News and Views
Multivariate analysis and classi
fi
cation of the Apidima 2 cranium from Mani,
Southern Greece
Katerina Harvati
a,*, Chris Stringer
b, Panagiotis Karkanas
caDepartment of Early Prehistory and Quaternary Ecology, Senckenberg Center for Human Evolution and Paleoecology, Eberhard Karls Universität Tübingen, Rümelinstrasse 23, Tübingen 72070, Germany
bNatural History Museum, Cromwell Road, London SW7 5BD, UK
cEphoreia of Paleoanthropology and Speleology of Southern Greece, Ardittou 34b, Athens, Greece
a r t i c l e i n f o
Article history: Received 16 May 2010 Accepted 30 September 2010 Keywords: Homo neanderthalensis Homo heidelbergensis GreeceIn 1978, two well-preserved fossil human crania were discovered at the Apidima site, a karstic cave complex in the Mani peninsula (southern Peloponnese;Fig. 1), during excavations by the University of Athens Medical School (Pitsios, 1995; Harvati and Delson, 1999; Harvati et al., 2009). Together with the Petralona cranium, the Api-dima specimens represent the most important paleoanthropolog-icalfinds in Greece, and some of the most significant human fossil discoveries in south-eastern Europe, a region whose paleoanthro-pological record remains relatively unexplored. However, despite their importance, these specimens remain obscure; little has been published about them in the international literature, and detailed description and documentation are still pending.
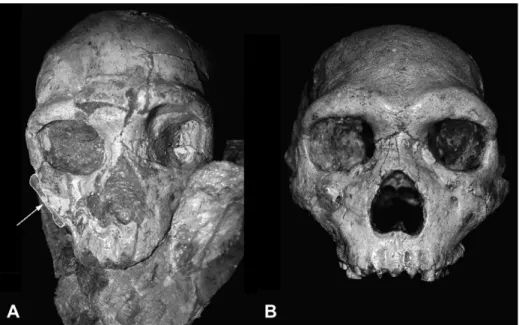
Of the two specimens, Apidima 2 is the better preserved, though somewhat distorted and showing multiple cracks (Fig. 2A). The occipital bone, parts of the temporals, as well as all of the teeth and part of the palate, are all absent. A fragment of the right mandibular ramus is preserved, attached to the encasing matrix and cranium in roughly anatomical position. Apidima 2 has been described as showing an elongated, low vault with a pronounced supra-orbital torus, a wide interorbital breadth, large rounded orbits, no canine fossa, a large nasal aperture, and a prognathic face (Harvati and Delson, 1999; Geanacos, 2001; Pitsios, 2002; Harvati et al., 2009). It is relatively gracile, and has been considered a female (Harvati and Delson, 1999; Pitsios, 2002; Harvati et al., 2009). Neanderthal affinities have been noted and the specimen has been proposed to
represent a pre-Neanderthal. Of particular interest is its possible relationship with the Petralona cranium from Northern Greece (Fig. 2B), and the suggestion that Apidima 2 may represent a female
Homo heidelbergensis comparable to Petralona in geological age. Apidima 1 is less complete and preserves only the posterior part of the neuro- and basicranium.
Attempts at identifying Apidima 2 to taxon have been stymied by the lack of both detailed description and extensive metric data. On the basis of its general morphology,Pitsios (1999) suggested that Apidima 2 lies on the Neanderthal lineage and proposed the nameHomo(sapiens)teanarius(Pitsios, 1995).Harvati and Delson (1999) suggested affinities with H. heidelbergensis (s.l.). Nine osteometric measurements published in a forensic report by Koutselinis et al. (1995)have been used as variables in previous metric analyses byManolis (1996)and, more recently, byHarvati et al. (2009). Both studies were inconclusive in their results, placing Apidima 2 either in an area of overlap between H. hei-delbergensis(s.l.) and early anatomically modern humans (Manolis, 1996), or betweenHomo neanderthalensis andH. heidelbergensis
(s.l.) (Harvati et al., 2009).
