Abstract
Alcohol use disorders (AUDs) represent one of the foremost public health concerns in the
United States in which alcohol-associated cues can attract excessive attention and contribute to
relapse. This attentional bias (AB) to alcohol cues has been widely documented but the
neurobiology remains poorly understood. In rats, we used Pavlovian conditioning to pair a light
cue with an alcoholic or non-alcoholic reward and assessed AB through Pavlovian conditioned
responses (CRs) to reward-predictive cues. Rats were exposed to chronic intermittent ethanol
(CIE) prior to Pavlovian conditioning to test whether CIE increases alcohol AB. We also
administered naltrexone (NTX), a drug used to treat AUDs, to see its effects on CRs. Our results
suggest that rats exhibit reduced CRs to alcoholic versus non-alcoholic rewards. Moreover, CIE
and NTX had minimal effect in changing alcohol AB. These findings may provide a framework
for future experimentation on neural pathways and processes involved in alcohol AB.
Introduction
Alcohol use disorders (AUDs) represent a pervasive public health issue that affects 17
million American adults.1 AUDs are characterized by compulsion to consume alcohol despite
negative consequences. Alcohol abuse may account for as many as 9.8% of deaths among
working-age American adultsand poses a $223.5 billion annual cost.2,3 Binge drinking, which is
defined as women consuming four or more drinks or men consuming five or more drinks in a
short time period, is responsible for at least half of all alcohol-attributable deaths and up to
three-quarters of the economic costs of alcohol abuse.2 Properly addressing the public health issues of
Because human studies on the effects of binge drinking are difficult and have strict
limitations, use of animal models is necessary to assess consequences of binge-like alcohol
exposure. Chronic intermittent ethanol (CIE) exposure is a technique used to model binge-like
drinking behavior in animals. CIE has been known to increase alcohol drinking and promote
reward-seeking behavior.4 Furthermore, mice previously exposed to CIE voluntarily consume
alcohol faster and, in the process, reach higher brain ethanol concentrations.5 One area for
exploration is how CIE affects attention to alcohol-related cues.
Alcohol-related cues can attract excessive attention from people after repeated exposure
to alcohol in the presence of the cues.6,7 An attentional bias (AB) to alcohol cues may be
acquired through Pavlovian learning7,8 since repeated pairings of these alcohol cues with alcohol
can result in these cues being able to control attention and trigger behaviors such as alcohol
seeking and consumption, which may ultimately promote relapse.9,10 The strength of AB towards
alcohol-related cues may even be able to predict relapse risk,11 alcohol craving,12,13 and drinking
frequency.14 One pharmaceutical treatment thought to mediate AB is the opioid receptor
antagonist, naltrexone (NTX). NTX is known to reduce drug-seeking behavior brought about by
drug-cue pairing in animal models15,16,17 and to reduce cue-induced alcohol cravings in
humans.18,19
While the clinical relevance of AB is widely accepted, the neural bases of alcohol AB
have yet to be uncovered. The major hurdle in studying neural mechanisms of AB is the
difficulty to model alcohol AB in rodents. Compulsion to drink can be elicited through exposure
to alcohol-related cues, which acquire salience through Pavlovian conditioning.9,10 Since
Pavlovian conditioned responses (CRs) have been widely used to study reward-predictive cues in
more invasive neurobiological measurements and manipulations than would be permissible in
human subject research.
The central aims of this experiment were to identify how CIE affects Pavlovian CRs to
alcoholic vs. non-alcoholic rewards and to provide a framework for future experimentation on
the neural circuitry and neuromodulation involved in alcohol AB. We hypothesized that a history
of binge-like CIE would increase AB to alcoholic cues vs. non-alcoholic cues and that NTX
would blunt this AB to alcohol cues. Therefore, we expected rats to show greater CRs to
alcoholic cues after CIE exposure and reduced CRs after NTX injection. To address these
hypotheses, we divided adult male rats into 3 exposure groups: CIE, water (H2O), or
non-manipulated (NM). After their assigned treatment, rats underwent Pavlovian conditioning
training to either an alcoholic or non-alcoholic reward. The reward was paired with a cue
consisting of a light and lever presentation. After initial Pavlovian conditioning, rats were given
an extinction session and later, rats were given NTX or saline prior to Pavlovian conditioning
sessions. Extensive video analysis was used to determine CRs from these sessions.
Methods Subjects:
A total of 56 male Sprague-Dawley rats (Charles River Labs, NC) were used in this 3 x 2
factorial study (exposure: CIE, H2O, NM; reward: Ensure® + ethanol (EtOH) [Alc], plain
Ensure® [N-alc]) with n = 8-10 per group. The animals were maintained in a climate-controlled
vivarium with ad libitum access to food and water. Animals were pair-housed with partners of
the same exposure condition during the exposure period and were then isolate housed prior to
of Laboratory Animals established by the National Institutes of Health (8th Edition), using
protocols approved by the University of North Carolina Institutional Animal Care and Use
Committee.
Procedure:
Exposure: All animals were handled twice prior to exposure for approximately five minutes so
that they would be accustomed to human contact. The CIE group received a total of 14 doses of
25% v/v EtOH in H2O intragastrically on a Monday, Tuesday, Thursday, and Friday schedule for
3.5 weeks. The first two doses were 3 g/kg and 4 g/kg respectively, while all 12 subsequent
doses were 5 g/kg. The H2O group received an equivalent volume of water on the same schedule
as the CIE group. The NM group was weighed before and after the 3.5-week exposure period.
