Copyright 0 1983 by the Genetics Society of America
CYTOPLASMIC INHERITANCE
OF A CELL SURFACE ANTIGEN
IN THE MOUSE
KIRSTEN FISCHER LINDAHL A N D BARBARA HAUSMANN Basel Institute for Immunology, Basel, Switzerland
Manuscript received June 25, 1982 Revised copy accepted November 4,1982
ABSTRACT
Mta is a cell surface antigen of the mouse and serves as a target for specific T killer lymphocytes. Using a killer cell assay, the antigen has been found in 72 strains of laboratory mice and, with one exception, in all tested samples of mice caught in the wild or bred from such, including Mus molossinus, Mus castaneus and Mus spretus. Five strains of rats, non-inbred NMRI mice, most substrains of NZB mice and the closely related strain NZO are negative for Mta. In reciprocal FI crosses between several Mta+ and two Mta- strains, the antigen is maternally transmitted; that is, Mta+ females bear only positive offspring, whereas Mta- females bear only negative offspring, regardless of the genotype of the male. Since 34 foster-nursed mice had the Mta type of their genetic mothers, the factor that determines expression of Mta must be transmitted before birth and not via the milk. The cytoplasmic genes of Mta+ strains have been combined with the chromosomal genes of Mta- strains, and vice versa, by repeated backcrossing. All progeny retained the Mta type of their maternal lines. Thus, the Mta type is determined solely by maternal inheritance and is not influenced by chromosomal genes. We found no evidence of incompatibility between the cytoplasmic factors and nuclear genes of Mta- and Mta+ strains.
OST
strains of mice carry the cell surface antigen Mta. It can be detected
M
by cytotoxic T lymphocytes raised in one of the rare negative strains. Mta
is expressed throughout life and has been found
on all lymphoid tissues, on
fibroblasts, and on a variety of tumor cell lines. On the basis of crosses involving
the Mta- strain
NZB
and Mta+ strains C58/J,
BALB/c
and
A . C A ,we concluded
that Mta
is
maternally transmitted
(FISCHER
LINDAHL,
BOCCHIERI
and
RIBLET
1980).
Here we survey the distribution
of
Mta types among various strains of
mice and rats, and we also test how general the maternal transmission
is,
whether Mta is passed by the milk, and whether the Mta type is influenced by
chromosomal genes.
MATERIALS AND METHODS
484
K. FISCHER LINDAHL AND B . HAUSMANNKettering Institute for Cancer Research, New York, N. Y. DR. J. KNIGHT, MRC Immunopathology Research Unit, University of Otago Medical School, Dunedin, New Zealand, sent samples of NZ mice, designated /OU in this paper. ICR/Swiss mice were obtained from DR. K. BURKI, Laboratory of Cell Differentiation, University of Geneva, Switzerland. NFS/N mice came from the animal colony of Ciba-Geigy, Basel, Switzerland. DDK mice were a gift from DR. J.-L. GUENET, Institut Pasteur, Paris, France, and STU mice from DR. W. SCHAFER, Max-Planck-Institut fur Virusforschung, Tubingen, Germany.
Wild mice: Mice caught in four different localities in Denmark were given us by DR. P. HJoRTH (P.H.), Institute for Molecular Biology, University of Aarhus, Aarhus, Denmark, together with mice bred from wild mice. These included Mus costaneus, originally from DR. V. CHAPMAN, Roswell Park Memorial Institute, Buffalo, N. Y., and the strains WID 75, WLA 76 and WPA 77, inbred by DR.
J.-
L. GUENET, Institut Pasteur, Paris, France, from mice caught in southern France. WLA 76 is of pure wild origin, whereas strains WID 75 and WPA 77 were produced by crossing wild males with laboratory females and backcrossing the female offspring to the wild males. From DRS. A. GKOPP and H. WINKING (G&W), Institute for Pathology, University of Liibeck, Liibeck, Germany, we obtained mice of strains CD, Mil 11, Zhadar and Mus spretus, all maintained in small but non-inbred colonies. DR. D. J. SCHENDEL, Institute for Immunology, University of Munich, Munich, Germany, sent us frozen spleen cells, prepared from mice caught for DR. D. BENNETT (D.B.), Sloan-Kettering Institute for Cancer Research, New York, N. Y.Rats: Rats of both sexes, about 2 months old, were obtained from Full or given to us by DR. D. BELLGRAU, Basel Institute for Immunology. The Hooded Lister strain originates from Olac.