Given the importance of the Apidima specimens and the small amount of information on them that currently exists, we re-examine the metric data published for Apidima 2. We 1) test the original nine reported measurements for possible error associated with distortion; 2) correct the measurements for size, so that we can examine shape differences; and 3) take into account as many measurements and Middle-Late Pleistocene specimens as possible, in order to achieve the best resolution for classifying Apidima 2. We review the geology and chronology of the site in light of recent re-evaluation of regional geomorphology and discuss the implications of ourfindings for the interpretation of the Apidima remains. Materials and methods
Our comparative sample comprises nineteen fossil human crania (Table 1) on which all of the variables known for Apidima 2 can be measured (Table 2). Most data were collected by one of us (CS), although some were taken from the literature (Trinkaus, pers. comm., for Shanidar 1;Arsuaga et al., 1997). For the purposes of our analyses, we grouped the European Middle Pleistocene specimens
*Corresponding author.
E-mail address:[email protected](K. Harvati).
Contents lists available atScienceDirect
Journal of Human Evolution
j o u rn a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / j h e v o l
0047-2484/$esee front matterÓ2010 Elsevier Ltd. All rights reserved.
Petralona and Sima 5, together with the African specimen from Broken Hill, into H. heidelbergensis (s.l.), due to their well-docu-mented overall morphological similarities (e.g., Stringer, 1974; Stringer et al., 1979; Rightmire, 2001; Harvati et al., 2007, 2010; Harvati, 2009). We recognize that the latter two specimens may or may not belong to this taxon (e.g.,Hublin, 1998, 2009; Endicott et al., 2010; Harvati et al., 2010). Following previous views on its classifi -cation, Apidima 2 was alsoa prioriassigned toH. heidelbergensis(s.l.). We conducted row standardization in order to control for outlier values (and possible measurement errors due to breakage and/or distortion) in the reported measurements for Apidima 2 in comparison with the mean Neanderthal, H. heidelbergensis, and
H. sapiensvalues for the same measurements. This was achieved by dividing each value by the mean of all measurements for Apidima 2 and for each of the three samples used. Two measurements that
were found to be outliers were removed from the analysis. The remaining seven raw variables were adjusted for size by subtract-ing the log geometric mean of each variable for each individual from each log-transformed measurement. We then conducted a canonical variates analysis (CVA) on the seven size-corrected variables and classified the specimen using discriminant analysis. Finally, we used three of the apparently undistorted reported measurements (glabella-opisthocranion length, maximum cranial breadth, and basion-bregma height) to estimate cranial capacity for Apidima 2 using the modified formula for Pleistocene humans provided byOlivier and Tissier (1975):
Cranial capacity: 0:304ðlengthbreadthbasion bregma heightÞþ346cc
Figure 1.Left panel: map of the most important Paleolithic sites in Greece. Stars represent sites where human remains have been recovered. Image adapted fromHarvati et al. (2009). Right panel: the Apidima locality. Caves AeE are labelled, as is the approximate position of the 5 masl highstand.
Figure 2. A: The Apidima 2 cranium. Photograph of a cast of the original specimen before complete separation from the block of breccia. The arrow points to the mandibular fragment adhering to the matrix. B: the Petralona cranium. Photographs provided by and copyright Eric Delson.
Results
Row standardization
When the row-standardized measurements for Apidima 2 were compared to the equivalent mean values for the other samples, bizygomatic breadth (ZYB) and orbital height (OBH) stood out as outliers (Table 2). These variables are likely affected by distortion or breakage: distortion is apparent in the area of the left orbit, and breakage can be seen in the region of the right zygomatic bone (Fig. 2B). ZYB and OBH were removed from further analysis.
