0032-0889/88/87/0239/05/$01.00/0
Abundance of
theMajor Chloroplast
Polypeptides during Development andRipening
of Tomato FruitsAN IMMUNOLOGICAL STUDY
ReceivedforpublicationAugust 17, 1987 and in revised form December 1, 1987
ALEXANDER LIVNE AND SHIMON GEPSTEIN*
Department ofBiology, Technion-lsraelInstitute ofTechnology, Haifa 32000, Israel
ABSTRACT
During maturation and ripening of tomato (Lycopersicon esculentum, cvTamar) fruits, there aredifferentialchangesinthesteadystatelevels ofchloroplast proteins. Western blot analysis indicated that with the exception of the corepolypeptideofphotosystemI(PSI)(subunit I) the wholecomplex disappears during thetransition of chloroplast to chrom- oplast.Theamountsof thecorepolypeptideofphotosystemII(PSII) (43 kilodaltons)andthelight harvesting chlorophyllprotein complexincrease duringmaturation and decreasethereafter.Incontrast,the 33 kilodalton subunit ofPSII is foundatthe highest levels from theearly recorded stages and decreases gradually untillate stages ofripening.The level of cytochromefdecreasesslowlyduring the maturation andripeningprocess, whereas theRieske proteinofthe samecomplexdisappears at afaster rate.There are alsodifferential changesin the subunits of thechloroplast coupling factor ATPasecomplex;aandfisubunitsincreaseduring ma- turation, whereas the level of the y subunit is alreadymaximal at the earliestrecorded stage ofdevelopmentanddepleted thereafter. The two subunits of theribulose-1,5 bisphosphatecarboxylase increase in abun- danceduring chloroplast maturation andgradually disappear after the transitionfromchloroplasttochromoplast. However,there are substantial differences in the ratesof increase and disappearanceofthelarge and smallsubunits of this enzyme. This imbalance is attributedtodifferent regulation of nuclear andchloroplast gene expression. In addition, the steady state levels ofchloroplastic superoxide dismutase and phospho- enolpyruvatecarboxylasehave beenfollowed. Both enzymes reach their maximaatthefinalstagesofripening. This increasecoincides with the climacteric riseofCO2release.
ing membrane disintegration, Chl breakdown, and starch dis- appearance (1,8, 16). The content of carotenoids increases and lycopene is synthesized in thecytoplasm and accumulated within innermembranesduring the transformation from chloroplaststo chromoplasts (4, 7, 13, 14). This transformation takes place at the samedevelopmental stage that other ripening processes such ascellwalldegradationandsoftening and accumulation ofsugars andorganic acids areoccurring (12). Thetransformation of chlo- roplasts, as well as the whole ripening process, is not a deteri- orative and aging process per se. Rather, it is characterized by aseries ofeventswhichinclude de novo synthesis and activation ofspecific enzymes which are significant for the normal ripening process. Activeprotein synthesis during ripening has been dem- onstrated by Spiresetal. (30) and includes the synthesis ofin- vertase (12),polygalacturonase (6, 12,28), pectin methyl ester- ase(12, 28), and other enzymes. Comparisonofprotein profiles ofchloroplasts and chromoplasts following separation on SDS- PAGE revealed manydifferencesboth quantitatively and qual- itatively (12, 30). During the transition from chloroplast to chromoplast,there isadecreasein manypolypeptidesaswellas a substantial increase in specific polypeptides (6, 12, 24). La- belingofproteinsinvivowith[35S]methioninehas demonstrated differences intherate atwhichspecific proteins are synthesized (1). Little is known regarding theidentity of the chloroplastic polypeptides that undergo changes during the development of fruits and the transition of chloroplasts to chromoplasts. The present study has focused on molecular events accompanying development and thetransformation of chloroplasts to chrom- oplasts. In particular, we have used immunological techniques tofollowselectivechangesinseveral of theprincipalpolypeptide componentsofthephotosynthetic apparatus.