Tail blood samples were taken from the CIE and H2O-treated animals twice during the exposure
period and used for blood ethanol concentration (BEC) measurements with an AM1 Alcohol
Analyzer (Analox Instruments, Lunenburg, MA).
Pavlovian conditioning: The Pavlovian procedures began in operant chambers (MedAssociates,
St. Albans, VT) one week after CIE exposure. Rats were isolate-housed and given a tasting
sample of plain chocolate Ensure solution in their home cages to make sure they would consume
the reward and to prevent neophobia. The rats were then “pre-trained” on two sessions (~40 min
long) of 25 rewards given on a variable interval 90s (VI90) schedule to establish where the
reward (Ensure + 10% EtOH or plain Ensure) was dispensed. Next, rats underwent six Pavlovian
conditioned approach (PCA) sessions (~40 min long) of 25 rewards in which the cue (stimulus
reward. These sessions were used to assess the acquisition and maintenance of CRs on a VI90
schedule. On day 7, the cues were presented on the same VI90 schedule, but rewards were
withheld (extinction) to assess changes in the latency and duration of CRs. Following these
initial PCA sessions, rats underwent surgery and testing for a separate experiment. Rats then
underwent a week of five more PCA sessions with reward presentation. On days 3 and 5 of that
week, rats were injected with 1 mg/kg of either NTX or saline (30 min prior to the session, s.c.,
balanced order). The amount of any fluid remaining in the cup and the number of lever presses
during cue presentation were measured after each PCA session. Tail blood samples for BEC
measurements were taken on the first day of the final week of PCA sessions immediately after
the session concluded.
Behavior analysis: All PCA sessions were videotaped and scored for the following variables:
reward delivery cup (unconditioned stimulus [US]) approach latency, cue (conditioned stimulus
[CS]) approach latency, time spent at the cup during cue presentation (US duration), time spent
at the cue during cue presentation (CS duration), number of entries to the cup zone, and number
of entries to the cue zone. To establish whether CRs were specific to the cue onset, baseline
behavior for the dependent variables was measured as the behavior in the 8s preceding the cue.
An elevation score was calculated as the change in behavior during the cue as compared to the
baseline. A positive score indicates that the rat spent more time engaging in the activity during
Statistics:
All data were analyzed using two-way and three-way repeated-measures ANOVAs in
SPSS (IBM, New York, NY). Tukey’s honest significance tests at the 95% confidence interval
were used for post-hoc analyses.
Results
BEC Measurements: We collected tail-blood samples on two occasions after exposure fluid was
administered to ensure binge-level BECs. BECs of H2O-exposed rats were taken to be the
baseline readings from the analysis instrument. These data are shown in Table 1. We also
collected blood samples after PCA sessions to see if exposure group affected BECs. These data
are shown in Table 2. There was not a significant effect of exposure on BECs.
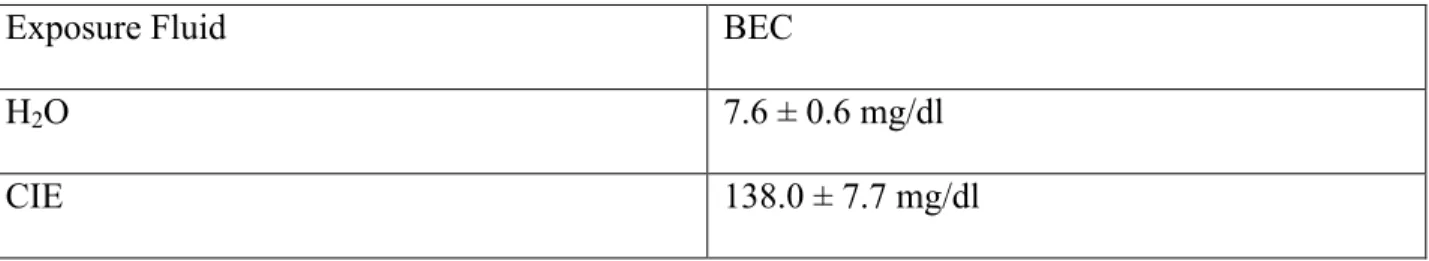
PCA Day 6: We looked at six measures of conditioned responses on the sixth day of Pavlovian
conditioning training to assess whether the rats were attributing salience to the cue. Analysis of
video data from the sixth Pavlovian conditioning session showed rats were responding to the cue
as shown by positive elevation scores for conditioned responses to the cue. While there was no
significant effect of exposure or interaction between exposure and reward on conditioned