Canonical variates analysis
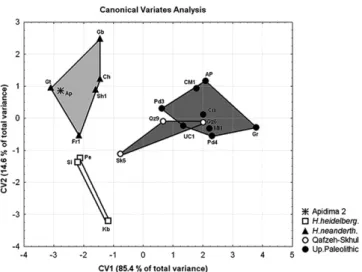
Canonical Axis 1 (85.4% of the total variance;Fig. 3) separatesH. sapiensfromH. heidelbergensis(s.l.) andH. neanderthalensis, and is influenced mostly by BBH, NPH, and GOL. Canonical Axis 2 (14.6% of the total variance) separates Neanderthals fromH. heidelbergensis
(s.l.) and is mostly influenced by XCB, NPH, and NLH. Apidima 2 was
a prioriassigned toH. heidelbergensis(s.l.) following previous views on its status as a pre-Neanderthal. Despite this assignment, Apidima 2 is pulled out of the range of the other threeH. heidelbergensis(s.l.)
specimens, and falls within the Neanderthal range on thefirst two canonical axes.
Discriminant analysis
Apidima 2 was classified as H. neanderthalensis (posterior probability 1.00). This analysis classified most specimens correctly to species with the exception of Skhul 5, which was misclassified as
H. heidelbergensis(s.l.) (posterior probability 0.85).
Cranial capacity
The estimated cranial capacity for the Apidima 2 individual is 1454 cc.
Geomorphological implications for the chronology of the Apidima site
The Apidima site comprisesfive caves (Caves AeE;Fig. 1) formed
in the Upper CretaceouseLate Eocene limestone on the coastal cliffs
of the inner Mani. Key in understanding the chronology of the specimens is the relationship of the “skull breccia” encasing the human specimens to other deposits within Cave A. Since the human remains were recovered at least 2.5 m above all otherfinds from Cave A (including Middle Paleolithic lithic artifacts [Kourstesi-Phi-lipaki, reported inHarvati and Delson, 1999], as well as Middle-Late Pleistocene faunal remains [Tsoukala, 1999]), they are most likely not contemporary with them (Lax, 1995). Attempts at radiometric dating have not been fruitful.Koutselinis et al. (1995)suggested an age between 300-100 ka, a chronology considered broadly in accordance with the specimens’perceived archaic morphology.
In the absence of radiometric dating and clearly associated archaeological and faunal evidence, understanding of the site’s chronology relies on evidence for Pleistocene sea level highstands present in the Mani region. The main formation of the caves is attributed to sea level highstands (Pitsios, 1985; Basiakos, 1993), which also produced notches and wave-cut platforms found at similar heights in the area (Fig. 1).
Cave B and the upper part of Cave E formed at a 12 masl high-stand (Pitsios, 1985), and were later filled by red Pleistocene breccia, forming a talus cone that fanned out along the cliff below Caves C and D, and sealed the entrances of Caves B and E. The talus cone was eroded and partially collapsed during a subsequent highstand, resulting in a reopening of Caves E and B. The main opening of Cave A seems to have occurred during a later and lower
Table 1
Specimens included in this study, the taxon to which they were assigned for the purposes of the canonical variates analysis (CVA), and the source of measurements used. Abbreviations used inFig. 3are shown in parentheses.