Mature green tomato fruits are capable of assimilating CO2 photosynthetically, mainly in the external pericarp where the highestphotosyntheticactivityhas been recorded(2,11, 17,34).
Thephotosyntheticassimilation of thesefruitsindicates the pres- enceof functional components of the photosyntheticapparatus and suggests apossible significance of this process for fruits. It has been suggested that the CO2 evolvedduring respiration in fruits is refixed andrecycled through the process of photosyn- thesis (2,33,34). Green tomato fruits containchloroplaststhat show the typical ultrastructural features of leaf chloroplasts, namely, grana, which contain the components involved in the light reactions, and the stromawhere the Calvincycle enzymes arelocalized (1). Thoughthere is considerable information re- garding the composition, function, and structural organization of the main polypeptide complexes in leaf thylakoids, little is known about the structure, stoichiometry, and organization of tomatofruitchloroplasts. Duringmaturation and fruitripening, thechloroplastsundergogradual ultrastructuralchangesinclud-
MATERIALS AND METHODS
PreparationofProteinSamples. Tomato (Lycopersicon escu- lentum, cvTamar) fruits atvarious stages ofdevelopmentwere harvested in the field.Eightdevelopmentalstages wereclassified according to fruit color and size: (a) green, 10 to 15 mm (di- ameter); (b) green, 15to30mm;(c) green, fullyexpanded (ma- ture); (d)green yellowish; (e)yellow; (f)orange; (g) orangered;
and (h) dark red and soft. Sampling was confined to theouter layer(2 mm) of thepericarp since it containsa high density of chloroplastsand thehighest photosyntheticactivity of all tissues inthe fruit(11, 17,34). Approximately 300 mg of the tissue were homogenizedat 4°C in 7 ml of100 mm Tris (pH 8.0). The ho- mogenate was filtered through four layers of cheesecloth, and 0.15 ml of a solution containing 1.85 M NaOH and 7.4% 2- mercaptoethanol were added for each ml of filtrate. After 10 min at 4°C, 150 gl of 50% TCA were added for each ml of solution, and the solutionwas held for an additional 10 min at 239
PlantPhysiol. Vol. 87, 1988 4°C before centrifuging at 17,000 rpm (Beckman-Rotor 20) for
10min at 4°C. Thepelletwassuspended in 10 ml of 80% acetone andrecentrifuged.The acetone wasremoved and thepellet dried withnitrogen and thensolubilized in 2% SDS. Allsampleswere storedat -20°C before electrophoretic separation.
Electrophoresisand Immunoblotting. Since the percentage of totalproteinperfreshweight was constant throughout the stages of fruit development (data not shown), the samples were cal- culatedonfresh weightbasis. Proteinsampleswere loaded and electrophoresedon 12.5% SDS-polyacrylamide gelsusing a La- emmli discontinuoussystem.The electrotransferofproteinswas followed by immunodetectionwith specific antibodies asprevi- ously described (3, 9, 31). Quantitationofsteady state levels of specific polypeptideswascarried outwith Beckman DU-8spec- trophotometer. Six biological replicates were carried out, and representative experiments aredepictedhere.
Isolation of the PSI Complex and Preparation of Antibodies.
The PSI complex was isolated from leaves of Swiss chard fol- lowing themethod of Nechushtai etal. (19, 20). The main sub- unitswereseparatedonSDS-PAGE andwereinjectedintorab- bitsaspreviously described(20). Antibodiesagainst subunitsof thefollowingchloroplastpolypeptideswerepreparedin our lab- oratory: RuBPcasel (35), Cytb6Ifasdescribed previously (15), ATPase (20-22), and the PSII complexasdescribedpreviously (5,18).Thespecificity oftheantibodieswasconfirmed bycom- parison oftheirreactivity in crude tomato extractswith that of antibodies kindlydonated byDr.N. Nelson(18-21). Antibodies against the chloroplastSOD and PEPcasewerepreparedaspre- viously described (29,32).
