Original Article
KLK4 is synchronically expressed to
CTSC in ameloblasts during amelogenesis molars
Lijie Wang1,2*, Xiaohua Xie1*, Yunduan Sun3, Jingyu Wang1, Yan Wang1, Xiumei Wang1
1Department of Stomatology, The 2nd Affiliated Hospital of Harbin Medical University, Harbin, Heilongjiang Province, China; 2Department of Stomatology, General Hospital of Daqing Oil Field, Daqing, Heilongjiang Prov-ince, China; 3The 1st Affiliated Hospital of Harbin Medical University, Harbin, Heilongjiang Province, China. *Equal contributors.
Received January 2, 2017; Accepted January 27, 2017; Epub May 1, 2017; Published May 15, 2017
Abstract: The aim of this study was to provide the evidence that the CTSC product, DPPI is the activator of KLK4 dur-ing amelogenesis by examindur-ing the expression pattern of KLK4 and CTSC in developing teeth. The 1st mandibular molars from E16.5 embryos and postnatal mice, and incisor from P5 mandibles were selected for the immunohisto-chemistry with antibodies against KLK4 and DPPI. The expression of KLK4 and CTSC were initiated from postnatal day 1 (P1) expressed in molar ameloblasts. From P5 on, KLK4 was distributed not only in ameloblasts, but also in the degenerating satellite reticulum of molar enamel organ. However, DPPI distribution is always restricted to the molar ameloblasts. In the P5 incisor, the distribution of KLK4 and DPPI were spatiotemporally consistent in the ameloblasts. The coincidence of KLK4 and DPPI distribution in ameloblasts strongly suggested that DPPI activated KLK4. The distribution of KLK4 in satellite reticulum suggested that KLK4 most likely plays a proteolytic role in degeneration of enamel organ.
Keywords: Amelogenesis, DPPI, KLK4, CTSC, papillon-lefèvre syndrome
Introduction
Papillon-Lefèvre syndrome (PLS) is a rare autosomal recessive disorder characterized by palmoplantar hyperkeratosis, prepubertal peri-odontitis and premature loss of both deciduous and permanent teeth. The mutations of Cathe- psin C gene (CTSC) are attributed to PLS [1]. A cysteine lysosomal protease belonging to the papaya superfamily is encoded by CTSC, name-ly dipeptidyl-peptidase I (DPPI) which hydroname-lyz- hydrolyz-es proteins or peptidhydrolyz-es in lysosomhydrolyz-es by its endopeptidase and carboxypeptidase activities [2, 3]. In PLS patients, dysfunction of DPPI decreases protein degradation resulting in hy- perkeratosis [4]. Meanwhile, DDPI also plays a key role in activating chymotrypin-like serine proteases by removing one or more N-terminal dipeptides from zymogen [5]. PLS patients ex- hibited reduced activities of polymorphonucle-ar leukocyte (PMN)-derived serine proteinases, such as elastase, cathepsin G and proteinase 3, which in turn decreased the production of antimicrobial peptide and the neutralization of
leukotoxin from anaerobic pathogens [6]. These findings explained how and why the mutations of CTSC or the loss of DPPI lead to hyperkerato-sis, prepubertal periodontitis and premature tooth loss in PLS. However, not all of the mani-festations in PLS patients or CTSC deficient mice was verified to result from this mecha-nism. For example, micro-hardness testing con-firmed that the enamel of CTSC knock-out mice is significantly softened compared with the nor-mal littermates [7]. Whether the serine prote-ase activity of DPPI contributes to the soft enamel of CTSC deficient mice is an attracting concern in PLS study.
and responsible for the ultimate removal of matrix proteins from enamel rods, the activity of KLK4 is regarded more critical for enam-el maturation and hardness [10, 12]. KLK4 is produced in a pre-pro-enzyme form in endoplasm reticulum, secreted into extracel-lular space as a pro-enzyme after the removal of signal peptide, and transformed into active serine proteinase by cutting N-terminal propeptide off [13-15]. However, the extracellular activator of KLK4 is still unknown. Although MMP20 could activate KLK4 in vitro, their short overlap during amelogene-sis and the activation of KLK4 in MMP20 knock-out mice revealed that the in vivo activation of KLK4 is performed by enzymes other than MMP20 [16].