Specimen Taxon Measurements source Apidima 2 (Ap2) H. heidelbergensis(s.l.) Koutselinis et al. (1995)
Atapuerca Cranium 5 (Si) H. heidelbergensis(s.l.) Arsuaga et al. (1997)
Petralona (Pe) H. heidelbergensis(s.l.) CS Broken Hill (Kb) H. heidelbergensis(s.l.) CS Guattari (Gt) H. neanderthalensis CS Forbes Quarry 1 (Gb) H. neanderthalensis CS La Chapelle (Ch) H. neanderthalensis CS La Ferrassie 1 (Fr1) H. neanderthalensis CS
Shanidar 1 (Sh1) H. neanderthalensis E. Trinkaus (pers. comm.) Qafzeh 6 (Qz6) H. sapiens CS
Qafzeh 9 (Qz9) H. sapiens CS Skhul 5 (Sk5) H. sapiens CS Abri Pataud (AP) H. sapiens CS Cro Magnon 1 (CM1) H. sapiens CS Chancelade (Cn) H. sapiens CS Grimaldi 6 (Gr) H. sapiens CS Mladec1 (Ml1) H. sapiens CS Predmostí 3 (Pd3) H. sapiens CS Predmostí 4 (Pd4) H. sapiens CS Upper Cave 101 (UC1) H. sapiens CS
Table 2
Top: raw measurements (in mm) reported for Apidima 2 (in bold) and mean values forH. heidelbergensis(s.l.),H. neanderthalensis, early anatomically modern humans, and Upper Paleolithic samples. Bottom: row-standardized values of the same variables. The values reported for ZYB and OBH for Apidima 2 clearly stand out in comparison to the other row-standardized values for those measurements.
Glabella-Opisthoc. length (GOL) Maximum Cranial breadth (XCB) Basion-Bregma height (BBH) Bizygom. breadth (ZYB) Nasion-Prosth. height (NPH) Nasal height (NLH) Nasal breadth (NLB) Orbital breadth (OBB) Orbital height (OBH) Raw measurements Apidima 2 197.00 148.00 125.00 130.00 81.00 57.00 31.00 41.00 41.00 MeanH. heidelbergensis(s.l.) 200.17 147.00 128.33 150.00 89.63 60.40 35.03 46.13 34.83 MeanH. neanderthalensis 203.04 153.50 129.70 143.60 84.24 61.08 33.12 44.96 36.82 Mean early anatomically modern 195.17 143.00 132.00 138.67 73.67 53.67 31.10 44.00 32.83 Mean Upper Paleolithic 196.69 142.38 137.25 140.75 69.68 52.44 25.95 41.49 30.29
Row standardized
Apidima 2 2.08 1.57 1.32 1.37 0.86 0.60 0.33 0.43 0.43
MeanH. heidelbergensis(s.l.) 2.02 1.48 1.30 1.51 0.90 0.61 0.35 0.47 0.35
MeanH. neanderthalensis 2.05 1.55 1.31 1.45 0.85 0.62 0.33 0.45 0.37
Mean early anatomically modern 2.08 1.52 1.41 1.48 0.79 0.57 0.33 0.47 0.35 Mean Upper Paleolithic 2.12 1.53 1.48 1.51 0.75 0.56 0.28 0.45 0.33
5 m highstand, which dissolved and eroded the red breccia as well as the underlying limestone (Basiakos, 1993). Cave A is tunnelled for several meters inside the red breccias (Fig. 3), and therefore could not have been filled with the cone sediments after its formation (contraLax, 1995). The“skull breccia”was found almost 7 m from the entrance of Cave A. Although no physical connection was observed, the excavators proposed that it was part of the talus cone breccia found in front of the cave, and through which the entrance of Cave A is tunnelled, because of sediment similarities (Pitsios, 1985). If so, the human crania must have been deposited
beforethe main opening phase of Cave A (Pitsios, 1985). At a later time, Middle Palaeolithic cultural remains were deposited on the
floor of the Cave A (see above).