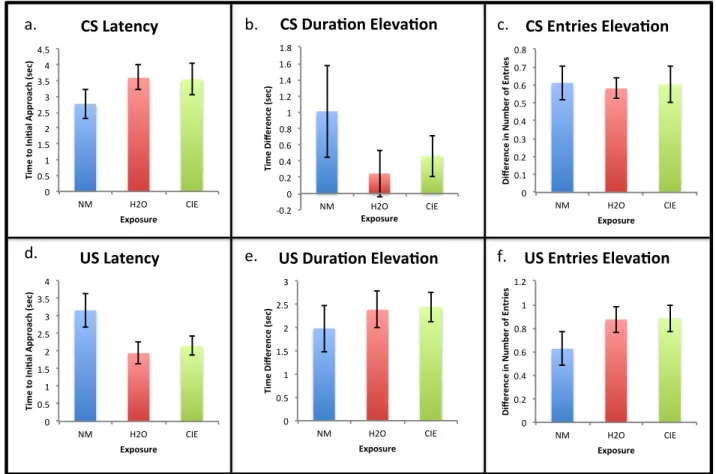
responses to the cue (Figure 1), there was a main effect of reward solution on latency to the
conditioned stimulus (CS) [F, (1, 51) = 5.471; p = 0.024]. Rats approached the cue zone 1.2
seconds faster for the plain nonalcoholic reward than for the alcoholic reward (Figure 2a). There
were no significant effects or interactions of exposure or reward on duration or number of CS or
PCA extinction: Rats underwent an extinction session where reward was not administered to
determine if exposure or reward would alter CRs under these conditions. Analysis of the video
data from the PCA extinction session showed that rats still exhibited positive elevation scores,
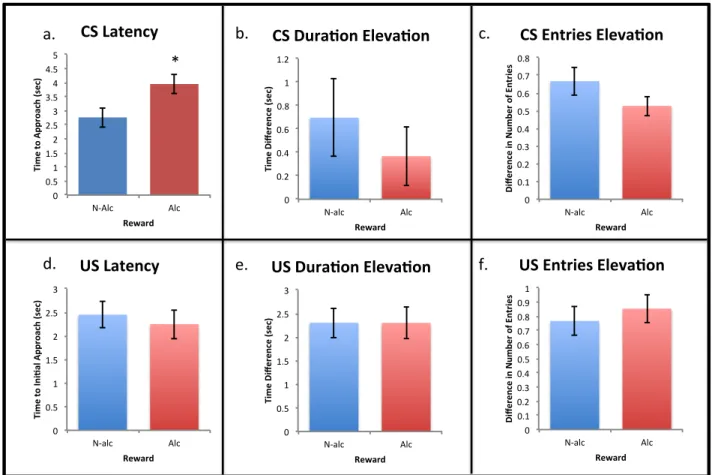
which signifies rats displayed conditional responses to the cues. The only main effect of
exposure on CRs was in latency to US approach [F, (2, 52) = 3.448; p = 0.04]. Compared to NM
rats, H2O and CIE rats tended to show decreased US latency, although this effect only reached
significance in the CIE group (p = 0.018; Figure 3d). The only main effect of reward on
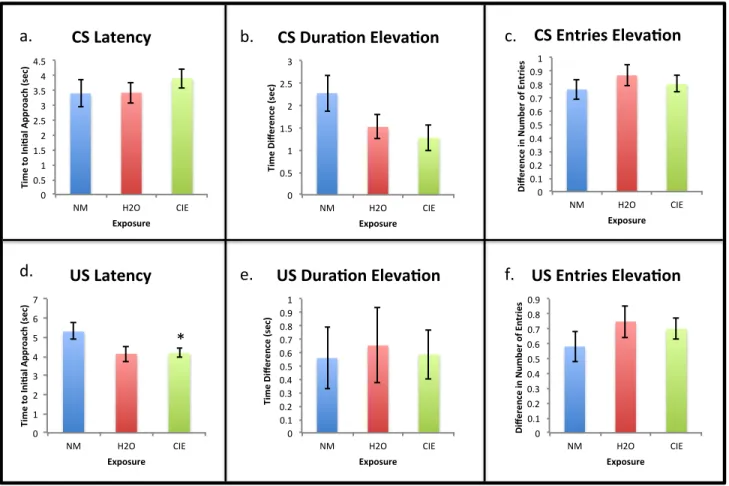
conditioned responses was in latency to the CS [F, (2, 52) = 4.154; p = 0.047]. As they did on
Day 6, rats approached the cue 0.85 seconds faster when presented with a non-alcoholic reward
compared to the alcoholic reward (Figure 4a). There were no significant effects or interactions of
exposure or reward on duration or number of CS or US zone entries, or on the latency to
approach the US (Figures 4b-f).
NTX: Rats were then given an injection of NTX or saline prior to PCA to see how NTX might
alter CRs to reward cues. Analysis of the video data from the NTX- and saline-treatment sessions
showed rats displaying conditioned responses to reward cues. Elevation scores remained
positive. 3-way repeated-measures ANOVAs revealed no main effect of NTX on any of the
measures for CRs (Figure 5), but showed several effects of exposure and reward solution when
collapsed across injection treatment.
The data showed a main effect of reward [F, (1, 91) = 15.602; p < 0.001] and an exposure
x reward interaction effect [F, (2, 91) =9.638; p < 0.001] on latency to the CS. The interaction
effect was driven by the NM group, which took significantly longer times (p < 0.001) to
solution on CS latency on Day 6. Furthermore, there was an exposure x reward interaction effect
[F, (2, 91) =3.980; p < 0.023] on elevation score for duration in the CS zone. The interaction
effect was driven by the NM group, which had significantly higher elevation scores (p = 0.006)
for the non-alcoholic reward, showing that NM rats exhibited stronger CRs when the cue was
paired with a non-alcoholic reward (Figure 6b). Similarly, there was an exposure x reward
interaction effect on elevation score for entries into the CS zone [F, (2, 91) =5.384; p = 0.006].
Once again the interaction effect was driven by the NM group, which entered the CS zone
significantly more often (p = 0.002) for non-alcoholic rewards (Figure 6c). Finally, there was an
exposure x reward interaction effect on the elevation score of duration in the US zone [F, (2, 91)
= 8.016; p < 0.001]. This effect was best seen in the NM group, where rats exhibited reduced
elevation scores (p = 0.003) for alcoholic versus non-alcoholic rewards. In contrast, the H2O
group had increased elevation scores (p = 0.013) for alcoholic rewards (Figure 6e). There were
no significant effects or interactions of exposure or reward on the latency to approach the US
(Figures 6d) or on number of US zone entries (Figure 6f).