As a serine proteinase activating extracellular zymogens, DPPI was found to be secreted into extra- cellular space [17]. Since the re- duced hardness of enamel was detected in both CTSC and KLK4 deficiency mice [7], DPPI was pro-posed as the activator of KLK4 during amelogenesis. Examining the expression pattern of CTSC and KLK4 during amelogenesis is the first step to verify this hypoth-esis. Only when the expression of KLK4 is spatiotemporally consis-tent with that of CTSC, the possi-bility of DPPI activating KLK4 can be established.
Materials and methods
[image:2.612.91.361.65.640.2]Animals and tissue preparations All the experiments were approv- ed by animal ethics committee of Harbin medical university and co- nducted in accordance with the requirements of ethics commit-tee. C57/BL6 male and female mice was mating and the morning vaginal plug appeared was record-ed as embryonic day 0.5 (E0.5) to get E16.5 embryos. The pregnant
mice were sacrificed by cervical dislocation and the embryonic mandibles were fixed in 4% poly-formaldehyde for 24 h at 4°C. For postnatal tissue preparation, the mandibles were separated from the mice after 10% chloral hydra- te intraperitoneal anesthesia and fixed in 4% polyformaldehyde for 24 h at 4°C. Then, the postnatal mandibles were put into 8% EDTA for decalcification. Both the em- bryonic and postnatal mandibles were dehydrated by gradient et- hanol and embedded in paraffin after xylene treatment. All sam-ples were sectioned into 4 um slices, adhered to polylysine am- monia acid treated slides, dried at 37°C and finally treated at 70°C oven overnight.
Antibodies
For immunohistochemical exami-nation, rabbit polyclonal antibody against KLK4 (Biorbyt, CA, USA) and rat polyclonal antibody ag- ainst DPPI were applied (Santa Cruz, CA, USA). Secondary antibo- dies against rabbit and rat conju-gated with horseradish peroxi-dase were purchased from ZSGB-BIO Inc, Beijing, China.
Immunohistochemistry
Paraffin sections were deparaf-finized in xylene and rehydrated with gradient alcohol. Endogenous peroxidase was inactivated in 3% hydrogen peroxide at room tem-perature for 10 minutes. Then, the sections were rinsed with distilled water and placed in phosphate buffer saline (PBS) for 5 minutes. Sections citrate sustained-release liquid (PH6.0, 10-2 M) were heated by microwave for antigen retrieval (high heat for 2 minutes, and then low heat for 5 minutes for KLK4; high heat for 5 minutes for DPPI). After cooled down to room
[image:3.612.91.359.70.639.2]perature and rinsed with PBS, the sections were blocked with bovine serum for 20 min and subsequently incubated with KLK4 antibody (1:100) or DPPI antibody (1:50) overnight at
[image:4.612.91.526.68.573.2]4°C. Then, the sections were rinsed with PBS and incubated with secondary antibody for 20 min at room temperature before develop the color with freshly prepared DAB.
Results
Distribution of KLK4 in developing molar germs
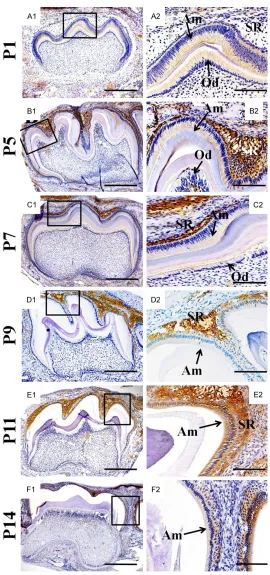
The immunohistochemistry examinations sh- own that at the E16.5 day, neither the pre-ame-loblasts nor pre-odontoblasts of the mouse molar germs exhibited the KLK4 expression (data not shown). In the 1st molar at P1, the KLK4 expression was first detected in the ame-loblasts and odontoblasts at secretory stage, but absent from the satellite reticulum of enamel organ (Figure A1 and A2). At the P5 molar enamel organ, the KLK4 distribution was detected not only in the ameloblasts at mature stage, but also even stronger in the satellite reticulum (Figure B1 and B2). However, the KLK4 distribution in the P5 odontoblasts was concentrated at the cusps region, instead of the entire layer of odontoblasts (Figure C1 and
C2). In the P7 molar germs, the KLK4 signals in odontoblasts almost diminished, but insisted in the ameloblasts and the degenerating satel- lite reticulum (Figure D1 and D2). From P9 on, the intensity of KLK4 became weaker and weaker in both the ameloblasts andsatellite reticulum (Figure E1 and E2). To the P14, the KLK4 signal was almost undetectable in amelo-blasts, though still obvious in degeneratedsat-ellite reticulum (Figure F1 and F2).