It therefore seems that the opening of Cave A, as well as the exposure of the “skull breccia,” is related to the 5 m highstand. Geomorphological evidence of this highstand, and of the higher coastline at ca.12 m related to Caves B and E, is reported from several locations around the Mani (Kelletat and Gassert, 1975; Tsartsidou et al., 2002). These two highstands were believed in the past to represent the Neo-tyrrhenian and Eutyrrhenian sea levels, respec-tively (Kelletat and Gassert, 1975; Eutyrrhenian is correlated with MIS 5e, i.e., the last interglacial. Neo-tyrrhenian could represent MIS 5c [e.g.,Keraudren et al., 2000] or MIS 5a). However, recent radio-metric dates and stratigraphic correlations assign two outcrops representing the 5 m highstand to the last interglacial, i.e., the Eutyrrhenian rather than the Neo-tyrrhenian highstand (at the Kalamakia Cave, close to Apidima, and at Lakonis Cave on the eastern coast of Mani;Darlas, 2007; Elefanti et al., 2008). This new assess-ment agrees with the general consensus that the global sea level peaked at least 6 m higher than today, during the last interglacial (Kopp et al., 2009). Therefore, there is a strong possibility that Cave A was formed during the last interglacial. If this is correct, Caves B and E were formed by an earlier and even higher sea level occur-rence, possibly during MIS 11 (w430e400 ka), the longest and one
of the warmest interglacials (Augustin et al., 2004), whose highstand is thought to range from close to the present sea level up to 20 masl (Waelbroeck et al., 2002; Olson and Hearty, 2009).
A correlation of the 12 m paleocoastline with MIS 11 puts the age of the Apidima crania between 400-130 ka.Rondoyanni et al. (1995), on the basis offield observations and tentative correlations with other studied areas, constrained the formation of the talus cone to the“Riss,”and probably late“Riss,”which is broadly correlated with
MIS 6 (190e130 ka). Given the tectonic instability of the area,
however, the possibility that the 5 m and 12 m paleocoastlines correlate with the Neo-tyrrhenian (MIS 5c) and Eutyrrhenian (MIS 5e) highstands cannot be excluded. This would constrain the age of the crania to be between 115 ka and 105 ka.
Discussion
Previous assessments of the phylogenetic position of Apidima 2, and the less complete Apidima 1, have relied on a few short pub-lished descriptions (e.g., Koutselinis et al., 1995; Pitsios and Liebhaber, 1995; Pitsios, 1999, 2002), brief visual inspections (e.g., Harvati and Delson, 1999), and morphometric analyses of a few published measurements (Manolis, 1996; Harvati et al., 2009). Apidima 2 has been described as showing some Neanderthal-like facial features, and possibly representing an early member of the Neanderthal lineage closely related to Petralona and likely dating to the Middle Pleistocene (Harvati and Delson, 1999; Pitsios, 1999, 2002; Harvati et al., 2009).
Our reassessment of the taxonomic status of Apidima 2 also relied on published measurements. However, by identifying and excluding variables affected by distortion or breakage, we were able to remove a possible source of error, which might have confounded previous results (Manolis, 1996; Harvati et al., 2009). We also adjusted the measurements for size and maximized both the comparative sample and the variables used. Contrary to our expec-tations, Apidima 2 did not show affinities withH. heidelbergensis(s.l.) or with the Petralona cranium in particular, but aligned more closely with our Neanderthal sample despite having been groupeda priori
withH. heidelbergensis(s.l.). A brief comparison of the Apidima 2 frontal view to that of Petralona (Fig. 2) confirms several differences between the two specimens, including a more Neanderthal-like, relatively narrower face and higher cranium in Apidima (although the exact position and orientation of the two specimens could not be controlled and is not identical). However, Apidima 2 appears to lack the extremely derived Neanderthal nasal morphology, including the very high and protruding nasal bridge. Our estimate of cranial capacity at 1454 cm3places Apidima 2 within the Neanderthal range of variation (1200e1700 cm3;Holloway et al., 2004) and above the
values reported for Middle Pleistocene specimens (Holloway et al., 2004), although Sima 4 approaches this value at 1390 cm3 (Arsuaga et al., 1997; Holloway et al., 2004). Similar cranial capacity values are reported for Ehringsdorf (1450 cm3), Reilingen (1430 cm3), and Krapina B (1450 cm3). This estimate agrees with our comparative analysis in pointing to Neanderthal affinities, but is limited in that it is based on three external cranial variables and is not a direct measurement of cranial capacity.