Breeding: F1 hybrids between NZB/Bom and NZW/CrBom were bred at Bom. All other crosses were made at this institute. In general, we used two females and one male per cage, and the same male was used for successive backcross generations. We carried two separate lines of each cross in parallel and sampled individuals from both lines, except for the backcrossing of DBA/ZJ to NMRI/ Bom, which was done in a single line. Females were picked for breeding from the first litter of each generation without previous typing for Mta. In the first backcross generation of DBA/Z to NMRI we selected albino females. In the backcrossing of NMRI/Bom to C57BL/6J, we selected agouti FI and black N1 females, thus ensuring homozygosity at the agouti locus (chromosome 2). From the N1 females we selected H-2*'* homozygotes (chromosome 17). The cervical lymph nodes were removed from anesthetized mice, cell suspensions were prepared, and binding of radiolabeled monoclonal antibodies specific for H-ZKh of C57BL/6J (KOHLER, FISCHER LINDAHL and HEUSSER 1981) and for H-Zq of NMRI/Bom mice (LEMKE et al. 1978, FISCHER LINDAHL, HAUSMANN and FLAHERTY 1982) was measured. All mice survived this operation.
Foster-nursing: Pregnant females were killed by cervical dislocation as close as possible to the end of term, and the fetuses were delivered by hysterectomy. They were cleaned and given to foster mothers whose own litters, born a day or two previously, were removed.
Torget cells. I. Mice: Cell suspensions were prepared from spleens (and occasionally lymph nodes) of individual mice and cultured for 2 days with 2 pg/ml concanavalin A (Con A). Before the assay the live lymphoblasts were purified by centrifugation over a Ficoll-Urovison cushion ( p = 1.077 g/ml), washed, and the fraction to be used as hot target cells was labeled with %r, all as previously described (FISCHER LINDAHL and HAUSMANN 1980).
Target cells. II. Rats: Lymph node cells were cultured as above, but with 8 pg/ml Con A, for 2 days. Before the assay they were washed three times in Hanks' BSS with 0.1 M a-methylmannoside, and then processed as mouse target cells. This treatment prevented nonspecific killing or inhibition and did not affect detection of Mta, as tested with mouse target cells.
CYTOPLASMIC INHERITANCE I N MICE
485
Before the assay the viable blasts were purified and labeled with 51Cr as required. They were excellent target cells with a low spontaneous release, and cells from standard inbred strains prepared in the same way typed as expected in the killer assay. Although the recoveries varied greatly (from fewer than 1 Xlo6
to 25 x IO6 blast cells out of 20 x IO6 cells frozen), cryopreservation appears to be a useful method of saving cells from rare mice for study in several experiments.Killer cell assay: Mta- mice were primed with H-2-compatible Mtaf cells (2 x IO7 pooled spleen, lymph node and thymus cells in serum-free medium) by three intraperitoneal injections, given at least 3 wk apart. Their spleen cells were then restimulated in mixed lymphocyte culture with irradiated Mta+ cells for 5 days as previously described (FISCHER LINDAHL and HAUSMANN 1980), except that the medium contained only 10% (v/v) fetal calf serum. The effector cells were then harvested and assayed. Two combinations were used: NZB/Bom anti-NZB/Full (an Mta+ subline of NZB (FISCHER LINDAHL, BOCCHIERI and RIBLET 1980)) and (NMRI/Bom X C57BL/6)Ft anti-C57BL/6. For the direct test, the killer cells (from 3 x IO4, 1 x IO5 and 3 x
lo5
initial responder cells cultured) were mixed with 1 x I O 4 51Cr-labeled target cells in triplicate wells of round-bottom microtiter plates in 200 pl of RPMI 1640 with 10% fetal calf serum. For the competition tests, killer cells from 3 Xlo5
initial responder cells and 1 xlo4
%4abeled target cells of known Mta type were mixed with excess unlabeled cells to be typed (usually 1.5 xlo5,
3 xlo5
and 6 xlo5).