Expression of CTSC and distribution of DPPI in developing molar germs
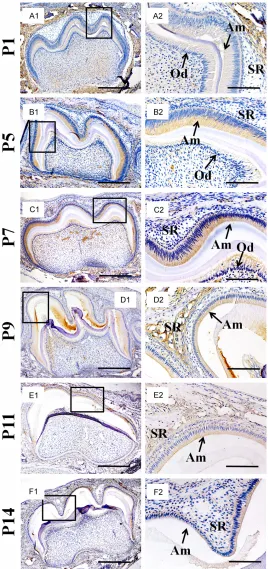
Similar to the expression of KLK4, the immuno-histochemistry with antibody against DPPI gave rise to no signal in the E16.5 molar germs (data not shown). Even at the P1 when DPPI was intensively detected in the alveolar bone sur-rounding molar germs, there was only slight sig-nals in the ameloblasts and odontoblasts (Figure 2A1 and 2A2). However, from P5 to P7, the DPPI intensity decreased dramatically in the alveolar bone and almost diminished in odontoblasts, while its intensity increased in the ameloblasts (Figure 2B1-C2). Such increase was reversed at P9 and decreased gradually until P14. Meanwhile, the surrounding alveolar bone, odontoblasts and degenerating satellite reticulum were almost devoid of DPPI distribu-tion (Figure 2D1-F2).
Distributions of KLK4 and DPPI in postnatal incisor
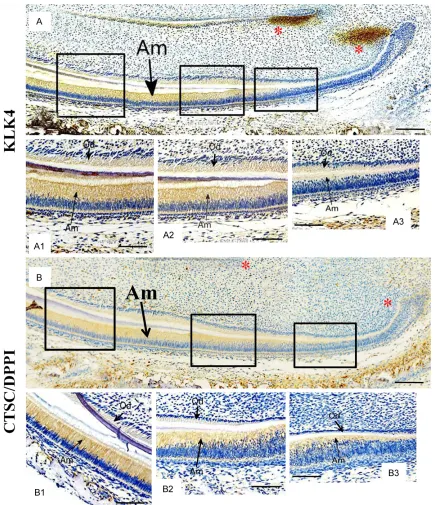
The P5 incisors which contain ameloblasts from progenitor to mature status were applied to the examination of the KLK4 and CTSC expression. In the para-sagittal section of the P5 incisor, the KLK4 expression was initiated the pre-secretory ameloblasts which started the polarization to transform into high colum-nar morphology (Figure 3A and 3A3). Intrigu- ingly, although the epithelial cells between the cervical loop and the polarizing domain were devoid of KLK4 expression, the mesenchyme adjacent to cervical loop exhibited a strong staining of KLK4 signal (Figure 3A). In contrast, both the polarized ameloblasts and odonto-blasts showed the continuous expression of KLK4 (Figure 3A2 and 3A1). On the other hand, the DPPI distribution exhibited the consistent pattern with that of KLK4 in the P5 incisor, except was absent from the mesenchyme adja-cent to cervical loop (Figure 3B and 3B1-B3). Noticeably, both KLK4 and CSTC displayed the more robust expression in ameloblasts than in odontoblasts (Figure 3A, 3A1-A3 and
3B, 3B1-B3).
Discussion
once thought as the activator of KLK4, howev-er, there were emerging evidences denying this notion. First, MMP20 and KLK4 expression overlap few chronically [20, 22]; second, KLK4 is normally activated in MMP20 knock-out mice and also able to inactivate MMP20 in vitro [16, 23]; finally, the severity of amelogenesis impe-fecta in MMP20 null mouse is milder than KLK4 deficient mouse [12, 24]. Therefore, KLK4 is activated by proteinases other than MMP20. Our study showed that the lysosomal protein-ase, DPPI encoded by CTSC exhibits the expres-sion pattern in ameloblasts identical to KLK4. Additionally, KLK4 expression reached the climax at P5 to P7, which was consistent with the DPPI expression. Previous study demon-strated that the reduced enamel hardness was observed in KLK4 deficient mouse as well as CTSC deficient mouse [7]. These findings strongly suggested that DPPI could serve as the activator of KLK4. However, there is few reports on the reduced enamel hardens in the PLS patients, which may result from the early loss of teeth. So it still needs further investiga-tion to address if the hardness of PLS teeth is reduced.
