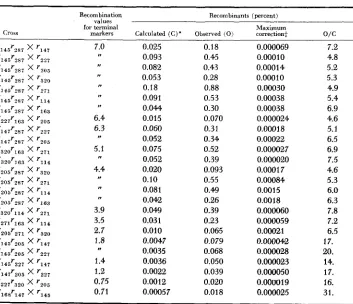
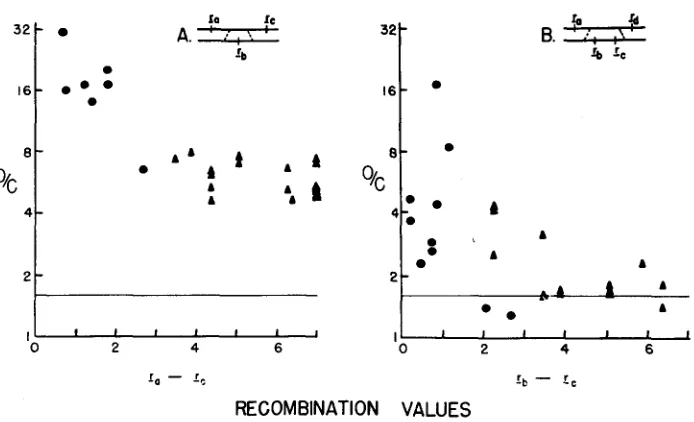
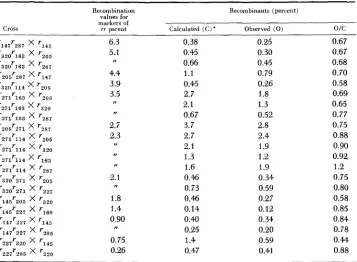
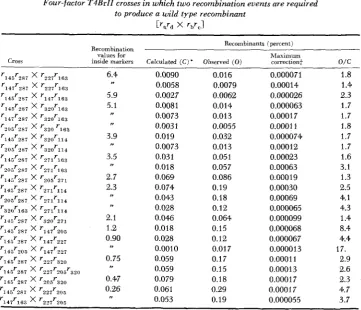
HIGH NEGATIVE INTERFERENCE OVER SHORT SEGMENTS OF THE GENETIC STRUCTURE OF BACTERIOPHAGE T4
22
0
0
Full text
Figure


+7
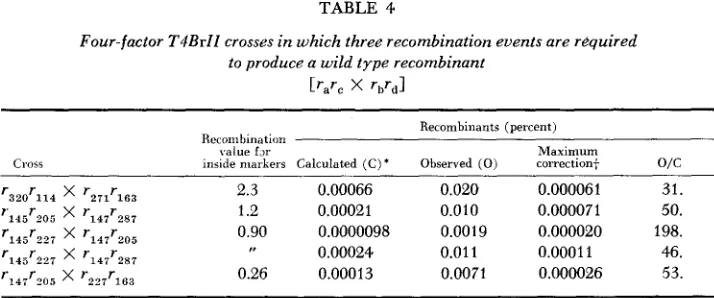
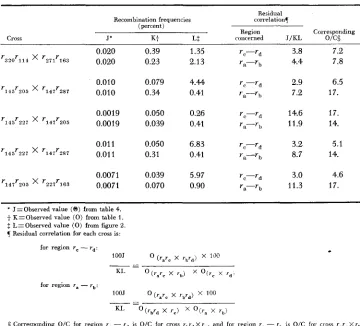
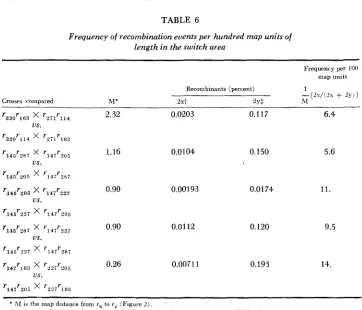
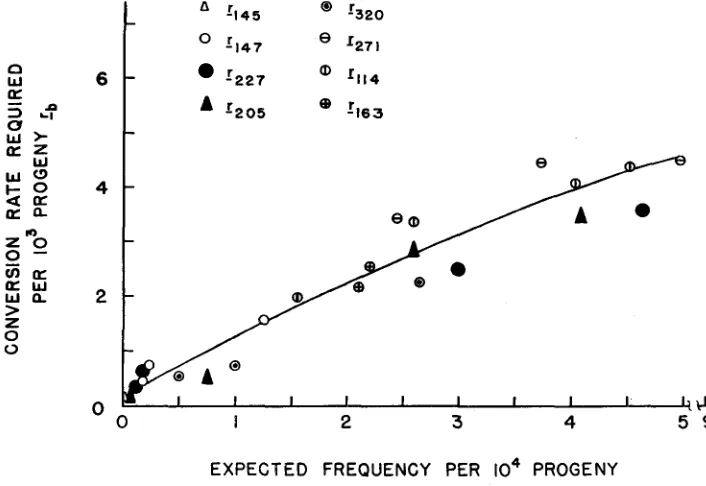
Related documents