark, T.A., Kapur, K., Xing, Y., Faham, M., et al.; Inflammation and Host Response to Injury Large-Scale Collaborative Research Program (2011). Human transcriptome array for high-throughput clinical studies. Proc. Natl. Acad. Sci. USA108, 3707–3712.
Yang, Z., Zhou, L., Wu, L.M., Lai, M.C., Xie, H.Y., Zhang, F., and Zheng, S.S. (2011). Overexpression of long non-coding RNA HOTAIR predicts tumor recur-rence in hepatocellular carcinoma patients following liver transplantation. Ann. Surg. Oncol.18, 1243–1250.