Discussion
We used Pavlovian conditioning as a method to assess attentional bias to
reward-associated cues and manipulated test conditions to see whether alcohol, when administered either
as a reward during conditioning or when administered intermittently prior to conditioning (CIE),
would alter this attentional bias. We found that rats do attribute salience to cues that are
repeatedly paired with a reward. Over the course of the study, rats displayed positive elevation
scores in number of approaches and time spent at the cue and reward delivery cup. The
an attentional bias to the cues. However, a history of CIE did not significantly affect conditioned
responses to the cue and thus did not seem to change attentional bias. We did find that alcoholic
rewards elicited significantly weaker conditioned responses than did non-alcoholic rewards. This
pattern of behavior may be indicative of a greater attentional bias to cues paired with
non-alcoholic rewards than those paired with non-alcoholic rewards. The differences between reward
groups were more evident in later PCA sessions, i.e. during the NTX and saline treatment
sessions, especially in the NM group. Our final observation was that NTX did not significantly
affect conditioned responses to the cue compared to saline for any reward. Contrary to results
from previous studies showing that NTX decreased alcohol-seeking behavior,17 NTX did not
decrease attentional bias to cues paired with alcoholic rewards under the present conditions.
A major goal of this study was to develop a model to mimic attentional bias in animal
models. While the model used in this study did not corroborate previous results, it did show
promise for future experimentation if a few changes can be made. The experimental timeline in
this study posed difficulties that could be mitigated in the future. It was difficult to directly
compare data from the final week of PCA to data from the first seven PCA sessions because rats
underwent surgery and were subject to electrochemical measurements in between these phases.
This design may have added additional variables with unanticipated effects on behavioral data. It
may be necessary to complete all PCA sessions not requiring invasive measurements prior to
surgery to minimize these effects. Moreover, the exposure period could be modified for future
CIE experimentation with this model. While the BECs for CIE-exposed rats met the criteria for
binge-like consumption at 138.0 ± 7.7 mg/dl, the stressful effect of intragastric intubation may
have impacted results (e.g., differences between the H2O and NM groups). Alternative
more suitable for future studies. One particular technique may be vapor exposure to alcohol for
CIE. Vapor inhalation allows animals to be exposed to ethanol in a stable environment and when
administered after an injection of pyrazole, an alcohol dehydrogenase inhibitor, it gives the
investigator greater control over BECs.20 Additionally, vapor-exposed animals can be compared
directly to air-exposed controls,5 thereby reducing the number of control subjects required for the
current design.
Furthermore, while the effects of CIE during adulthood on attentional bias were
inconclusive in this study, CIE during adolescence has been shown to produce long-term changes
in the brain21 and could play a role in increasing attentional bias. Binge drinking has become
more common among adolescents in the past decade with 20.8% of high school-aged children
reporting having had five or more drinks on one occasion.22 Studies on adolescent intermittent
ethanol (AIE) exposure in rats have shown that exposure to ethanol during adolescence increases
alcohol intake in adulthood.23 Additionally, AIE in rats has been shown to increase sign-tracking
behavior during Pavlovian conditioned approach training in adulthood.24 An increase in
sign-tracking behavior may represent a strengthening of attentional bias to alcohol cues. Further
application of Pavlovian conditioning in adolescent rats with long-term binge ethanol exposure
may provide insight on whether the age during which binge drinking takes place has an effect on
attentional bias.
Ultimately, understanding the effects of alcohol exposure on attentional bias will be
critical in finding novel treatment options for AUDs. Pavlovian conditioning can be paired with
numerous modern techniques to investigate the neural processes and physiological
manifestations of attentional bias. One technique that has been used consistently by this
release in response to a salient cue while an animal is in an operant chamber.25In vivo
microdialysis offers a second option for measuring neurotransmitter release in the brain through
analysis of extracellular fluid samples.26 FSCV has already been used to show that CIE reduces
dopamine transmission in mice,27 while microdialysis has revealed that NTX inhibits prolonged
dopamine release in rats.28 Combining the Pavlovian conditioning model with FSCV or
microdialysis would allow analysis of how changes in dopamine transmission during the
presentation of the cue may be correlated with attentional bias.
Finally, we can use techniques to manipulate certain brain regions to see how they are
involved in attentional bias. In vivo optogenetics provides a means of upregulating or
downregulating neural processes in specific regions of the brain using opsin proteins designed to
respond to different light frequencies.29 When used in combination with Pavlovian conditioning,
optogenetics can tell us which neural pathways are involved in developing conditioned responses
and ultimately can shed more light on how attentional bias may proceed and which
pharmaceutical agents might best combat AB. Gathering this information can help medical
professionals tailor treatment approaches to fit AUD patients based on the extent to which they
have developed attentional bias to alcohol cues.
In conclusion, this study provided a means to investigate how attentional bias can be
modeled in rats and sought to identify what effects exposure to CIE and administration of NTX
might have on AB. While we saw that rats attributed salience to the cue after Pavlovian
conditioning, we did not find any effects of CIE exposure or NTX on conditioned responses to
the cue. We took these findings to mean that rats did develop an attentional bias to
reward-predictive cues, but it is inconclusive whether binge-like alcohol exposure or administration of a
to cues paired with non-alcoholic rewards than to cues paired with alcohol cues. In the future we
will use Pavlovian conditioned approach training to analyze more specific aspects of the neural
mechanisms of attentional bias. These types of studies may aid in identifying the most effective
treatment methods to control relapse risk in people with AUDs.