The hot target cells were chosen to differ from the stimulator cells, especially at H-2, to obviate any killing not specific for Mta, and positive and negative controls were included in each assay. The plates were centrifuged 5 min at 20 x g and incubated 3.5 hr at 37'. A sample of supernatant (100 pl) was then collected from each well and the amount of 51Cr released measured in a gamma spectrometer. Results are geometric means of duplicates or triplicates, expressed as a percentage of the maximum release in the presence of a detergent. Spontaneous release was measured from target cells incubated with medium alone. The 95% confidence limits were calculated using FIELLER'S theorem (FINNEY 1964).RESULTS
Strain
survey: Table 1 lists 79 strains of laboratory mice that have been typed
for Mta to date. Since we have found that the sublines NZB/BlPt and its
derivative NZB/Full differed from other NZB mice (FISCHER
LINDAHL,
BOCCHIERI
and
RIBLET
1980), we obtained some NZ mice directly from Otago University.
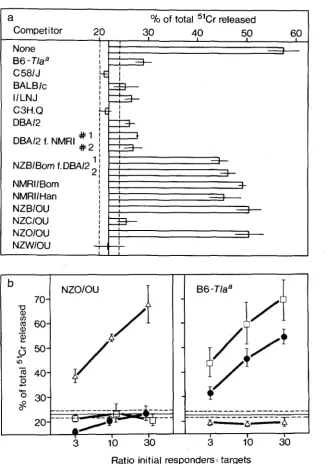
Figure
1
shows the typing of these and other inbred and foster-nursed mice.
The cold target competition assay (Figure l a ) is our most stringent test for a
positive strain. If the cells share Mta with the 51Cr-labeled target cells, they will
compete for the killers and inhibit the release of 51Cr. It is apparent that NZC
and NZW are Mta+. The NZB and NZO strains originated from the same female
in 1948
(BIELCHOWSKY
and GOODALL
1970), and it is therefore not surprising that
both are Mta-. Negative typing results should be confirmed by a direct test
(Figure Ib), since a strain with a cross-reactive form of Mta
or a quantitative
difference in expression might not compete efficiently and could be falsely
scored as negative. Labeled NZO target cells were not affected by killer cells
specific for Mta, but they can be killed by cells specific for H-2" (not shown) or
Qed-lb, both antigens that NZO shares with NZW.
Strain Rb(6.12)3Sic carries a Robertsonian translocation chromosome derived
from a mouse caught in Sicily (GROPP
and WINKING
1981). NMRI females may
have been used in the process of isolating this chromosome, which could explain
their being Mta- (A. GROPP,
personal communication).
486
K.
FISCHER LINDAHL AND B. HAUSMANN TABLE 1Laboratory mouse strains typed for Mta
Negative: NMRI/Bom, NMRI/Ico, NMRI/Han, NZB/BINJ, NZB/CrBom, NZB/Hz, NZB/Icr, NZB/Ola, NZB/OU, NZO/OU, Rb(6.12)3Sic
Positive: A.AL, A.CA, AKR/J, AU/Ssj, BALB.B, BALB.K, BALB/cByJ, BALB/cJ, BDP/J, BUB/ BnJ, B6.AK1, B6-H-Zk, B6.K1, B6.K2, B6-Tla", BlO.A(4R), BlO.A(JR), BlO.BR, BlO.CAS1, BlO,DZ(M504), BlO.DRB6.2, BlO.KPA42, BlO.KPA44, B1O.M. BlO.MOL1, BlO,RIII(71NS), BlO.S, BlO.WB, BlO.Y, CBA/J, CE/J, CJH/HeJ, CJH/Tif, CJH-H-Z", C3H.JK, CJH.Q, C J H S W , C57BL/6J, C57BL/lOScSr, C57L/J, C58/J, DBA/2, DDK, HTI, I/LnJ, I/LnReJ, I/St, ICR/Swiss, LG/J, LP/J, MA/MyJ, NFS/N, NIH/Ola, NZB/BlPt, NZB/Full, NZC/OU, NZW/CrBom, NZW/Ola, NZW/OU, P/ J, PL/J, RF/J, RIIIS/J, SEA/GnJ, SEC/lReJ, SJL/J, SM/J, ST/bJ, STU, SWR, WB/ ReJ, 129/01a
1969; FESTING
1979; MORSE
1981); all other tested descendants of that stock were
Mta+ (ICR/Swiss, NIH/Ola, NFS/N, RF/J, SJL/J and SWWJ).
Wild mice: The mitochondrial DNA of a broad sample of laboratory strains
was identical when analyzed with restriction enzymes, suggesting that the mice
were all derived from the same female ancestor
(YONEKAWA
et
al.