RESULTS
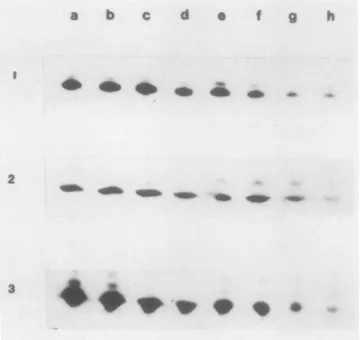
Figure 1depicts the changesintheamountsofthethree sub- units of the CF1 complex during development and ripening of tomatofruits. The a-and,-subunitsincreasedduring theearly stagesofmaturation, reached theirhighest levelsatstage c(ma- turegreen), and thendecreasedasripening andsenescence pro- gressed(Table I). In contrast,theamountof the y-subunitwas at its highest level during the earliest recorded stage ofdevel- opment (a)anddecreased gradually thereafter. Asimilar trend of disappearance of the three CF1 subunits has been demon- strated. Duringthefinalstagesofsenescence,detectable amounts of the polypeptideswerestill present(Fig. 1) indicatingincom- plete destruction of this complex.
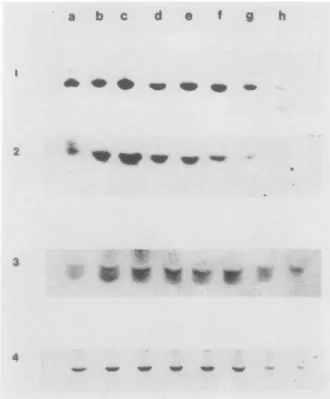
Changesin the fivesubunits of the PSIcomplexare shownin Figure2.The'core' of thePSI (subunitI, 66-70kD)wasstable throughout development and ripening,butamounts of subunits II, III, VI and VII increased until stage c (maturegreen) and thendecreasedduring the remainder of the ripeningprocess(Fig.
2). With theexception of subunit I, the complex of PSI disap- peared during the transition of chloroplaststochromoplast(stages dande).
Dataon the levels ofCytb6IfareincludedinFigure3 (1, 2).
Cytf(34kD)wasrelativelystablethroughout thematuration and ripeningprocess anddisappeared onlyattheterminal stages of senescence. Incontrast, the Rieskeprotein (subunit III of this complex) disappeared, and at stage f the polypeptide was not detected.
The PSII isrepresented inFigure 3bythreesubunits: the 43 kD core polypeptide, the 33 kD polypeptide, and the LHCP complex. The 43 kD polypeptides and the LHCP complex are both associated with Chl; both subunits increased significantly (1.5-to2.5-fold)during maturation of thetomatofruit (stagea
IAbbreviations: RuBPcase, ribulose-1,5 bisphosphate carboxylase;
LSU, large subunit; SSU, smallsubunit;CF,, chloroplast coupling factor;
SOD, superoxide dismutase; PEPcase, phosphoenolpyruvate carboxy- lase;LHCP, light harvesting Chlprotein.
a b c d e f g h
2
*, ,
FIG. 1. Changes in the steady state amounts of theCF,-ATPasecom- plex duringdevelopment and ripening of tomato fruit as visualized by Western blot analysis. Lanes a to h are the developmental stages as described in "Materials and Methods," a to d are stages in fruit expan- sion, and d through h are stages inripening; 1, 2, and 3 are the a,f3,
and ysubunits,respectively. Samples equivalent to 10 mg of fresh weight were subjected to SDS-PAGE and blot transferred to nitrocellulose as described under "Material and Methods."
to c), but both were degraded during subsequent ripening and senescence. Incontrast, the 33 kD subunit was at its highest level from the earliest stages of fruit development (a), and its level remained relatively constant until stage e. All three peptides of the PSII complex completely disappeared during the ripening stage.
Thesteady state levels of both the large and the small subunits ofRuBPcase increased from stage a to c (Fig. 4). However, the small subunit increased at a much faster rate, by up to threefold, whereas thelargesubunit increasedby only 150% over the same period (Table I). There was a gradual loss in the amounts of both subunits starting in stage c, but the large subunit still de- tectable in the final stages of senescence whereas the small sub- unit had completelydisappeared (Fig. 4).
