142
INTRACELLULAR OXIDATION-REDUCTION
STUDIES
V. A COMPARISON OF INTACT AND CYTOLYSED
STARFISH EGGS BY THE IMMERSION METHOD
BY ROBERT CHAMBERS, LYLE V. BECK
AND
DAVID E. GREEN.
(New York University.)
(From the Eli Lilly Research Division, Marine Biological Laboratory, Woods Hole, Mass.)
(Received 4th September, 193a.)
I N a previous communication (o of this series on the permeability of Echinoderm eggs to the oxidation-reduction indicators of the Clark and Cohen series it was observed that none of the sulphonic acid indicators entered the eggs whereas all the others of the series penetrated with ease. Moreover, it was shown that any possible extracellular oxidation-reduction reaction of Echinoderm eggs, or, for that matter, of sea water, is of negligible importance compared with the reaction inside the cells. It followed that the sulphonated indicators can serve their purpose only when the permeability factor is ruled out, viz. in micro-injection studies, and in immersion experiments with cytolysed material.
The purpose of the present investigation was a comparison of the intensity and rate factors, and the effects of chemical agents upon rates of indicator reduction in intact and cytolysed starfish eggs.
It has been stated d) that individual eggs when cytolysed lose their reducing action. This was based on the occasional return, with cytorysis, of the colour of the oxidant in eggs containing the reductant of an injected indicator. The suggestion was made that, upon cytolysis, the substance or substances responsible for the maintenance of a reducing intensity in the cell are lost to the surrounding medium, while oxygen or its equivalent enters the cytolysed structure and induces re-oxidation.
The question still remained open as to whether the cell contents, liberated by osmotic cytolysis, possess the same reducing properties as the intact eggs, or whether the possible destruction of surfaces incident to cytolysis involves a diminu-tion in reducing power.
Iniracelhdar Oxidation-reduction Studies 143
MATERIAL AND METHODS.
The material used for these experiments were the ova of the starfish (Atterias
forbesu). These ova are obtainable in quantities, they can be readily cytolysed, and
when cytolysed are easily distinguished from those which are intact. The eggs are relatively colourless, a pale orange tint being apparent only when they are massed together. The presence of this natural pigment does not interfere with the deter-mination of oxido-reduction colour changes, since all the indicators used, with one exception, are blue as oxidants and colourless when reduced.
The excised ovaries were rinsed in sea water, cut up with scissors, and strained through cheese cloth into sterile sea water. The egg suspension was then divided among several centrifuge tubes and the eggs rapidly sedimented. The supernatant liquid was removed by suction and replaced with fresh sea water. By this procedure as much as possible of the debris from ovarian tissue was separated from the intact eggs.
For anaerobic studies on indicator reduction, the material was exhausted of air in standard Thunberg tubes equipped with a side bulb into which the indicator solution was introduced. The use of this tube made it possible to complete the exhaustion and equalise the temperature before mixing the egg suspension and indicator solution.
EXPERIMENTAL.
(1) INTENSITY FACTOR, INTACT AND CYTOLYSED EGGS.
The indicators chosen for this purpose are listed in Table I. Toluidine blue, Janus green and Cresyl violet, introduced here in the experimental work of this series, have been described in the monograph of Wurmserd).
For aerobic experiments, a series of test-tubes of equal bore were selected and paired. A uniform egg suspension was made up and 2 c.c. placed in each of the pair. Drops of M/iofiOO indicator solution were added to one of the pair and comparison made with the other as control for the extent of reduction. The tubes were constantly shaken to insure aerobic conditions throughout. A further test to detect possible reduction was made by adding one or two drops of 1 per cent, potassium ferricyanide as oxidising agent. This compound rapidly penetrates the eggs. To balance the effect of the yellow colour of the ferricyanide in the colour comparisons an equal quantity was added to the control.
The same method was used in studying the reducing intensity of cytolysed eggs. To produce cytolysis, the mass of sedimented eggs was diluted with a M/15 phosphate buffer solution at pH 7-0, and the mixture shaken vigorously for several minutes. The material was used only when microscopic examination showed that cytolysis was complete. A quinhydrone electrode determination gave for the cytolysed egg buffer mixture the same pH as for the buffer solution alone, ijt. 7-0, since the acidity produced by cytolysis was not sufficient to change the pH of the buffer solution.