1980; FERRIS,
SAGE
and WILSON
1982). It is therefore not surprising that they also share a
maternally transmitted antigen. Wild mice, on the other hand, represent many
different maternal lines of inheritance. Yet, among 51 mice sampled from 17
locations and several species, we found only a single negative mouse (Table
2).
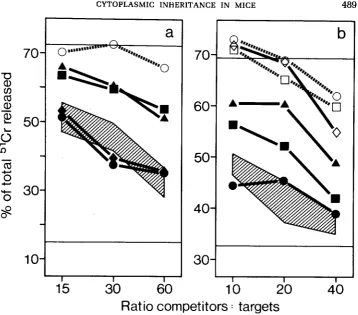
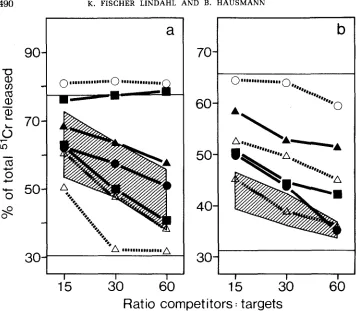
Strains WID 75 and WPA 77 have their genomes from wild mice, but their
maternal line from laboratory mice. They too were Mta+. When possible, the
mice were typed both by cold target competition and direct lysis. Some (WLA
76, Mil I1 and
Mus
spretus) inhibited Mta killing incompletely in all tests (Figs.
2
and
3),
even though they were clearly positive and could be killed directly.
This might be a sign that cells of these strains express less Mta
or, more
interestingly, have a cross-reacting form; however, from the few tests done, we
cannot exclude a random fluctuation in expression, which we see occasionally
also in our standard inbred strains. The finding of two positive and one negative
mouse from the same stock of
Mus castaneus is puzzling and requires further
study.
Rats: We tested cells from five strains of rats: Lewis (Figure
2),F344,
DA, BN
and Hooded Lister. All were negative for Mta in both competition and direct
killing assays.
CYTOPLASMIC INHERITANCE IN MICE
None
B6-Tlaa
C581J
BALBlc
I I I
I
1 - 1
1 1 1 1
1 1
I
*
,
I -
DBA/2
f.
NMRl
# 2
al!E
I1 ;
NZBlBom f.DBA/2
i
i
NMRllHan
-1NZBlOU
NZClOU
NZOlOU
NZWlOU
I INMRll
Bom
1 . : I I I I II I
70-
60-
50-
40-
NZO/OU
T
B6-T/aa
I
487
Ratio initial responders: targets
FIGURE 1.-Mta typing by cold target competition (a) and direct lysis (b). In (a) NZB/Bom anti- NZB/Full killer cells were assayed on 51Cr-labeled B6-Tla" target cells, and unlabeled competitors were added at a ratio of 30 cold to 1 hot target cell. In (b) the NZB/Bom anti-NZB/Fiill killer cells
(0--0) were titrated directly on labeled target cells from NZB/OU and B6-Tla". Killer cells specific for Qed-1" (O----O, CB6F1 anti-BIO.A(5R) (FISCHER LINDAHL, HAUSMANN and FLAHERTY 1982)) and Qed-lb (A--A, B1O.BR anti-C3H/HeJ (FISCHER LINDAHL and HAUSMANN 1980)) served as positive and negative controls. Means and 95% confidence limits of triplicates are shown; the 95%