The most interesting discovery in our study is that the distribution of KLK4 is much broader than that of DPPI confined in ameloblasts. Considering the previous study showing that KLK4 was only transcribed in ameoloblasts [22], the extended spread of KLK4 to the degenerating enamel organ suggested that KLK4 was supposed to be activated by DPPI around ameloblasts and then, transported by other binging proteins to satellite reticulum. The broader distribution of KLK4 also suggest-ed that KLK4 was most likely play a proteolytic role in the degeneration of enamel organ. The other intriguingly findings in our study is the detection of KLK4-expressing domains sur-rounding the cervical loops of incisor. As shown in previous studies, the mesenchyme adjacent to cervical loop of incisor acts as the stem cell pool of odontoblasts [25]. Our discovery implies that KLK4 contributes to the maintenance or renewal of the mesenchymal stem cell pool, and may fulfill the role through its protelytic function. On the other hand, DPPI was not detected in the same mesenchymal region, suggesting that DPPI is not the sole enzyme capable of activating KLK4.
Acknowledgements
This study is funded by: the Sciences and technology program of Education Bureau in Heilongjiang Province (12541499: The relation-ship between AEG-1 expression and prognosis in oral squamous cell cancer), Funds on leading candidates for future leaders in Heilongjiang Province, and Natural Science Foundation of Heilongjiang Province of China (Grant H2014- 18) and the grant from Natural Science Foundation of China (NSFC NO. 81600848).
Disclosure of conflict of interest
None.
Address correspondence to: Dr. Xiumei Wang, De- partment of Stomatology, The 2nd Affiliated Hospital of Harbin Medical University, 246 Xuefu Road, Harbin 150086, Heilongjiang, China. Tel: (+86) 451-86605544; E-mail: [email protected]
References
[1] Toomes C, James J, Wood AJ, Wu CL, McCor-mick D, Lench N, Hewitt C, Moynihan L, Rob-erts E, Woods CG, Markham A, Wong M, Wid-mer R, Ghaffar KA, Pemberton M, Hussein IR, Temtamy SA, Davies R, Read AP, Sloan P, Dixon MJ, Thakker NS. Loss-of-function mutations in the cathepsin C gene result in periodontal dis-ease and palmoplantar keratosis. Nat Genet 1999; 23: 421-424.
[2] McGrath ME. The lysosomal cysteine proteas-es. Annu Rev Biophys Biomol Struct 1999; 28: 181-204.
[3] Santilman V, Jadot M, Mainferme F. Impor-tance of the propeptide in the biosynthetic maturation of rat cathepsin C. Eur J Cell Biol 2002; 81: 654-663.
[4] Meade JL, de Wyntr EA, Brett P, Sharif SM, Woods CG, Markham AF, Cook GP. A family with papillon-lefèvre syndrome reveals a require-ment for cathepsin C in granzyme B activation and NK cell cytolytic activity. Blood 2006; 107: 3665-3668.
[5] Tran TV, Ellis KA, Kam CM, Hudig D, Power JC. Dipeptidyl peptidase I: importance of progran-zyme activation sequences, other dipeptide sequences,and the N-terminal amino group of synthetic substrates for enzyme activity. Arch Biochem Biophys 2002; 403: 160-170. [6] de Haar SF, Hiemstra PS, van Steenbergen MT,
mcomitans. Infect Immun 2006; 74: 5284-5291.
[7] Tye CE, Pham CT, Simmer JP, Bartlett JD. DPPI may activate KLK4 during enamel formation. J Dent Res 2009; 88: 323-327.
[8] Lu Y, Papagerakis P, Yamakoshi Y, Hu JC, Bartlett JD, Simmer JP. Functions of KLK4 and MMP-20 in dental enamel formation. J Biol Chem 2008; 389: 695-700.