Tabl e I . Indicator s •C m-bromopheno l indopheno l D o-chloropheno l indopheno l K o-creao l indopheno l L o-creso l indo-2 , 6-dichloro -pheno l O /-naphthoI-2-*ulphonat e indopheno l D l-naphthol-a-culphonate -3 , 6-dichloropheno l Q Tohiylen e blu e chlorid e Q , Creay l bhi e chlorid e Q , Tohndin e blu e R Metfayien e blu e S , Ethy l Capr i blu e T Indig o trwulphonat e U Indig o disulphonat e U , Cresy l viole t W Janu s green , li t ste p „ an d ste p X Phenosafrani n £j'a t ptiro +0-24 8 + 0-23 3 + 0-19 5 + 0-18 1 + 0-12 3 + 0-11 9 + 0-11 5 + 0-04 0 + o-on j + O-OI I - 0-06 0 -0-08 1 — 0-12 5 -0-167 ! ? -0-258 ! — 0-24 2 r H 22 3 21 8 30 5 2O-I l8 l l8-O 17-9 15 5 14 4 14 4 I2-O n-3 9 9 8-3
? 52 —
Reductio n b y starfis h egg s Livin g egg s Aerobi c Reduce d
tt tt $t
Notr*ductd\ I t Reduce d I
f tt tt
Partiall y reduce d Notrtductd » No t reduce d
•» >t it
Anaerobi
c
Reduce
d
tt t> it
Notrtdtued
I
I
Reduce
d
tt tt • •
Not rmhictd ft Reduce d »» No t reduced ? No t reduce d Cytolyve d egg t Aerobi c Reduce d
it tt tt ti *> it tt tt 11
Partiall y reduce d No t reduce d
tt tt tt
Anaerobi
c
Reduce
d
•
1 tt ti
» tt i* tt » tt tt tt tt tt
No t reduced ? No t reduce d Remark s Penetrate * livin g egg s
tt tt tt
Sulphonated , doe s no t penetrat e livin g egg s *» Penetrate s livin g egg i
tt tt »» »»
Sulphonated , doe s no t penetrat e livin g egg s i i Penetrate s livin g egg s
tt tt tt
Th e letterin g i s th e sam e a s tha t use d i n th e previou s paper s o f th e series . Statement s italicise d refe r t o non-reductio n becaus e o f non-penetration . E,' value * take n fro m Wurmser(a) . Th e value s no t marke d ar e thos e take n fro m th e previou s paper s o f th e series .
IntraceUular Oxidation-reduction Studies 145
Exhaustion was accomplished with a water pump, after which the indicator solution was mixed with the suspension by tilting the tube. Shaking the tube accelerated the process of reduction and, when decoloration took place, the reaction was com-plete within 5-10 min. To induce reoxidation the stopcock was then opened and the tube again shaken to insure complete aeration.
The results are presented in Table I. It will be noted that the sulphonated indicators in the concentrations used are not detectably reduced in the presence of intact eggs under either aerobic or anaerobic conditions. On the other hand, they are reduced in the presence of cytolysed eggs.
The aerobic apparent reduction potential for both intact and cytolysed eggs was found to be the same, lying within the range of ethyl Capri blue (—0*06 volt). This indicator was only partially reduced, since, with minimal detectable amounts, complete decoloration was never observed. Basing our results from the evidence secured by use of Janus green and Cresyl violet, the anaerobic oxidation-reduction potential may be considered to lie between the limits of —1-6 and —2-43 volte. The anaerobic value for both intact and cytolysed eggs appears to be identical.