488
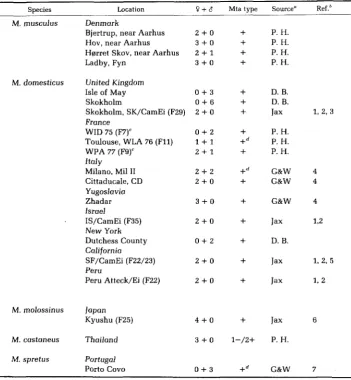
K. FISCHER LINDAHL AND B. HAUSMANNTABLE 2
Mice of wild origin typed for Mta
~
Species Location O + 8 Mta type Source" Ref.*
M. musculus Denmark
Bjertrup, near Aarhus 2 + 0
+
P. H.Hov, near Aarhus 3 + 0
+
P. H.Hgrret Skov, near Aarhus 2
+
1+
P. H.
Ladby, Fyn 3 + 0
+
P. H.M. domesticus United Kingdom
Isle of May Skokholm
Skokholm, SK/CamEi (F29) France
WID 75 (F7)'
Toulouse, WLA 76 (F11)
WPA 77 (F9)' ItaJy Milano, Mil I1 Cittaducale, CD YugosJovia Zhadar lsraeJ
' IS/CamEi (F35)
New York Dutchess County California
SF/CamEi (F22/23) Peru
Peru Atteck/Ei (F22)
0 + 3
+
0 + 6+
2 + 0
+
0 + 2
+
2 + 1
+
1 + 1 +d2 + 2 +d
2 + 0
+
3 + 0
+
2 + 0
+
0 + 2
+
2 + 0
+
2 + 0
+
D. B.
D. B.
Jax 1, 2, 3
P. H. P. H. P. H.
G&W 4
G&W 4
G&W 4
Jax 12
D. B
Jax 1, 2, 5
Jax 1, 2
M. molossinus Japan
Kyushu (F25) 4 + 0
+
Jax 6M . castaneus Thailand 3 + 0 1-/2+ P . H .
M . spretus Portugal
Porto Covo 0 + 3 +d G&W 7
See Materials and Methods, section on Wild Mice.
*
References: 1, WALLACE 1971; 2, WALLACE 1981; 3, FERRIS, SAGE and WILSON 1982; 4, GROPP andWINKING 1981; 5, S A G E 1981; 6, POTTER 1978; 7, WINKING, NIELSEN and GROPP 1980. Maternal line from inbred laboratory mice.
Incomplete inhibition, see text and Figures 2 and 3.
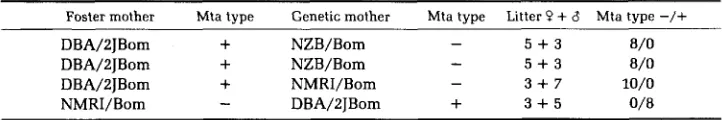
Foster-nursing: We have tested whole litters of mice delivered by hysterec-
tomy and foster-nursed by females of the opposite Mta type. Since these foster-
nursed mice retained the Mta type of their genetic mother in every case (Table
CYTOPLASMIC INHERITANCE IN MICE
489
a
I I I
15
30
60
70-
60-
50-
40-
n
b
10
20
40
Ratio competitors
:targets
FIGURE 2.-Mta typing of mice bred from wild ancestors. In one experiment (a), NZB/Bom anti- NZB/Fiill killer cells were tested on labeled BG-Tla" target cells, in another (b) (NMRI/Bom X C57BL/6)Fl anti-C57BL/G killers were tested on labeled SJL target cells. Killing in the absence of competitors is indicated by the upper horizontal lines, spontaneous release by the lower lines. The competitors were NMRI/Bom (0---0), NZB/Bom (U----U), Zhadar
(e-.),
CD(e--+),
Mil II( c - W ) , Mus spretus (A--A), and Lewis rat (0--0). The cross-hatched area in panel (a) includes BG-Tla", CBH.Q, C57BL/6 and (C57BL/6 X A/J)Fl competitors, in panel (b) BG-Tla", (DBA/ 2 X NMRI/Bom)Fl, SJL, ST/bJ and 129/01a competitors.
communication), who exchanged newborns between NZB/Icr and BALB/c
mothers.
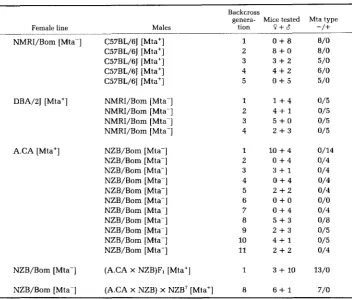
Repeated backcrossing: Apparent maternal transmission is no proof for an
extra-chromosomal mechanism. The direction of shell coiling in Limnaea
(PLAGGE
1938, cited by
GRUN
1976) and female sterility in Drosophila
(ENGELS
1979) are examples of characters with strict maternal transmission in the first
generation, but can be shown by repeated backcrossing to be determined by
chromosomal genes. We have therefore tested whether the same might not be
the case for Mta (Table
5).
490
90
m
Q)
(I)
a
Q)
-
!??