Immunoblottingwithantibodies raised againstchloroplastSOD revealed about a threefold increase in the enzyme during de- velopment and maturation (from stage a-c) and about a fourfold increase during ripening andsenescence,after thetransitionfrom chloroplaststochromoplast(Figs. 3 and 4; TableI). Surprisingly, high levels of SOD remained in the tissue during the late stages ofsenescence. The amount of PEPcase in the pericarp, as de- termined byWesternblotting, was stable during the 'green' stages of fruit development, but the level increased during the later stages and reached its maximum concentrations late in the rip- ening process (Fig. 4).
DISCUSSION
Immunologicalstudies offer an effective approach for the study of thestructure andorganizationofthephotosyntheticapparatus in fruits. The use ofquantitative immunolabelingenabled us to follow the sequential changes of the mainchloroplasticpolypep- tides during the development of tomato fruits, in particular dur- ing the transition from chloroplast tochromoplast.
240 LIVNE AND GEPSTEIN
Table I. Changes in the AmountsofVarious Polypeptidesduring Developmentand Ripeningof Tomato Fruit
The amounts areexpressedas apercentage of theamountof thepolypeptideatstagea. Quantitationwascarried out as outlined in "Materials and Methods."
Stage Polypeptide
a b c d e f g h
CF1
a 100 110 132 117 113 119 10 9
f3 100 115 118 94 69 72 23 3
y 100 96 73 52 61 42 16 8
bdlf
I 100 85 96 80 53 44 26 26
III 100 115 117 24 23
PSII
33 kD 100 71 103 75 56 8
43 kD 100 132 157 95 26
LHCP 100 205 263 49 10
PSI
I 100 75 78 121 128 106 109 58
II 100 140 143 60 12
III 100 125 148 14
VI 100 121 191
VII 100 98 151 45 22
RuBPcase
I 100 114 158 85 123 90 51 9
II 100 252 305 207 194 57 10
PEPcase 100 96 91 96 113 139 53 35
SOD 100 109 284 228 164 432 247 107
a b c d e f g h
a b c d e f g h
A, _ A _ _
2 '4 _
* . . ,,
_ _ 4b _ -
'0e*
FIG. 2. Changesin theamountsoffive subunits ofthe PSIcomplex during developmental andripeningstagesoftomatofruitsasvisualized byWestern blotting. Lanesa through h are asindicated inFigure 1.
Panels 1to5 indicateblots reacted with different antibodiesasfollows:
1,68 kDpolypeptideofPSIcomplex (10mgfreshweight) 2,subunitII (15 mgfreshweight); 3,subunit III(15mgfreshweight); 4,subunit VI (15 mgfreshweight); 5,subunit VII(15mgfreshweight).
FIG. 3. Steadystate levels ofthylakoid proteins in tomatofruitsat various stages ofdevelopment and ripening (lanes a-h). 1, Cyt f; 2, Rieskeprotein; 3,43 kDsubunitofPSII; 4,33 kD subunit ofPSII; 5, LHCP of PSII.
2
3
*w im q
4
3
5
4
5
PlantPhysiol. Vol. 87, 1988
a b c d e f 9 h
-b4 qw 40 4
2
3
.4
4
IV IW VW -A
.im ....:
.W V- W. wi
W W W W %
FIG. 4. Changes in the immunologically detectable amounts of RuBPcase, SOD,and PEPcaseduring developmentandripeningofto- matofruits. Thestages(lanes a-h)areindicated inFigure 1.1,LSU of RuBPcase; 2,SSU ofRuBPcase; 3, SOD; 4,PEPcase.