The reaction with Janus green requires special mention. It was found that a c.c. of an egg suspension containing o-8 c.c. of egg sediment in an exhausted Thunberg tube will reduce 0-35 c.c. of Af/10,000 Janus green in 20 min. The colour of the suspension changed from greenish blue of the oxidant through purple to deep red of the reductant. After standing, the supernatant liquid was seen to be purplish blue, while the sedimented eggs were distinctly red in colour. Re-admission of air to the tube resulted throughout in a gradual return within several minutes of the original, greenish blue colour of the oxidant. The slight colour change in the supernatant fluid is doubtless due to the presence of some cytolytic products, since a shift to red of the supernatant fluid is much more pronounced when the egg suspension is completely cytolysed prior to exhaustion. The second step in the reduction from red to colourless was not observed during the first few hours. When the eggs were allowed to stand for 5-6 hours, complete decoloration did take place. However, it is possible that this was due to bacterial action. Rapkine, Struyk and Wurmserb), using the potentiometric method, record the first step in the reduction of Janus green to be practically irreversible. Apparently the condi-tions necessary for reversibility are present in the starfish eggs used by us.
(a) RATE FACTORS FOR INTACT AND CYTOLYSED STARFISH EGGS
UNDER ANAEROBIC CONDITION.
resus-146 R. CHAMBERS, L. V. BECK and D. £. GREEN
pended in an equal volume of sea water. The two portions of intact and cytolysed egg suspensions were then divided into 2 c.c. lots in Thunberg tubes each of which contained in the side arm 0-25 c.c. of M/10,000 solution of an indicator. The eight tubes were then simultaneously exhausted of air and a record taken of the time required for complete reduction in each tube.
Table II.
Indicator
o-cresol indophenol Brilliant Cretyl blue Metfaylene blue Ethyl Capri blue
Reduction potential at pH 7-0
0-195 0-04 +O-OII —0-06
Time to secure complete anaerobic reduction
Intact egg»
< i aec. 13 min. " » 56 „
Cytolysed egg*
< i sec. 61 min. 118 „ 180 „
Table III.
Different batches of eggs
1 3
3 4 5
Time for a ex. egg suspension to reduce oas ex. Jtf/io,ooo methylene blue (min.)
Intact eggs
ia
10
5
ia
10
Cytolysed eggs
6a 74
i
1
65
The results given in Table II are taken from one typical experiment. With different batches of eggs there was a considerable variation in the actual times for reduction but all the experiments show definitely that the higher an indicator lies in the oxidation reduction scale, the more rapidly is it reduced by both intact and cytolysed eggs. The variability of different lots of eggs for methylene blue re-duction is shown in Table III. The ratio (cytolysed/intact) in terms of time to secure complete reduction varied from 3 to 8.
(3) EFFECTS OF CHEMICAL AGENTS ON RATES OF REDUCTION
UNDER ANAEROBIC CONDITIONS.
The chemical agents used were ether, phenyl urethane, ethyl alcohol, potassium cyanide, lead carbonate and mercuric oxide.
For experiments with intact eggs the chemicals were dissolved in sea water and where necessary the solutions were brought to the pH of sea water by electro-metric titration with M/10 HC1 and NaOH. For experiments with cytolysed eggs the chemicals were dissolved in M/15 phosphate buffer at pH 7-0, and when necessary the solutions were brought back to this pH by electrometric titration.
Intracelhdar Oxidation-reduction Studies
147
Saturated stock solutions of ether, phenyl urethane, lead carbonate and mercuric oxide were prepared in sea water and in Af/15 phosphate buffer. The solubility constants of these compounds in sea water and in the phosphate buffer were assumed to be approximately the same as for distilled water.Various dilutions of a given stock solution were prepared with egg suspensions in a series of Thunberg tubes and care was taken to have the same amount of egg suspension in each tube. The highest concentrations selected were those which did not induce cytolysis within several hours of immersion. A control tube of egg suspension with no experimental reagent was also set up. The tubes were simultaneously exhausted and brought to the same temperature in a bath of running sea water. The indicator solutions and egg suspensions were then rapidly mixed and the time required to secure complete reduction in each tube was determined. For cytolysed material the procedure was the same except that the buffer solution was substituted for sea water.
(a) Narcotics.
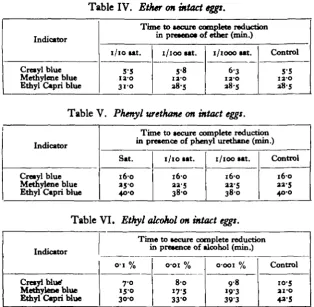
[image:6.451.68.383.326.633.2]Tables IV, V and VI present the results of the effects of varying concentrations of ether, phenyl urethane and ethyl alcohol on the anaerobic reducing power of intact eggs.