[9] Caterina JJ, Skobe Z, Shi J, Ding Y, Simmer JP, Birkedal-Hansen H, Bartlett JD. Enamelysin
(matrix metalloproteinase 20)-deficient mice
display an amelogenesis imperfecta pheno-type. J Biol Chem 2002; 277: 49598-49604. [10] Hart PS, Hart TC, Michalec MD, Ryu OH,
Sim-mons D, Hong S, Wright JT. Mutation in kalli-krein 4 causes autosomal recessive hypomat-uration amelogenesis imperfecta. J Med Genet 2004; 41: 545-549.
[11] Kim JW, Simmer JP, Hart TC, Hart PS, Ramas-wami MD, Bartlett JD, Hu JC. MMP-20 muta-tion in autosomal recessive pigmented hypo-maturation amelogenesis imperfecta. J Med Genet 2005; 42: 271-275.
[12] Simmer JP, Hu Y, Lertlam R, Yamakoshi Y, Hu JC. Hypomaturation enamel defects in Klk4 knockout/LacZ knockin mice. J Biol Chem 2009; 284: 19110-19121.
[13] Simmer JP, Fukae M, Tanabe T, Yamakoshi Y, Uchida T, Xue J, Margolis HC, Shimizu M,
De-Hart BC, Hu CC, Bartlett JD. Purification, char -acterization, and cloning of enamel matrix serine proteinase 1. J Dent Res 1998; 77: 377-386.
[14] Yoon H, Blaber SI, Li W, Scarisbrick IA, Blaber
M. Activation profiles of human kallikrein-relat -ed peptidases by matrix metalloproteinases. Biol Chem 2013; 394: 137-47.
[15] Ryu O, Hu JC, Yamakoshi Y, Villemain JL, Cao X, Zhang C, Bartlett JD, Simmer JP. Porcine kalli-krein-4 activation, glycosylation, activity, and expression in prokaryotic and eukaryotic hosts. Eur J Oral Sci 2002; 110: 358-365.
[16] Yamakoshi Y, Simmer JP, Bartlett JD, Karakida T, Oida S. MMP20 and KLK4 activation and in-activation interactions in vitro. Arch Oral Biol 2013; 58: 1569-1577.
[17] Overall CM, Limeback H. Identification and
characterization of enamel proteinases isolat-ed from developing enamel: amelogeninolytic serine proteinases are associated with enamel maturation in pig. Biochem 1988; 256: 965-972.
[18] Pavlic A, Petelin M, Battelino T. Phenotype and enamel ultrastructure characteristics in pa-tients with ENAM gene mutations g.13185-13186insAG and 8344delG. Arch Oral Biol 2007; 52: 209-217.
[19] Moradian-Oldak J, Jimenez I, Maltby D, Fin-cham AG. Controlled proteolysis of amelo-genins reveals exposure of both carboxy- and amino-terminal regions. Biopolymers 2001; 58: 606-616.
[20] Zhu L, Liu H, Witkowska HE, Huang Y, Tanimoto K, Li W. Preferential and selective degradation and removal of amelogenin adsorbed on hy-droxyapatites by MMP20 and KLK4 in vitro. Front Physiol 2014; 5: 268.
[21] Bartlett JD, Simmer JP. Kallikrein-related pepti-dase-4 (KLK4): role in enamel formation and revelations from ablated mice. Front Physiol 2014; 5: 240.
[22] Xie X, Liu C, Zhang H, Jani PH, Lu Y, Wang X, Zhang B, Qin C. Abrogation of epithelial BMP2 and BMP4 causes amelogenesis imperfecta by reducing MMP20 and KLK4 expression. Sci Rep 2016; 6: 25364.
[23] Yamakoshi Y, Richardson AS, Nunez SM, Yama-koshi F, Milkovich RN, Hu JC, Bartlett JD, Sim-mer JP. Enamel proteins and proteases in Mmp20 and Klk4 null and double-null mice. Eur J Oral Sci 2011; 19 Suppl 1: 206-216. [24] Smith CE, Richardson AS, Hu Y, Bartlett JD, Hu
JC, Simmer JP. Effect of kallikrein 4 loss on enamel mineralization: comparison with mice lacking matrix metalloproteinase 20. J Biol Chem 2011; 286: 18149-18160.