Most ofthe chloroplastic polypeptide complexeshavebi-gen- omic origin; namely, they include polypeptides coded byboth chloroplastand nuclear DNA. Of the subunits studied,the fol- lowingaresynthesizedwithin thechloroplastof 70S ribosomes:
P700reactioncenterproteinofPSI,43 kD subunit ofPSII, LSU ofRuBPcase, a andthe,8subunits ofCF1. The present results indicate that all of these are already present in high levels at early stages of development in L. esculentum and thereafter undergoagradualdeclineduringtheripeningprocess.
The results summarize the changes of the levels of the Ru- BPcase andarenoteworthyinthecontextthatSSU mRNA levels have beenreportedtoincreaseatafasterrate attheearlystages of development and to disappear more rapidly than the LSU mRNAatadvancingstagesofripening (22, 23). This correlation between the steady state levels of mRNA andthe immunolog- icallydetectableamountsofeach of the subunits of the RuBPcase suggeststhat, atleastatthe first stages offruitdevelopment and atlatestages ofripening,geneexpressioncontrols the levels and activityofRuBPcase. The differences and the imbalance between theamountsoftheLSU and SSU mRNAs have been attributed todifferentregulationofnuclear andchloroplastgeneexpression inthepericarptissue(22, 23). Thus,geneexpressionoftheSSU
maylimittherateofsynthesisand assemblyoftheholoenzyme during development and at laterstages during degeneration of thechloroplast.
Thepotential for photosyntheticelectron transport generally declines during leaf senescence, and a similar trend has been observed in the amounts of thylakoid protein present (3, 25).
The observation thatmature tomatofruitpericarpexhibitsmax-
imal photosynthetic activity (27) is consistent with our obser- vation thatatthis stage theimmunologically detectable thylakoid polypeptides reach thehighestlevel and declinethereafter.
The 68 kD polypeptide of PSI (SUI) wasfound to be atthe highest level already at early stages ofdevelopment andstayed
constantthroughout the developmental and ripeningprocesses.
Similarly, in etiolated leaves this P700 polypeptide is already presentinproplastids, whereas all the othersubunitsofthe PSI complexappear at later stages (19, 20).
The changes of LHCP correlate verywell with the temporal patternof the decline of LHCPmRNAduringtomatofruitrip- ening (23). Thus, the fast disappearance of the LHCP at this stage canbe attributed to aninactivation of the gene encoding for thisprotein.
This datarevealdifferences in thestability of thevarioussub- units of theCytb6/fcomplex. Cytfwasrelatively stable,whereas the Rieskeprotein disappeared duringthetransitionfrom chlo- roplast tochromoplast. In contrast tothisprocess, during leaf senescence, Cyt fwasfoundtodecreaseatearlystages and was suggestedto constitutethe rate-limiting factor of the photosyn- thesis (3, 25).
Theparallel disappearanceofChl, PSI, PSII, andb6/fcomplex indicates that during the conversion of chloroplast to chromo- plast, the capability for light harvesting, photochemical reac- tions, andelectron transport islost, andnormal photosynthesis doesnottake place.
Doubts have beenraised regarding theexistence of net pho- tosynthetic CO2fixation in tomato fruits (34). Ourresults indi- cate the presence of RuBPcase even at very late stages ofrip- ening.However,it seemsunlikelythat the Calvin cycleis operable at this stage, since the maincomponents of the light reactions aredestroyedatearlierstages, andthereis nosupply of the light reaction products (ATP, NADPH) which are required for the dark reactions ofphotosynthesis.
Ithas beenproposed that during the ripeningprocess, thehigh activity of PEPcase enables thecarboxylation and recycling of therespiratory CO2(2,33,34). Thepresentstudy has provided evidence that the amount of PEPcase isalreadypresent athigh levels at early stages ofdevelopment but increases and shows two peaks, one during chloroplast maturation and the second onefollowingthetransitiontochromoplast. The increase inPEP- case activity duringtomato fruitripening (34) may well bedue to a parallel increasein the amount oftheimmunologicallyde- tectable protein as shown in the present study. Of particular interest is the observation that the increase in the amount of PEPcase anditscarboxylation capabilitycoincides with the cli- mactericrise(stageseandf).Itwillbe temptingtosuggestthat the increase in PEPcase ispossibly associated with the require- mentofincreasedcapabilitytorecyclethehighamounts ofCO2 releasedduringtheclimactericrise. This phenomenonhas also beenreportedto occur notonlyin tomatofruits butalso in other nonleaforganssuchasflowers, buds, fruits,etc. (24, 33)
The chloroplast SOD which functions as a detoxifier of su- peroxideradicals has also beenfollowed. Thisenzyme reached itsmaximallevelonlyaftertheconversion tochromoplast(stage f),aboutthetime oftheclimactericrespiratory rise.This riseof superoxide dismutase wouldbe expected since the levels ofsu- peroxideradicalssignificantlyincreaseinsenescing tissues.