Acknowledgments
I would like to thank the members of the Robinson lab for their help and support with this
project over the last two years. I am especially grateful to Dr. Donita Robinson and to my direct
mentor, Dr. Maggie Broadwater, for all the time and effort they have put into helping me
improve as a student and researcher. I offer special thanks to the Bowles Center for Alcohol
Studies and to Dr. Fulton Crews for the grant proposal for this project. Additionally, I would like
to thank the UNC department of biology and specifically Dr. John Bruno and Dr. Gidi Shemer
for overseeing this thesis project and countless others. Finally, I want to express my gratitude to
my parents, Radhika and Raghunatha Vemuru, and the rest of my family and friends for their
References
1. SAMHSA. Substance Dependence or Abuse in the Past Year among Persons Aged 18 or Older, by Demographic Characteristics: Numbers in Thousands, 2011 and 2012. National
Survey on Drug Use and Health (NSDUH). 2012.
2. Stahre M, Roeber J, Kanny D, Brewer RD, Zhang X. Contribution of excessive alcohol consumption to deaths and years of potential life lost in the United States. Prev Chronic Dis. 2012; E109(11).
3. Bouchery EE, Harwood HJ, Sacks JJ, Simon CJ, Brewer RD. Economic costs of
excessive alcohol consumption in the United States, 2006. Am J Prev Med. 2011; 41(5): 516–24.
4. DePoy L, Daut R, Wright T, et al. Chronic alcohol alters rewarded behaviors and striatal plasticity. Addiction Biology. 2015; 20(2): 345-348.
5. Griffin WC, Lopez MF, Yanke AB, Middaugh LD, Becker HC. Repeated cycles of chronic intermittent ethanol exposure in mice increases voluntary ethanol drinking and ethanol concentrations in the nucleus accumbens. Psychopharmacology. 2009; 201(4): 569-580.
6. Townshend JM, Duka T. Attentional bias associated with alcohol cues: differences between heavy and occasional social drinkers. Psychopharmacology. 2001; 157(1): 67-74.
7. O’Brien CP, Childress AR, Ehrman R, Robbins SJ. Conditioning factors in drug abuse: can they explain compulsion? J Psychopharmacol. 1998; 12: 15-22.
8. Tiffany ST. A cognitive model of drug urges and drug-use behavior: role of automatic and nonautomatic processes. Psychological review. 1990; 97(2): 147-168.
9. Stacy AW, Wiers RW. Implicit cognition and addiction: a tool for explaining paradoxical behavior. Annual review of clinical psychology. 2012; 6: 551-575.
10.Robinson TE, Berridge KC. Addiction. Annual review of psychology. 2003; 54: 25-53. 11.Cox WM, Hogan LM, Kristian MR, Race JH. Alcohol attentional bias as a predictor of alcohol abusers’ treatment outcome. Drug and alcohol dependence. 2002; 68 (3): 237-243.
12.Field M, Mogg K, Bradley BP. Craving and cognitive biases for alcohol cues in social drinkers. Alcohol and alcoholism. 2005; 40(6): 504-510.
13.Field M, Munafó MR, & Franken IH. A meta-analytic investigation of the relationship between attentional bias and subjective craving in substance abuse. Psychological
bulletin. 2009; 135(4): 589-607.
14.Field M, Eastwood B. Experimental manipulation of attentional bias increases the motivation to drink alcohol. Psychopharmacology. 2005; 183(3): 350-357.
15.Herz A. Endogenous opioid systems and alcohol addiction. Psychopharmacology. 1997; 129(2): 99-111.
16.Ciccocioppo R, Martin-Fardon R, Weiss F. Effect of selective blockade on mu(1)or delta opioid receptors on reinstatement of alcohol-seeking behavior by drug-associated stimuli in rats. Neuropsychopharmacology. 2002; 27(3): 391-399.
18.Rohsenow DJ, Monti PM, Hutchison KE, Swift RM, Colby SM, Kaplan GB. Naltrexone's effects on reactivity to alcohol cues among alcoholic men. Journal Of
Abnormal Psychology. 2000; 109(4): 738-742.
19.Monti PM, Rohsenow DJ, Hutchison KE, et al. Naltrexone's Effect on Cue-Elicited Craving Among Alcoholics in Treatment. Alcoholism: clinical and experimental
research. 1999;23(8): 1386-1394.
20.Knapp DJ, Breese GR. Models of chronic alcohol exposure and dependence. Psychiatric
Disorders. 2011; 829: 205-230.
21.Pascual M, Pla A, Minarro J, Guerri C. Neuroimmune activation and myelin changes in adolescent rats exposed to high-dose alcohol and associated cognitive dysfunction: A review with reference to human adolescent drinking. Alcohol and Alcoholism. 2014; 49(2): 187-192.
22.Centers for Disease Control and Prevention. Youth Risk Behavior Survey – United States, 2013. Morbidity and Mortality Weekly Report Surveillance Summaries. 2014; 63(4): 1-168.
23. Pascual M, Boix J, Felipo V, and Guerri C. Repeated alcohol administration during adolescence causes changes in the mesolimbic dopaminergic and glutamatergic systems and promotes alcohol intake in the adult rat. Journal of Neurochemistry. 2009; 108: 920– 931.