Table IV. Ether on intact eggs.
Indicator
Cre»yl blue Methylcne blue Ethyl Capri blue
Time to secure complete reduction in prcMoc* of ether (min.)
1/10 tat.
5-5 ia-o 310
1/100 tat.
5-8 ta-o *8-5
1/1000 tat.
63 ia-o a8S
Control
5-5 ia-o a8-5
Table V. Phenyl urethane on intact eggs.
Indicator
Cretyl blue Methylene blue Ethyl Capri blue
Time to secure complete reduction in pretence of phenyl urethane (min.)
Sat.
16-0 350 40-0
1/10 tat.
16-0 aa-5 380
1/100 tat.
16-0 aa-5 380
Control
16-0 aa-5 40-0
Table VI. Ethyl alcohol on intact eggs.
Indicator
Cretyl blue1 Methylene blue Ethyl Capri blue
Time to tecure complete reduction in pretence of alcohol (min.)
o-i % 7-0 150 30-0
o-oi %
8-o 175 330
o-ooi %
98 193 393
Control
148 R. CHAMBERS, L. V. BECK and D. £. GREEN
By comparison with the controls in the last column of the tables, it will be seen that ether and phenyl urethane have no appreciable effect on the reduction. Ethyl alcohol exerts a definite accelerating influence.
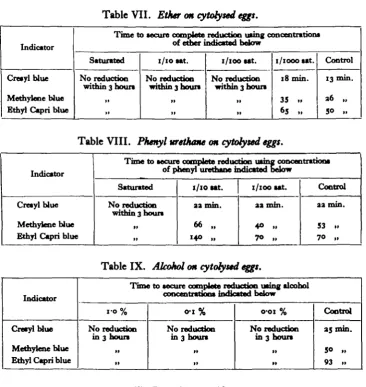
[image:7.451.44.411.184.571.2]Tables VII, VIII and IX present the effects on cytorysed eggs. It is seen that concentrations of ether and alcohol which had no inhibitory effect on reductions of intact eggs, destroy the reducing power of cytorysed eggs. A similar effect of phenyl urethane occurred only in the highest concentration, probably owing to the relative insolubility of phenyl urethane in water.
Table VII. Ether on cytofysed eggs.
Indicator
Cresylblue
Methyiene blue Ethyl Capri blue
Time to secure oomplrtn reduction using concentration* of ether indicated below
Saturated
No reduction within 3 hour*
11 11
1/10 sat.
No reduction within 3 hours
1 1 1 1
1/100 sat.
No reduction within 3 noun
1 1 11
I/IOCO sat.
18 min.
35 „ 65 .,
Control
13 min
36 „ 50 ..
Table VIII. Phenyl urethane on cytofysed eggs.
Indicator
Time to secure complete reduction using concentration* of phenyl urethane mdicatedT '
Saturated 1/10 sat. 1/100 sat. Control Cresylblue
Methyiene blue Ethyl Capri blue
No reduction
within 3 hour* aa min. 66 „
aa min.
40 »
aa nun.
53 .. 70 ,.
Table IX. Alcohol on cytofysed eggs.
Indicator
Tim© to secure p gifrthictioffi ohol trations indicated below
0-1% 0-01% Control
Cresylblue
Methyiene blue Ethyl Capri blue
No reduction
in 3 hours No reductionin 3 hours No reductionin 3 hours 35 min. 50 ,, 93 »
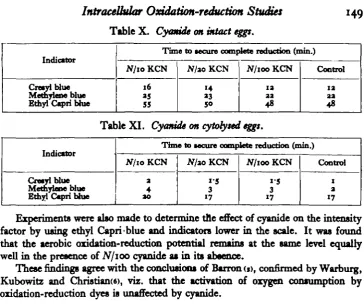
(b) Potassium cyanide.
IntraceUular Oxidation-reduction Studies
Table X. Cyanide on intact eggs.
149
Indicator
Cresyibtue Metbylene blue Ethyl Capri blue
Time to secure complete redaction (min.)
N/io KCN
16
55
N/zo KCN
14
»3
50
N/100 KCN
ia aa 48
Control
1a aa 48
Table XI. Cyanide on cytofyted eggt.