Acknowledgments-Theauthors thankDrs.BenjaminHorwitz andE. B. Dum- broff forcriticalreadingofthemanuscript.
LITERATURE CITED
1. BATHGATEB,MEPURTON,DGRIERSON, PWGOODENOUGH 1985Plastid
changes duringthe conversion ofchloroplaststochromoplastinripening tomatoes.Planta 165: 197-204
2. BEANRC,GW TODD 1959Photosynthesisandrespirationindevelopingfruits.
PlantPhysiol34:425-429
3. BEN DAVIDH, N NELSON, S GEPSTEIN 1983 Differential changesin the amountofprotein complexesin thechloroplastmembraneduringsenescence ofoatandbean leaves. PlantPhysiol 73:507-510
4. BENEDICTCR,CLROSENFIELD,JRMAHAN,SMADHAVAN,H YOKOYAMA 1985The chemicalregulationof carotenoidbiosynthesisin citrus. PlantSci 41:169-173
5. BERTHOLD DA1981Ahighly resolved oxygen-evolving PSIIpreparationsfrom
242 LIVNEANDGEPSTEIN
spinach thylakoidmembranes EPR and electron transportproperties.FEBS Lett 134: 231-234
6. BRADYCJ,GMACALPINE,WBMcGLAssoN,Y UEDA 1982Polygalacturonase intomatofruitsand the induction ofripening.Aust J PlantPhysiol9: 171- 178
7. CAMARA B,J BRANGEON 1981 Carotenoidmetabolismduringchloroplastto chromoplast transformation inCapsicumannuumfruit. Planta 151: 359-364 8. DOSTAL HC, AC LEOPOLD 1967 Gibberellin delays ripening of tomatoes.
Science 158: 1579-1580
9. DOUGLAS MG,RA BUTOW 1976 Variant forms ofmitochondrial translation products inyeast. Evidence for location of determinantsonmitochondrial DNA.Proc Natl Acad Sci USA 73: 1083-1086
10. ELLIs RJ 1981 Chloroplast proteins synthesis,transportandassembly. Annu Rev PlantPhysiol32: 111-137
11. FAIRINEAUJ,D LAVEL-MARTIN 1977Lightversusdark carbon metabolism incherrytomatofruit.I.Relationshipbetween malate metabolism andpho- tosynthetic activity.PlantPhysiol60:877-880
12. GRIERSOND 1981 Control of ribonucleic acid andenzymesynthesis during fruitripening. InMLieberman, ed,Post HarvestPhysiologyandCropPres- ervation.NATO AdvancedStudyInstitutesSeries,Series A: Life Sciences 46,pp45-60
13. HARRIS WM,ARSPURR1969aChromoplastsoftomatofruitI,ultra-structure of lowpigment and high-betamutants: carotene analyses.Am J Bot 56:
369-379
14. HARRISWM,ARSPURm 1969bChromoplastsoftomatofruitII,the redtomato.
AmJ Bot 56: 380-389
15. HURT E, G HAUSKA 1982Identification of thepolypeptidesin thecytochrome B6/f complexfromspinach chloroplastswithredox-center-carryingsubunits.