24.McClory AJ, Spear LP. Effects of ethanol exposure during adolescence or in adulthood on Pavlovian conditioned approach in Sprague-Dawley rats. Alcohol. 2014; 48(8): 755-763.
25.Willuhn I, Wanat MJ, Clark JJ, Phillips PEM. Dopamine Signaling in the Nucleus Accumbens of Animals Self-Administering Drugs of Abuse. Current Topics in
Behavioral Neurosciences. 2010; 3: 29–71.
26.Darvesh AS, Carroll RT, Geldenhuys WJ, et al. In vivo brain microdialysis: advances in neuropsychopharmacology and drug discovery. Expert Opinion on Drug Discovery. 2011; 6(2): 109–127.
27.Karkhanis AN, Rose JH, Huggins KN, Konstantopoulos JK, Jones SR. Chronic intermittent ethanol exposure reduces presynaptic dopamine neurotransmission in the mouse nucleus accumbens. Drug and Alcohol Dependence. 2015; Epub.
28.Valenta JP, Job MO, Mangieri RA, Schier CJ, Howard EC, Gonzales RA. Mu-opioid receptors in the stimulation of mesolimbic dopamine activity by ethanol and morphine in long-evans rats: A delayed effect of ethanol. Psychopharmacology. 2013; 228(3): 389-400.
29.Sidor MM, Davidson TJ, Tye KM, Warden MR, Diesseroth K, McClung CA. In vivo optogenetic stimulation of the rodent central nervous system. Journal of Visualized
Tables and Figures
Exposure Fluid BEC
H2O 7.6 ± 0.6 mg/dl
CIE 138.0 ± 7.7 mg/dl
Table 1: Mean BECs of rats given exposure fluids intragastrically from tail-blood samples taken 60 minutes after doses 4 and 13.
Exposure Group BEC
NM 11.7 ± 2.5 mg/dl
H2O 21.0 ± 7.8 mg/dl
CIE 29.6 ± 7.5 mg/dl
Figure 1: Conditioned responses to cues on the sixth session of Pavlovian conditioning (Day 6) collapsed across reward solution. There were no significant differences between exposure groups for a) time to initial approach of the CS zone, b) elevation score for time spent in the CS zone, c) elevation score for number of entries into the CS zone, d) time to initial approach of the US zone, e) elevation score for time spent in the US zone, or f) elevation score for number of entries into the US zone.
0" 0.5" 1" 1.5" 2" 2.5" 3" 3.5" 4" 4.5"
NM" H2O" CIE"
Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Exposure% CS%Latency% 00.2" 0" 0.2" 0.4" 0.6" 0.8" 1" 1.2" 1.4" 1.6" 1.8"
NM" H2O" CIE"
Ti me%Di fferen ce%( sec) % Exposure% CS%Dura*on%Eleva*on% 0" 0.1" 0.2" 0.3" 0.4" 0.5" 0.6" 0.7" 0.8"
NM" H2O" CIE"
Di fferen ce%i n%N umb er%o f%E nt ries % Exposure% CS%Entries%Eleva*on% 0" 0.5" 1" 1.5" 2" 2.5" 3" 3.5" 4"
NM" H2O" CIE"
Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Exposure% US%Latency% 0" 0.5" 1" 1.5" 2" 2.5" 3"
NM" H2O" CIE"
Ti me%Di fferen ce%( sec) % Exposure% US%Dura*on%Eleva*on% 0" 0.2" 0.4" 0.6" 0.8" 1" 1.2"
NM" H2O" CIE"
Di fferen ce%i n%N umb er%o f%E nt ries % Exposure% US%Entries%Eleva*on%
a." b." c."
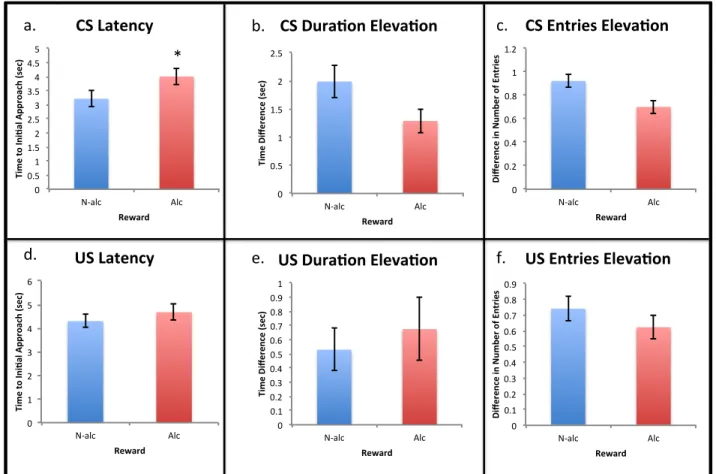
Figure 2: Conditioned responses to cues on the sixth session of Pavlovian conditioning (Day 6) collapsed across exposure group. a) There was a significant increase in time to initial approach to the CS zone for rats given an alcoholic reward compared to whose given a non-alcoholic reward (* p < 0.05). There were no significant differences between reward groups for b) elevation score for time spent in the CS zone, c) elevation score for number of entries into the CS zone, d) time to initial approach of the US zone, e) elevation score for time spent in the US zone, or f)
elevation score for number of entries into the US zone.