In conclusion, our observations suggest that a late Middleeearly
Late Pleistocene chronology is most consistent with the morphology the Apidima fossils. Our review of the site’s geological setting points to 400e105 ka as the most likely period for the
accumulation of the relevant sediments. This time bracket is very wide, but is likely further constrained to the latter part of this time period. It should be pointed out that our results rely on only a small number of metric data without detailed morphological description, and should therefore be considered tentative. A detailed descrip-tion and a more exhaustive comparative analysis based on addi-tional linear or 3-D data, is crucial to arriving at a conclusive assessment. Nevertheless, the Neanderthal affinities indicated here place the Apidima site and specimens among the Middle Paleolithic localities known from the immediate geographic vicinity, which have yielded a number of well-documented, though fragmentary, fossil human remains (Darlas and de Lumley, 1998; Panagopoulou et al., 2002e2004; Harvati et al., 2003; Richards et al., 2008; Smith et al., 2009).
Figure 3.Canonical variates analysis, canonical axes 1 and 2. Abbreviated specimen labels as inTable 1. Although early modern humans from Skhul and Qafzeh are labelled separately from Upper Paleolithic specimens, the two samples were pooled in the analysis asH. sapiens.
Acknowledgements
We thank Eric Delson for helpful discussion and for kindly providing the photographs used inFig. 2. Constructive comments were also provided by Andreas Darlas. We are grateful to the Ephoreies of Paleoanthropology and Speleology (of Southern and of Northern Greece) of the Greek Ministry of Culture and to the University of Athens. This work was supported by Eberhard Karls Universität Tübingen and the Senckenberg Center for Human Evolution and Paleoecology. Chris Stringer is a member of the Ancient Human Occupation of Britain project, funded by the Leverhulme Trust.
References
Arsuaga, J.L., Martinez, I., Gracia, A., Lorenzo, C., 1997. The Sima de los Huesos crania (Sierra de Atapuerca, Spain). A comparative study. J. Hum. Evol. 33, 219e281.
Augustin, L., Barbante, C., Barnes, P.R.F., Barnola, J.M., Bigler, M., Castellano, E., Cattani, O., Chappellaz, J., Dahl-Jensen, D., Delmonte, B., Dreyfus, G., Durand, G., Falourd, S., Fischer, H., Flückiger, J., Hansson, M.E., Huybrechts, P., Jugie, G., Johnsen, S.J., Jouzel, J., Kaufmann, P., Kipfstuhl, J., Lambert, F., Lipenkov, V.Y., Littot, G.C., Longinelli, A., Lorrain, R., Maggi, V., Masson-Delmotte, V., Miller, H., Mulvaney, R., Oerlemans, J., Oerter, H., Orombelli, G., Parrenin, F., Peel, D.A., Petit, J.-R., Raynaud, D., Ritz, C., Ruth, U., Schwander, J., Siegenthaler, U., Souchez, R., Stauffer, B., Steffensen, J.P., Stenni, B., Stocker, T.F., Tabacco, I.E., Udisti, R., van de Wal, R.S.W., van den Broeke, M., Weiss, J., Wilhelms, F., Winther, J.-G., Wolff, E.W., Zucchelli, M., 2004. Eight glacial cycles from an Antarctic ice core. Nature 429, 623e628.
Basiakos, Y., 1993. Xronolόghsh apoliqumάsun sphlaíun kai sphlaioapoq3mάsun m3 sh mέqodo soy synsonismoύ sh2 hl3ksronikή2 ssro4ormή2 kai m3lέsh mor4ología2 ypόg3ioy karsskaisun sc3sikώn radiom3srikώn kai g3ulogikώn synqhkώn s3 sphlaiop3ribάllonsash2 p3riocή2 DyroύMάnh2. Ph.D. Disser-tation, University of Athens.
Darlas, A., 2007. The Mousterian of Greece under the light of recent research. L’Anthropologie 111, 346e366.