JBioenergBiomembr 14(5/6):405-424
16. IWATSUKI N, R MORIYAMA,T ASAHI1984 Isolation andpropertiesofintact chromoplastsfromtomatofruits.PlantCellPhysiol25:763-768 17. LAVAL-MARTIND,JFARINEAU,J DIAMOND 1977Lightversusdark carbon
metabolism incherrytomatofruit. PlantPhysiol60: 872-876
18. LIVEANUV,CFYOCUM,N NELSON 1986Polypeptidesof theoxygen-evolving photosystemII complex: immunologicaldetection andbiogenesis. J Biol Chem 261: 5296-5300
19. NECHUSHTAI R,N NELSON1985BiogenesisofphotosystemI reactioncenter during greeningofoatbean andspinachleaves. Plant Mol Biol 4: 377-384 20. NECHUSHTAIR,NNELSON,AKMATTOO,MEDELMAN 1981 Siteofsynthesis of subunits tophotosystem I reactioncenterand the proton ATPase in Spirodela.FEBS Lett 125: 115-119
21. NELSON N 1983 Structure and synthesis ofchloroplast ATPase. Methods En- zymol 97: 510-523
22. PIECHULLAB, E PICHERSKY, AR CASHMORE, W GRUISSEM1986 Expression of nuclearand plastidgenesfor photosynthesis-specificproteins duringto- matofruitdevelopment and ripening. Plant Mol Biol 7: 367-376 23. PIECHULLAB, KRCHONOLES IMLAY, W GRUISSEM 1985 Plastidgene expres-
sion duringfruit ripening intomato.PlantMol Biol 5: 373-384
24. RHODES MJC 1980 Respiration andsenescenceofplantorgans.InEC Conn, PStumpf, eds,TheBiochemistry ofPlants, Vol 2. Academic Press, New York,pp419-462
25. ROBERTS DR,JE THOMPSON,EB DUMBROFF, S GEPSTEIN, AK MATOO 1987 Differential changes in the synthesis and steady statelevels of thylakoid proteins during bean leafsenescence.Plant Mol Biol. 9: 343-353 26. ROTH R, N NELSON 1981 Purification and immunologicalproperties ofproton-
ATPase complexes fromyeastandratliver mitochondria. J BiolChem 256:
9224-9228
27. SACHER JA 1973 Senescence and postharvest physiology. Annu Rev Plant Physiol24:197-224
28. SAWAMARA M, E KNEGT, J BRUINSMA 1978 Levels of endogenousethylene, carbondioxide and soluble pectin and activities of pectin methylesteraseand polygalecturonase in ripeningtomatofruits. Plant Cell Physiol19: 1061- 1069
29. SHAALTIEL Y, NH CHUA, S GEPSTEIN, J GRESSEL 1987 Dominantpleiotropy controlsenzymesco-segregating withparaquat resistance in Conyzabon- ariensis.TheorApplGenet. Inpress
30. SPEIRS J, CJ BRADY, D GRIERSON, E LEE 1984 Changes in ribosomeorga-
nizationandmessengerRNAabundance in ripeningtomatofruits.AustJ Plant Physiol 11: 225-233
31. TOWBINH, T STAEHELIN, J GORDAN 1979 Electrophoretic transferof proteins frompolyacrylamide gelstonitrocellulose sheets: procedureandsomeap-
plications.ProcNatlAcad Sci USA 76: 4350-4357
32. UEDANK, T SUGIYAMA 1976 Purification and characterization ofphosphoenol pyruvatecarboxylase from maize leaves. Plant Physiol 571:906-910 33. VuJCuV, G YELENOSKY, MG BAUSHER 1985 Photosynthetic activityin the
flowerbuds of ValenciaorangeCitrussinensis(L.) Osbeck. Plant Physiol.
78:420-423
34. WILLMERCM,WRJOHONSTON 1976 Carbon dioxide assimilation insome
serialplantorgansand tissues. Planta 130:33-37
35. ZEMELE, S GEPSTEIN 1985 Immunological evidence for thepresenceof ri- bulosebisphosphate carboxylase in guard cell chloroplasts.Plant Physiol 78:
586-590