0" 0.5" 1" 1.5" 2" 2.5" 3" 3.5" 4" 4.5" 5" N*Alc" Alc" Ti m e% to %Ap pr oa ch %(s ec )%% Reward% CS%Latency% "*" 0" 0.2" 0.4" 0.6" 0.8" 1" 1.2" N*alc" Alc" Ti me%Di fferen ce%( sec) % Reward% CS%Dura<on%Eleva<on% 0" 0.1" 0.2" 0.3" 0.4" 0.5" 0.6" 0.7" 0.8" N*alc" Alc" Di fferen ce%i n%N umb er%o f%E nt ries % Reward% CS%Entries%Eleva<on% 0" 0.5" 1" 1.5" 2" 2.5" 3" N*alc" Alc" Ti m e% to %In i< al %Ap pr oa ch %(s ec )% Reward% US%Latency% 0" 0.5" 1" 1.5" 2" 2.5" 3" N*alc" Alc" Ti me%Di fferen ce%( sec) % Reward% US%Dura<on%Eleva<on% 0" 0.1" 0.2" 0.3" 0.4" 0.5" 0.6" 0.7" 0.8" 0.9" 1" N*alc" Alc" Di fferen ce%i n%N umb er%o f%E nt ries % Reward% US%Entries%Eleva<on%
a." b." c."
Figure 3: Conditioned responses to cues during the extinction session of Pavlovian conditioning (Day 7) collapsed across reward. d) There was a significant decrease in time to initial approach to the US zone for rats exposed to CIE compared to NM rats (* p < 0.05). There were no
significant differences between exposure groups for a) time to initial approach of the CS zone, b) elevation score for time spent in the CS zone, c) elevation score for number of entries into the CS zone, e) elevation score for time spent in the US zone, or f) elevation score for number of entries into the US zone.
0" 0.5" 1" 1.5" 2" 2.5" 3" 3.5" 4" 4.5"
NM" H2O" CIE"
Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Exposure% CS%Latency% 0" 0.5" 1" 1.5" 2" 2.5" 3"
NM" H2O" CIE"
Ti me%Di fferen ce%( sec) % Exposure% CS%Dura*on%Eleva*on% 0" 0.1" 0.2" 0.3" 0.4" 0.5" 0.6" 0.7" 0.8" 0.9" 1"
NM" H2O" CIE"
Di fferen ce%i n%N umb er%o f%E nt ries % Exposure% CS%Entries%Eleva*on% 0" 1" 2" 3" 4" 5" 6" 7"
NM" H2O" CIE"
Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Exposure% US%Latency% 0" 0.1" 0.2" 0.3" 0.4" 0.5" 0.6" 0.7" 0.8" 0.9" 1"
NM" H2O" CIE"
Ti me%Di fferen ce%( sec) % Exposure% US%Dura*on%Eleva*on% 0" 0.1" 0.2" 0.3" 0.4" 0.5" 0.6" 0.7" 0.8" 0.9"
NM" H2O" CIE"
Di fferen ce%i n%N umb er%o f%E nt ries % Exposure% US%Entries%Eleva*on% """""*"
a." b." c."
Figure 4: Conditioned responses to cues during the extinction session of Pavlovian conditioning (Day 7) collapsed across exposure group. a) There was a significant increase in time to initial approach to the CS zone for rats given an alcoholic reward compared to whose given a non-alcoholic reward (* p < 0.05). There were no significant differences between reward groups for b) elevation score for time spent in the CS zone, c) elevation score for number of entries into the CS zone, d) time to initial approach of the US zone, e) elevation score for time spent in the US zone, or f) elevation score for number of entries into the US zone.
0' 0.5' 1' 1.5' 2' 2.5' 3' 3.5' 4' 4.5' 5' N1alc' Alc' Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Reward% CS%Latency% 0' 0.5' 1' 1.5' 2' 2.5' N1alc' Alc' Ti me%Di fferen ce%( sec) % Reward% CS%Dura*on%Eleva*on% 0' 0.2' 0.4' 0.6' 0.8' 1' 1.2' N1alc' Alc' Di fferen ce%i n%N umb er%o f%E nt ries % Reward% CS%Entries%Eleva*on% 0' 1' 2' 3' 4' 5' 6' N1alc' Alc' Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Reward% US%Latency% 0' 0.1' 0.2' 0.3' 0.4' 0.5' 0.6' 0.7' 0.8' 0.9' 1' N1alc' Alc' Ti me%Di fferen ce%( sec) % Reward% US%Dura*on%Eleva*on% 0' 0.1' 0.2' 0.3' 0.4' 0.5' 0.6' 0.7' 0.8' 0.9' N1alc' Alc' Di fferen ce%i n%N umb er%o f%E nt ries % Reward% US%Entries%Eleva*on% ''''*'
a.' b.' c.'
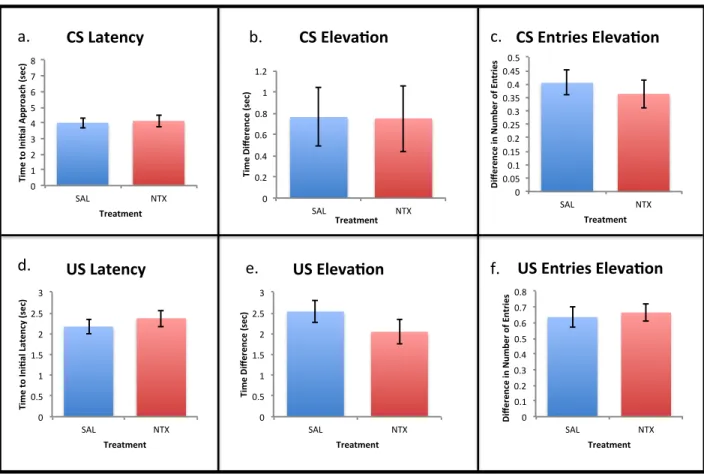
Figure 5: Conditioned responses to cues from the NTX and Saline treatment sessions of Pavlovian conditioning collapsed across exposure and reward. There were no significant
differences between exposure groups for a) time to initial approach of the CS zone, b) elevation score for time spent in the CS zone, c) elevation score for number of entries into the CS zone, d) time to initial approach of the US zone, e) elevation score for time spent in the US zone, or f) elevation score for number of entries into the US zone.