Darlas, A., de Lumley, H., 1998. Fouilles franco-helléniques de la grotte de Kalamakia (Aréopolis; Péloponnèse). Bull. Corresp. Hell. 122, 655e661.
Elefanti, P., Panagopoulou, E., Karkanas, P., 2008. The transition from the Middle to the Upper Palaeolithic in the southern Balkans: the evidence from the Lakonis I Cave, Greece. Eur. Prehist. 5, 85e95.
Endicott, P., Ho, S., Stringer, C., 2010. Using genetic evidence to evaluate four palaeoanthropological hypotheses for the timing of Neanderthal and modern human origins. J. Hum. Evol. 59, 87e95.
Geanacos, C., 2001. Report of a meeting.“Homo (sapiens) taenarius”, the youngest or the oldest Neanderthal in Europe? Hum. Evol. 16, 243e244.
Harvati, K., 2009. Petralona: link between Africa and Europe? In: Schepartz, L., Bourbou, C., Fox, S. (Eds.), New Directions in the Skeletal Biology of Greece Occasional Wiener Laboratory Series. ASCSA, Athens, pp. 31e49.
Harvati, K., Delson, E., 1999. Conference report: paleoanthropology of the Mani Peninsula (Greece). J. Hum. Evol. 36, 343e348.
Harvati, K., Panagopoulou, E., Karkanas, P., 2003. First Neanderthal remains from Greece: the evidence from Lakonis. J. Hum. Evol. 45, 465e473.
Harvati, K., Hublin, J.-J., Gunz, P., 2007. Evolution of Middle-Late Pleistocene human cranio-facial form. Am. J. Phys. Anthropol. S44, 126e127.
Harvati, K., Panagopoulou, E., Curtis, R., 2009. The paleoanthropology of Greece. Evol. Anthropol. 18, 131e143.
Harvati, K., Hublin, J.-J., Gunz, P., 2010. Evolution of Middle-Late Pleistocene human cranio-facial form: a 3-D approach. J. Hum. Evol. 59, 445e464.
Holloway, R.L., Broadfield, D.C., Yuan, M.S., 2004. Methods and materials of endocast analysis. In: Schwartz, J.H., Tattersal, I. (Eds.), The Human Fossil Record. Brain EndocastsdThe Paleoneurological Evidence, vol. 3. John Wiley and Sons, Inc.,
New Jersey, pp. 28e37.
Hublin, J.-J., 1998. Climatic changes, paleogeography and the evolution of the Neanderthals. In: Akazawa, T., Aoki, K., Bar-Yosef, O. (Eds.), Neandertals and Modern Humans in Western Asia, pp. 295e310. New York.
Hublin, J.-J., 2009. The origin of Neandertals. Proc. Natl. Acad. Sci. U. S. A. 106, 16022e16027.
Kelletat, D., Gassert, D., 1975. Quartärmorphologische Untersuchungen im Küs-tenraum der Mani-Halbinsel, Peloponnes. Zeitschrift für Geomorphologie 22, 8e56.
Keraudren, B., Dalongeville, R., Bernier, P., Caron, V., Renault-Miskovsky, J., 2000. Le Pléistocène supérieur marin (Tyrrhénien) en Crète nord-orientale (Grèce). (The Marine Upper Pleistocene (Tyrrhenian) in north-eastern Crete (Greece)). Géo-morphologie: relief, processus, environnement 3, 177e190.
Kopp, R.E., Simons, F.J., Mitrovica, J.X., Maloof, A.C., Oppenheimer, M., 2009. Prob-abilistic assessment of sea level during the last interglacial stage. Nature 462, 863e867.
Koutselinis, A., Dritsas, C., Pitsios, T.K., 1995. Iasrodikassikήdi3r3ύnhshsoy kraníoy LAO 1/S2apόso ApήdhmaLakunía2. Acta Anthropologica 1, 105e117.