70
7
6
In
-
a
0
0
CI
-
50
8
30
60-
50-
40-
K. FISCHER LINDAHL AND B. HAUSMANN
a
I I
15
30
$0
701
b
3
0
7
15
30
60
Rat
io
competitors
:targets
FIGURE 3.-Mta typing of wild mice by cold target competition. Results of two experiments are shown, each testing NZB/Bom anti-NZB/Full killers on labeled A.CA target cells. The upper horizontal lines indicate the killing in the absence of competitors, the lower lines the spontaneous release. The cross-hatched areas in both panels show the inhibition by cells of the inbred strains A.CA, B6-Tla", BlO.DRB62, BlO,RIII(71NS), BlO.WB, C3H.JK, C57BL/6, I/LnReJ and I/St. The strongest and weakest inhibitor among four (panel a) and seven (panel b) mice caught in Denmark are shown (A---A). The other symbols refer to NMRI/Bom (0---0), Mus castaneus (W--W),
WLA 76 (A--A) and Mus molossinus (O--O).
from a standard positive strain. The Mta type was immediately reversed when
a backcross male was mated to an NZB/Bom female. Similarly, the fourth
generation of DBA/ZJ backcrossed to Mta- NMRI/Bom males (96.9% NMRI
genes) and the fifth generation of NMRI/Bom backcrossed to Mta+ C57BL/6J
males (98.4% C57BL/6 genes) remained true to their female line.
CYTOPLASMIC INHERITANCE IN MICE
491
TABLE 3Mta typing of Ft mice
Cross
(NZB/Bom x A.CA) (A.CA X NZB/Bom)
(NZB/Bom x SJL) (SJL X NZB/Bom)
(NZB/Bom X NZW/CrBom) (NZW/CrBom x NZB/Bom) (NMRI/Bom X NZB/Bom)
(NMRI/Bom x C3H.Q)
(NMRI/Bom x C57BL/6) (DBA/2 x NMRI/Bom)
Mice tested Mta type
(9 x 6 ) P + 6 -/+
-X+ 3 + 1 4/0
+X- 2 + 2 0/4
-X+ 2 + 2 4/0
+X- 2 + 2 0 /4
-X+ 5 + 0 510
+X- 5 + 0 0 / 5
-X- 3 + 1 4/0
-X+ 1 + 1 2 / 0
-X+ 1 + 1 2 / 0
+X- 2 + 0 0 / 2
TABLE 4
Mta typing of foster-nursed mice
Foster mother Mta tvpe Genetic mother Mta type Litter P
+
6 Mta tvpe-/+
5 + 3 8/0DBA/ZJBom
+
NZB/Bom -5 + 3 8/0
DBA/2JBom
+
NZB/Bom-
DBA/2JBom
+
NMRI/Bom - 3 + 7 10/0NMRI/Bom
-
DBA/ZTBom+
3 + 5 0/8DISCUSSION
Expression of Mta is determined by a maternally transmitted factor. But the
relationship between the factor and Mta expression is as yet unknown. It is
possible that the positive strains receive the structural gene coding for Mta, or
that the negative strains have a maternally transmitted repressor of a chromo-
somal, structural gene shared by all mice. Our survey of laboratory and wild
mice shows that Mta+ is the prevalent type.
Mitochondrial DNA is a useful marker of the maternal line of inheritance in
mammals (HUTCHISON
et
al.