Indicator
Cretylblue Methyleoe blue Ethyl Capri blue
Time to secure complete reduction (min.)
N/10 KCN
a 4 ao
tf/aoKCN
i-5 3 17
N/ioo KCN
i-5 3 17
Control
1 a 17
Experiments were also made to determine tne effect of cyanide on the intensity factor by using ethyl Capri blue and indicators lower in the scale. It was found that the aerobic oxidation-reduction potential remains at the same level equally well in the presence of N/100 cyanide as in its absence.
These finding" agree with the conclusions of Barron <i>, confirmed by Warburg, Kubowitz and Christianw, viz. that the activation of oxygen consumption by oxidation-reduction dyes is unaffected by cyanide.
(c) Mercury and lead.
[image:8.451.46.411.391.633.2]The results with intact eggs presented in Tables XII and XIII show that mer-curic oxide in exceedingly small concentrations destroys the reducing mechanism, while lead carbonate in comparable concentrations is not as effective.
Table XII. Mercuric oxide on intact eggs.
Indicator
Time to secure complete reduction
2U X IO
M/lOO C.C.
a-A x io~*
M/ioo c.c M/100 ex. Control
Crcayl blue
Methyleoe blue Ethyl Capri blue
No reduction within 3 noun
No reduction
within 3 noun No reductionwithin 3 noun 13 mm. 34 » 60 .,
Table XIII. Lead carbonate on intact eggs.
Indicator
Cretylblue MetfayUaeblue Kttrrl Cuvihliu
Time to secure complete reduction (min.)
7 4 x 10-*
JM/IOOCX.
39 ao 150
TA X IO-* M/ioo ex.
*s
17 130
7-4 x io~*
M/ioo c.c.
18 14 130
Control
150 R. CHAMBERS, L. V. BECK and D. E. G R E E N
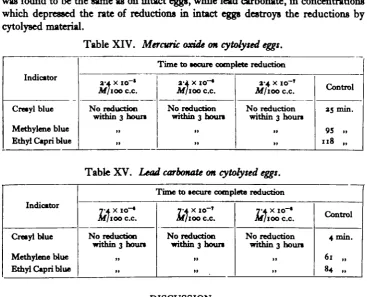
[image:9.451.43.411.99.396.2]For cytolyscd eggs, sec Tables XTV and XV, the effect of the mercuric oxide was found to be the same as on intact eggs, while lead carbonate, in concentrations which depressed the rate of reductions in intact eggs destroys the reductions by cytolysed material.
Table XIV. Mercuric oxide on cytolysed eggs.
Indicator
Cresyl blue
Methytene blue Ethyl Capri blue
Time to secure complete reduction
I ' l X IO~* Af/100 c.c.
N o reduction within 3 hours
>> >>
a-i. x io~*
M/lOO C.C.
N o reduction within 3 hours
»> >»
a-4 x io~T
M/ioo c.c.
No reduction within 3 hours
it >>
Control
35 min.
95 » 118 „
Table XV. Lead carbonate on cytolysed eggs.
Indicator
Creaylblue
Methylene blue Ethyl Capri blue
Time to secure complete reduction
T4 X 10-*
M/100 c.c.
No reduction within 3 hours
If II
T4 X IO-'
M/ioo c.c
No reduction within 3 hours
11 11
7-A X IO~*
Jw/ico c.c.
No reduction within 3 hours
11 11
Control
4 min.
61 „ 84 ..
DISCUSSION.
There is a marked tendency of penetrating indicators to accumulate preferen-tially in the eggs. If, under aerobic conditions, an amount of indicator is present in excess of that which can be reduced the eggs soon become deeply coloured. If the eggs are then placed under anaerobic conditions, the colour disappears, provided the indicator is reducible. On readmission of air, the colour returns to the eggs but no colour is seen in the supernatant liquid. This is true for o-cresol indophenol as well as for the basic dyes lower down in the scale. On the other hand, when solutions of the sulphonated indicators are added to intact eggs, and the eggs are allowed to settle, the supernatant liquid always remains coloured, whereas the sediment shows no trace of being coloured with the dye. Anaerobioais does not change the situation. The inference is that these sulphonated indicators are not reduced because they do not penetrate.