Lax, E., 1995. Quaternary faunal remains from the cave site of Apidima (Lakonia, Greece). Acta Anthropol. 1, 127e156.
Manolis, S.K., 1996. Hellenic Late Pleistocene fossils. Anthropologie (Brno) 1e2,
89e97.
Olivier, G., Tissier, H., 1975. Determination of cranial capacity in fossil men. Am. J. Phys. Anthropol. 43, 353e362.
Olson, S.L., Hearty, P.J., 2009. A sustainedþ21 m sea-level highstand during MIS 11 (400 ka): direct fossil and sedimentary evidence from Bermuda. Quatern. Sci. Rev. 28, 271e285.
Panagopoulou, E., Karkanas, P., Kotjabopoulou, E., Tsartsidou, G., Harvati, K., Ntinou, M., 2002e2004. Late Pleistocene archaeological and fossil
human evidence from Lakonis cave, Southern Greece. J. Field Archeol. 2, 323e349.
Pitsios, T.K., 1985.Palaioanqrupologikέ2έr3yn32 ssh qέsh«Apήdhma»sh2Mέsa
Mάnh2. Arcaiología15, 26e33.
Pitsios, T.K., 1995. Paleoanthropological research at the cave site of Apidima, Laconia, Greece. Acta Anthropol. 1, 1e180.
Pitsios, T.K., 1999. Paleoanthropological research at the cave site of Apidima and the surrounding region (South Peloponnese, Greece). Anthropol. Anzeiger 57, 1e11.
Pitsios, T.K., 2002. The fossil hominidfindings from the Cave site of Apidima, South Peloponnese Greece. Colegium Anthropol. S26, 158.
Pitsios, T.K., Liebhaber, B., 1995. Research conducted in Apidima and the surrounding regioneTaenarios Man. Acta Anthropol. 1, 175e179.
Richards, M., Harvati, K., Grimes, V., Smith, C., Smith, T., Hublin, J.-J., Karkanas, P., Panagopoulou, E., 2008. Isotope evidence of Neanderthal mobility. J. Archaeol. Sci. 35, 1251e1256.
Rightmire, G.P., 2001. Patterns of hominid evolution and dispersal in the Middle Pleistocene. Quatern. Int. 75, 77e84.
Rondoyanni, T., Mettos, A., Georgiou, C., 1995. Geologicalemorphological
observa-tions in the greater Oitilo-Diros area, Mani. Acta Anthropol. 1, 93e102.
Smith, T.M., Harvati, K., Olejnizak, A.J., Reid, D.J., Hublin, J.-J., Panagopoulou, E., 2009. Dental development and enamel thickness in the Lakonis Neanderthal molar. Am. J. Phys. Anthropol. 138, 112e118.
Stringer, C.B., 1974. A multivariate study of the Petralona skull. J. Hum. Evol. 3, 397e404.
Stringer, C.B., Howell, F.C., Melentis, J., 1979. The significance of the fossil hominid skull from Petralona, Greece. J. Arch. Sci. 6, 235e253.
Tsartsidou, G., Gaki-Papanastasiou, K., Maroukian, H., 2002. Coastal evolution of the inner Lakonic Gulf in the late Pleistocene-Holocene period. In: Proceedings of the 6th Pan-Hellenic Geographical Conference of the Hellenic Geographical Society, Thessaloniki, Greece, pp. 380e387.
Tsoukala, E., 1999. Quaternary large mammals from the Apidima caves (Lakonia, S Peloponnese, Greece). Beitr. Paläont. 24, 207e229.
Waelbroeck, C., Labeyrie, L., Michel, E., Duplessy, J.C., McManus, J.F., Lambeck, K., Balbon, E., Labracherie, M., 2002. Sea-level and deep water temperature changes derived from benthic foraminifera isotopic records. Quatern. Sci. Rev. 21, 295e305.