1974; HAYASHI
et
al. 1978; GILES
et
al. 1980;
YONEKAWA
et al. 1980). FERRIS,
SAGE and WILSON
(1982) recently concluded
from a representative panel of long established inbred strains that these were
descended from a single female, whereas several different restriction enzyme
patterns were found in the mitochondrial DNA of wild mice. Their analysis
also showed that SK/CamEi had the mitochondrial DNA of SF/Cam, and that
some
Mus
molossinus samples had the mitochondrial DNA of the long estab-
lished inbred strains. The number of independent maternal lines listed in Table
492
K. FISCHER LINDAHL AND B. HAUSMANN TABLE 5Mto typing of successive generations of backcross progeny
Backcross
genera- Mice tested Mta type
Female line Males tion ? + $ -/+
~~
NMRI/Bom [Mta-] C57BL/6J [Mta+] C57BL/6J [Mta+] C57BL/6J [Mta+] C57BL/6J [Mta+] C57BL/6J [Mta+]
DBA/PJ [Mta'] NMRI/Bom [Mta-] NMRI/Bom [Mta-] NMRI/Bom [Mta-] NMRI/Bom [Mta-]
A.CA [Mta+] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-] NZB/Bom [Mta-]
1 0 + 8 8 / 0
2 8 + 0 8 / 0
3 3 + 2 5/0
4 4 + 2 6 / 0
5 0 + 5 5/0
1 1 + 4 0/5
2 4 + 1 0/5
3 5 + 0 0/5
4 2 + 3 0/5
1 1 0 + 4 0/14
2 0 + 4 0/4
3 3 + 1 0/4
4 0 + 4 0/4
5 2 + 2 0/4
6
o + o
o/o
7 0 + 4 0/4
8 5 + 3 0 / 8
9 2 + 3 0/5
10 4 + 1 0/5
11 2 + 2 0/4
NZB/Bom [Mta-] (A.CA X NZB)F, [Mta'] 1 3
+
10 13/0NZB/Bom [Mta-] ( A C A x NZB) x NZB7 [Mta+] 8 6 + 1 7/0
a Lists only mice typed by the killer assay. Since all female breeders must have had the same
type as their offspring, the total number sampled in each generation may be increased by at least two.
The stock from which the NZ mice were inbred must have contained both
Mta+ and Mta- mice. This stock originally came from a colony of obscure origin
maintained at Mill Hill, London, during the twenties
(BIELCHOWSKY
and
GOOD-
ALL1970). Unfortunately, we do not know of any other descendants from that
colony. Published records show no link between NZB and NMRI mice (FESTING
1979; MORSE
1981). It will be interesting to analyze the mitochondrial DNA of
the Mta- mice.
CYTOPLASMIC INHERITANCE IN MICE
493
T killer lymphocytes. The hypothesis requires biochemical confirmation which
will have to wait until antibodies specific for Mta become available,
The Mta type of a mouse is fixed before birth, as it cannot be changed by
foster-nursing or contact with mice of another type. Recent embryo transfer
experiments show that it is transmitted in the egg and not determined by
intrauterine influences (FISCHER
LINDAHL
and BURKI 1982). Examples of maternal
inheritance in mice are rare (reviewed by GRUN
1976
and MCLAREN
1979), and
we are not aware of any with the same strain distribution as Mta. A mitochon-
drial gene or a defective virus seem the most likely extrachromosomal genetic
vectors for Mta.
We are grateful to DRS. D. BENNETT, A. GROPP, J.-L. GUENET, P. HJORTH, J. KNIGHT, W. SCHAFER,
D.
J. SCHENDEL and H. WINKING for mice and cells, and to DR. I. M ~ L L E R NIELSEN and Ms H. HOLST at G1. Bomholtgaard for the production of foster-nursed mice. We thank DRS. M. F. W. FESTING and H. C. MORSE 111 for information and Drs. S. FAZEKAS de STCROTH and C. STEINBERG for discussions and critical reading of this manuscript. The Basel Institute for Immunology was founded and is supported by F. Hoffmann-La Roche and Co. Ltd., Basel, Switzerland.LITERATURE CITED
BIELCHOWSKY, M. and C. M. GOODALL, 1970 Origin of inbred NZ mouse strains. Cancer Res. 3 0
834-836.
ENGELS, W. R., 1979 Hybrid dysgenesis in Drosophila melanogaster: rules of inheritance of female sterility. Genet. Res. 33: 219-236.
FERRIS, S. D., R. D. SAGE, and A. C. WILSON, 1982 Evidence from mtDNA sequences that common laboratory strains of inbred mice are descended from a single female. Nature 295: 163-165. FESTING, M. F. W., 1979 Inbred Strains in Biomedical Research. MacMillan Press, London. FINNEY, D.
J.,
1964 Statistical Method in Biological Assay. Charles Griffin and Co., Ltd., London. FISCHER LINDAHL, K., M. BOCCHIERI and R. RIBLET, 1980 Maternally transmitted target antigen for FISCHER LINDAHL, K., and K. BURKI, 1982 Mta, a maternally inherited cell surface antigen of theFISCHER LINDAHL, K. and B. HAUSMANN, 1980 Qed-1-a target for unrestricted killing by T cells.
FISCHER LINDAHL, K., B. HAUSMANN and L. FLAHERTY, 1982 Polymorphism of a Qa-1-associated
FISCHER LINDAHL, K., and
1.