Figure!5:!
0! 1! 2! 3! 4! 5! 6! 7! 8! SAL! NTX! Ti m e% to %In i* al %Ap pr oa ch %(s ec )% Treatment% CS%Latency% 0! 0.2! 0.4! 0.6! 0.8! 1! 1.2! SAL! NTX! Ti me%Di fferen ce%( sec) % Treatment% CS%Eleva*on% 0! 0.05! 0.1! 0.15! 0.2! 0.25! 0.3! 0.35! 0.4! 0.45! 0.5! SAL! NTX! Di fferen ce%i n%N umb er%o f%E nt ries % Treatment% CS%Entries%Eleva*on% 0! 0.5! 1! 1.5! 2! 2.5! 3! SAL! NTX! Ti me%to %In i*al %Laten cy %(sec) % Treatment% US%Latency% 0! 0.5! 1! 1.5! 2! 2.5! 3! SAL! NTX! Ti me%Di fferen ce%( sec) % Treatment% US%Eleva*on% 0! 0.1! 0.2! 0.3! 0.4! 0.5! 0.6! 0.7! 0.8! SAL! NTX! Di fferen ce%i n%N umb er%o f%E nt ries % Treatment% US%Entries%Eleva*on%a.! b.! c.!
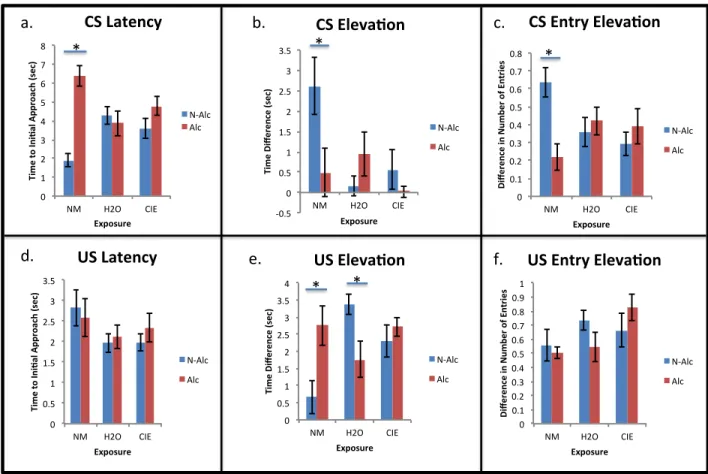
Figure 6: Conditioned responses to cues by exposure group and reward solution, collapsed across NTX and Saline treatment sessions of Pavlovian conditioning. a) NM rats approached the cue zone more quickly when the cue paired with a non-alcoholic reward versus an alcoholic reward (* p < 0.05). b) NM rats also spent a smaller proportion of time near the cue when the cue was illuminated compared to when the cue was off when the cue paired with an alcoholic reward versus a non-alcoholic reward (* p < 0.05). c) NM rats entered the cue zone less frequently when the cue was illuminated compared to when the cue was off for cues paired with an alcoholic reward versus a non-alcoholic reward (* p < 0.05). e) NM rats spent a greater proportion of time near the reward cup when the cue was illuminated compared to when the cue was off when the cue paired with an alcoholic reward versus a non-alcoholic reward (* p < 0.05). H2O-exposed rats spent a smaller proportion of time near the reward cup when the cue was illuminated compared to when the cue was off when the cue paired with an alcoholic reward versus a non-alcoholic reward (* p < 0.05).There were no significant differences between exposure groups for d) time to initial approach of the US zone, or f) elevation score for number of entries into the US zone. !0.5% 0% 0.5% 1% 1.5% 2% 2.5% 3% 3.5%
NM% H2O% CIE%
Ti me%Di fferen ce%( sec) % Exposure% CS%Eleva8on% N!Alc% Alc% 0% 1% 2% 3% 4% 5% 6% 7% 8%
NM% H2O% CIE%
Ti m e% to %In i8 al %Ap pr oa ch %(s ec )% Exposure% CS%Latency% N!Alc% Alc% 0% 0.1% 0.2% 0.3% 0.4% 0.5% 0.6% 0.7% 0.8%
NM% H2O% CIE%
Di fferen ce%i n%N umb er%o f%E nt ries % Exposure% CS%Entry%Eleva8on% N!Alc% Alc% 0% 0.5% 1% 1.5% 2% 2.5% 3% 3.5% 4%
NM% H2O% CIE%
Ti me%Di fferen ce%( sec) % Exposure% US%Eleva8on% N!Alc% Alc% * * %*% * %*%
a.% b.% c.%
d.% e.% f.%
0% 0.5% 1% 1.5% 2% 2.5% 3% 3.5%
NM% H2O% CIE%
Ti m e% to %In i8 al %Ap pr oa ch %(s ec )% Exposure% US%Latency% N!Alc% Alc% 0% 0.1% 0.2% 0.3% 0.4% 0.5% 0.6% 0.7% 0.8% 0.9% 1%
NM% H2O% CIE%