Nevertheless, the proof that such sulphonated indicators are not at all reduced must await direct chemical or electrometric measurement. The reason for this is apparent when one considers that small amounts of reduction may perhaps have escaped detection in the simple colorimetric method here employed.
Intracelhtlar Oxidation-reduction Studies 151
Barron(5) has pointed out recently that though there is no necessary relation between the speed and free energy of a reaction, cases have been recorded in biological systems where such a relation holds. For example, they found that the catalytic effect of oxidation reduction indicators on oxygen consumption varies directly with the position of the indicator on the oxidation-reduction potential scale. Also Voegtlin, Johnson and Dyer (7), working with extracts of noraud and cancerous tissue, found that the time required to secure reduction of an indicator was approximately a logarithmic function of its electrode potential.
Cytolysis has a marked retarding effect on the rate of reduction. It is possible that osmotic cytolysis destroys surfaces which are involved in the reduction of indicators; it is alto possible that the preferential accumulation of the indicators in living intact cells makes for higher effective concentrations than would be present in cytolysed egg suspensions.
Ether, phenyl urethane and alcohol which exert a profound inhibitory effect on the reducing power of cytolysed egg material have no such action on intact eggs. It is generally assumed that dehydrogenases are' concerned in the reduction of oxidation-reduction indicators, and that narcotics preferentially inhibit the action of these enzymes. It would have seemed likely, therefore, that narcotics should inhibit the reduction of oxidation-reduction indicators. Success in securing marked inhibition by ether, alcohol and phenyl urethane on cytolysed material, but not on intact material, suggests that while the cell is alive the narcotic does not reach the postulated dehydrogenase.
The difference in action of lead carbonate on cytolysed as compared with that on intact material is also of considerable interest. Indeed, a comparison of the results secured by the use of cytolysed with those obtained by the use of intact eggs, indicates that no accurate prediction can be made from the former of what would be obtained by experiments with the latter.
Micro-injection in conjunction with immersion methods would seem to be adequate for the study of most cellular oxidation-reduction phenomena; they deal with the living cell, are relatively non-injurious, and actually take less time than do extraction methods.
SUMMARY.
1. The aerobic, apparent, oxidation-reduction potential for both intact and cytolysed starfish eggs at pH 6-8-7-O is about — 0-06 volt, the anaerobic is below —0*167 vo^"
a. A reversibility of the first step in the reduction of Janus green by starfish eggs was observed, viz. a colour change from blue to red, and, by oxidation, a return to blue.
3. The higher an indicator lies, in the oxidation-reduction scale the more rapidly is it reduced by both intact and cytolysed eggs.
4. Cytolysis results in a lowered rate of reduction.
152 R. CHAMBBRS, L. V. BECK and D. E. GREEN
speed up the rate. However, alcohol and ether completely destroy the reducing power of cytolysed eggs; phenyl urethane, which is relatively insoluble, acts similarly though not to as marked a degree.
6. KCN has no effect either on the intensity or rate factor of intact or cyto-lysed eggs.
7. Mercuric oxide, one part in a million, destroys the reducing power of both intact and cytolysed eggs. Lead carbonate, in a similar concentration, destroys the reducing power of cytolysed eggs but only retards that of intact eggs.
REFERENCES.
(1) CHAMBDS, R., COHIN, B. and POLLACK, H. (1931). Joint. Exp. Biol. 8, 1.
(a) W u n n n , R. (1930). Oxydatiom t rdittetkm. Paria.
(3) RATON*, L., STKUYK, A. P. and W o u a n , R. (1939). Jam*. Chtm. Phyt. 36, 340. (4) CHAiaaa, R. and POLLACK, H. (1937). Jam. Omu Pkytiol. 10, 739.
(5) BARRON, E. S. G. (1939). Jtmm. Bid. Chtm. 81, 445.
(6) WARBUBO, O., KUBOWTTZ, F. and CHRISTIAN, W. (1930). BtocA. Zatschr. 227, 345.
(7) VOBOTLIN, C , JOHNSON, J. M. and D m , H. A. (1935). Jour*. Pharm. and Exp. Thtrap. 24, 305.
ADDENDUM.