LANGHORNE, 1981 Medial histocompatibility antigens. Scand. J. Im-GILES, R. E., H. BLANC, H. M. CANN and D. C. WALLACE, 1980 Maternal inheritance of human
GROPP, A. and H. WINKING, 1981
unrestricted killing by NZB T lymphocytes. J. Exp. Med. 152: 1583-1595.
mouse, is transmitted in the egg. Proc. Natl. Acad. Sci. USA
79:
5362-5366. Eur. 1. Immunol. 10: 289-298.antigen defined by cytotoxic T cells. I. Qed-1" and Qed-ld. Eur. J. Immunol. 1 2 159-166.
munol. 14: 643-654.
mitochondrial DNA. Proc. Natl. Acad. Sci. USA 7 7 6715-6719.
patterns and biological consequences of heterozygosity. Symp. 2001. Soc. Lond. 47: 141-181. Robertsonian translocations: cytology, meiosis, segregation
GRUN, P., 1976
HAYASHI, ].-I., H. YONEKAWA, 0. GOTOH, J. WATANABE and Y. TAGASHIRA, 1978
HUTCHISON C. A., 111, J. E. NEWBOLD, S . S. POTTER and M. H. EDGELL, 1974
Cytoplasmic Genetics and Evolution. Columbia University Press, New York.
inheritance of rat mitochondrial DNA. Biochem. Biophys. Res. Commun. 83: 1032-1038. Strictly maternal
494
K. FISCHER LINDAHL A N D B. H A U S M A N NKOHLER, G., K. FISCHER LINDAHL and C. HEUSSER, 1981 Characterization of a monoclonal anti- H-2Kh antibody. pp. 202-208. In: The Immune System, Vol. 2, Edited by
c.
M. STEINBERG and I. LEFKOVITS. Karger, Basel.Hybrid cell lines secreting monoclonal antibody specific for major histocompatibility antigens of the mouse. Nature 271:
LEMKE, H., G. J. HAMMERLING, C. HOHMANN and K. RAJEWSKY, 1978
249-251.
LYNCH, C. J., 1969 The so-called Swiss mouse. Lab. Anim. Care 1 9 214-220.
MCLAREN, A., 1979 The impact of pre-fertilization events on post-fertilization development in mammals. pp. 287-320. In: Moternal effects in development, Edited by
D.
R. NEWTH and M. BALLS. Cambridge University Press, Cambridge.NZB x BALB/c recombinant inbred lines. Mouse News Letter 64: 46- 47.
The laboratory mouse-a historical perspective. pp. 1-16. In: The Mouse in Biomedical Research, Vol. 1, Edited by H. L. FOSTER, J.
D.
SMALL and J. G. Fox. Academic Press, New York.Perspect. Virol. 6: 75-87.
In: Origins of Inbred Mice. Edited by H. C. MORSE 111. Academic Press, New York. MOLLER NIELSEN, I., 1981
MORSE, H. C., 111, 1981
MUHLBOCK, 0. and P. BENTVELZEN, 1968 The transmission of the mammary tumor viruses.
Comments on the relationship of inbred strains to the genus Mus. pp. 497-509.
Origins of SF/Cam strain. Mousc News Letter 64: 73. POTTER, M., 1978
SAGE, R. D., 1981
SCHREIER, M. H. and R. TEES, 1981 Long-term culture and cloning of specific helper T cells. pp. 263-275. In: Immunological Methods, vol 11. Edited by 1. LEFKOVITS and B. PERNIS. Academic Press, New York.
WALLACE, M. E., 1971 An unprecedented number of mutants in a colony of wild mice. Environ.
WALLACE, M. E., 1981 The breeding, inbreeding and management of wild mice. Symp. Zool. Soc.
WINKING, H., K. NIELSEN and A. GROPP, 1980 Variable positions of NORs in Mus musculus. Cytogenet. Cell Genet. 26 158-164.
YONEKAWA, H., K. MORIWAKI, 0. GOTOH, J. WATANABE. J.-I. HAYASHI, N. MIYASHITA, M. L. PETRAS and Y. TAGASHIRA, 1980 Relationship between laboratory mice and thc subspecies Mus musculus domesticus based on restriction endonuclease cleavage patterns of mitochondrial DNA. Jpn. J. Genet. 55: 289-296.
Pollut. 1: 175-184.
Lond. 47: 183-204.