QTL Mapping of Flavonoid and Phenolic Acid Variation in Broccoli
160
0
0
Full text
(2)
(3)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(29)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
(39)
(40)
Figure

+7
Related documents
The highest content of total hydroxybenzoic acids, total hydroxycinnamic acids, total flavan-3-ols, total flavones and flavonols and total stilbenes was observed in the skin of
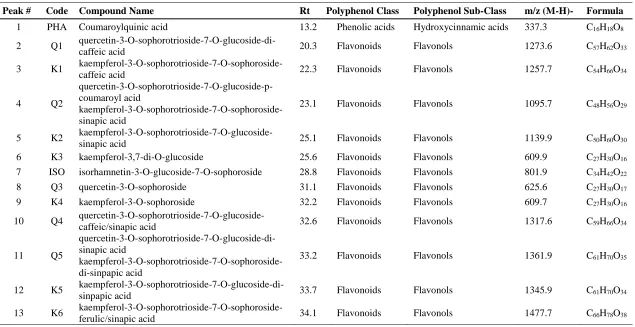
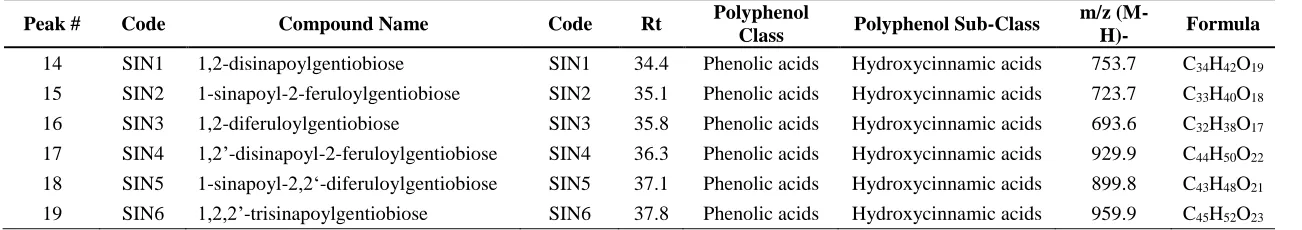
In ad- dition, hydroxycinnamic acid derivatives were also detected in the studied samples, being synapoylmalic acid the major one in the class of phenolic acids (2.8 ± 0.1 and 1.29
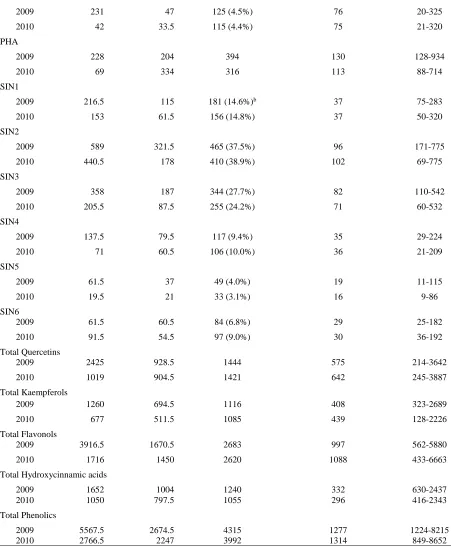
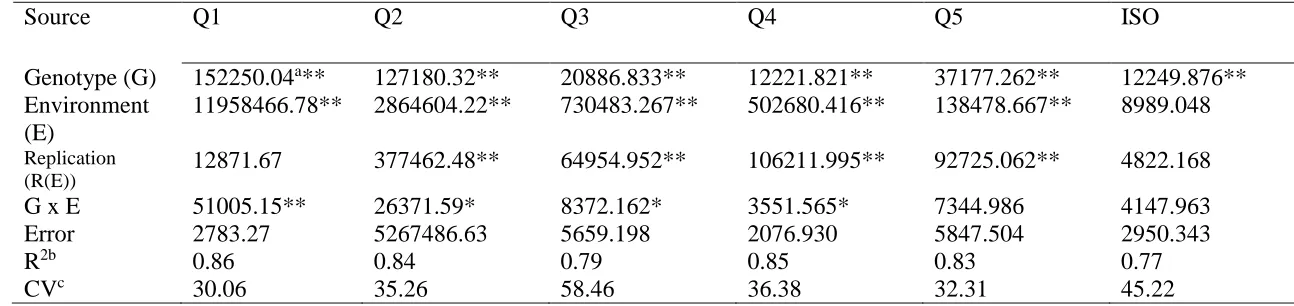
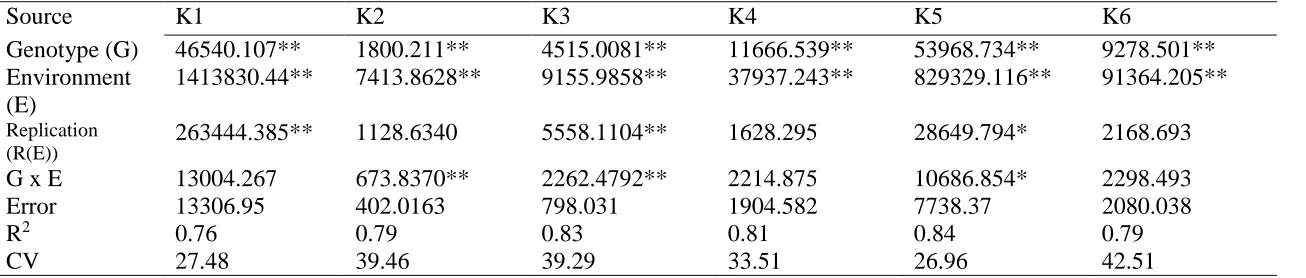
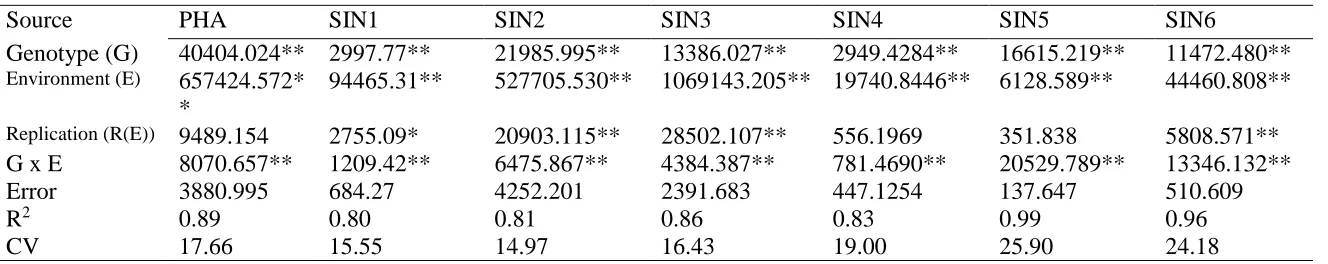
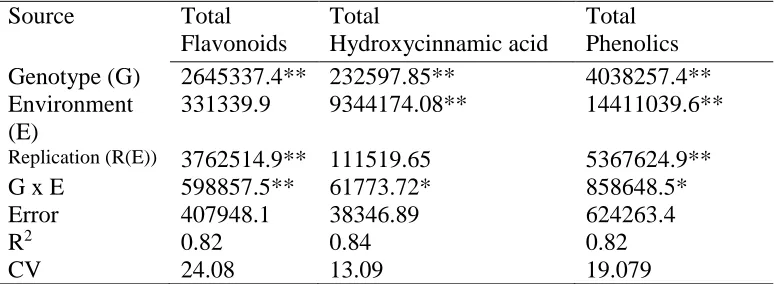
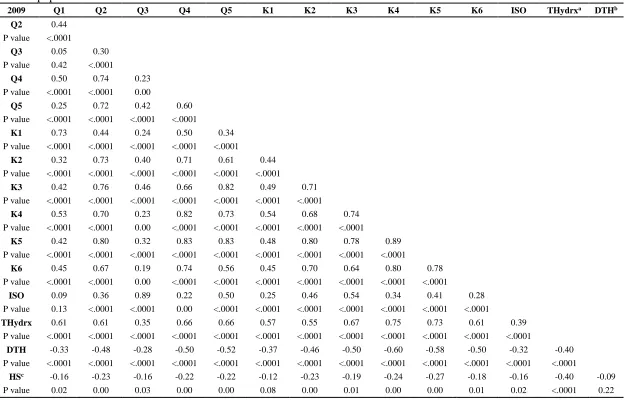
Characterization of the variation in total phenolic content and antioxidant capacity in the VI-158 × BNC broccoli mapping population corroborated the expectation that both genetic and
The major phenolic compounds present in different fruit peels (apple, pomegranates, mango, pineapple, and citrus peels) include hydroxybenzoic and hydroxycinnamic acids (caffeic
The greatest delay in chlorophyll degradation and florets senescence in broccoli during 0°C was found with 1.5 % ascorbic acid treatments that was followed the lowest
Our objectives were to (1) determine the total antioxidant activity, phenolic, flavonoid and ascorbic acid contents of commonly consumed vegetables in Nigeria and
The highest total flavonoid content (193.3 µg quercetin equivalent/g dry weight), total phenolic content (717.8 µg gallic acid equivalent/g dry weight) and antioxidant activity
In estimation of phenolic content, by comparing total phenolic content of both permeate and retentate, permeate shows higher phenolic content in two different MWCO
