Copyright 6 1996 by the Genetics Society of America
The Large Subunit of Replication Factor C (Rfclp/Cdc44p) Is Required
for
DNA Replication and DNA Repair
in
Saccharomyces cerevkiae
Michael A. McAlear,’
K
Michelle
Tuff0
and
Connie
Holm
Department of Pharmacology, Division of Cellular and Molecular Medicine, University of California at San Diego, L a Jolla, California 92093-0651
Manuscript received August 3, 1995 Accepted for publication October 2, 1995
ABSTRACT
We used genetic and biochemical techniques to characterize the phenotypes associated with mutations affecting the large subunit of replication factor C (Cdc44p or Rfclp) in Saccharomyces cermisim. We demonstrate that Cdc44p is required for both DNA replication and DNA repair in vivo. Cold-sensitive cdc44 mutants experience a delay in traversing S phase at the restrictive temperature following alpha factor arrest; although mutant cells eventually accumulate with a G2/M DNA content, they undergo a cell cycle arrest and initiate neither mitosis nor a new round of DNA synthesis. cdc44 mutants also exhibit an elevated level of spontaneous mutation, and they are sensitive both to the DNA damaging agent methylmethane sulfonate and to exposure to UV radiation. After exposure to UV radiation, cdc44 mutants
at the restrictive temperature contain higher levels of single-stranded DNA breaks than do wild-type cells. This observation is consistent with the hypothesis that Cdc44p is involved in repairing gaps in the DNA after the excision of damaged bases. Thus, Cdc44p plays an important role in both DNA replication and DNA repair in vivo.
G
ENETIC and biochemical studies have revealed an underlying conservation in the basic enzymology of DNA replication and DNA repair in a variety of pro- karyotic and eukaryotic systems (reviewed in CARR and HOEKSTRA 1995; STILLMAN 1994). Among the central proteins that are required for these processes are the DNA polymerases and their accessory proteins. Two classes of DNA polymerase accessory proteins that have been identified are factors known as “sliding DNA clamps” and the “clamp loaders” (O’DONNELL et al.1993). These types of proteins constitute part of a widely conserved mechanism for enhancing the efficiency of DNA replication. The sliding DNA clamp class of pro- teins includes the
p
subunit of DNA polymerase I11 in Eschm’chia coli and proliferating cell nuclear antigen (PCNA) in eukaryotes. Although unrelated in amino acid sequence, these two proteins share striking func- tional and structural similarities (KONG et al. 1992;KRISHNA et al. 1994). In each case, the proteins assemble as multimers to form doughnut-shaped rings that can encircle the DNA double helix, tethering the DNA poly- merase to the DNA template.
Central players in DNA replication in vitro are the clamp loaders, which may play an important role in switching DNA synthesis from DNA polymerase a to
Corresponding authur: Connie Holm, Department of Pharmacology, Division of Cellular and Molecular Medicine, University of California at San Diego, Mail Code 0651, 9500 Gilman Dr., La Jolla, CA 92093- 0651. E-mail: [email protected]
‘Present address: Department of Molecular Biology and Biochemis- try, Wesleyan University, Middletown, CT 06459.
Genetics 142 65-78 (January, 1996)
DNA polymerase 6 (TSURIMOTO et al. 1990; EKI et al.
1992). Clamp loaders serve to open the ring-like sliding clamps and assemble them around the double helix (O’DONNELL et al. 1993). Some examples of clamp load- ers include the gp44/62 proteins from bacteriophage T4, the gamma complex in E. coli, and replication factor C (RF-C) in eukaryotic cells (O’DONNELL et al. 1993). In each case, the clamp loader is a multiprotein com- plex that has several characteristic activities in vitro. RF- C, for example, is a multisubunit DNA-binding protein that interacts with the 3‘-OH terminus at primer-tem- plate junctions (LEE et al. 1988; YODER and BURGERS
1991; FIEN and STILLMAN 1992; PODUST et al. 1995). In addition to binding DNA, RF-C also associates with PCNA, and it can load PCNA onto the double helix in an ATP-dependent reaction (LEE and HURWITZ 1990;
TSURIMOTO and STILLMAN 1991). Although biochemi- cal evidence suggests that this reaction is important for processive DNA replication in vitro, the role that these proteins play in DNA replication and DNA repair in
vivo remains to be determined.
66 M. A. McAlear, K M. Tuffo and C. Holm
to containing the conserved RF-C box, it also includes an additional 330-amino acid Gterminal domain and a 300-amino acid N-terminal domain, part of which has homology to sequences found in prokaryotic DNA li- gases. Thus, the large subunit is likely to play a distinct role in contributing to RF-C function. Although studies on both human and mouse RF-C indicate that the large subunit can bind to DNA on its own in vitro (TSURIMOTO
and STILLMAN 1991; BURBELO et al. 1993), the precise role that this protein plays in DNA metabolism in vivo is not clear.
In this study, we demonstrate that the large subunit of RF-C (Cdc44p or Rfclp) is required for both DNA replication and DNA repair in vivo. When cdc44 mutants are released from an alpha factor arrest at the restrictive temperature, they progress through S phase very slowly, and they ultimately arrest with a G2/M DNA content before the initiation of mitosis. cdc44 mutants also ex- hibit a mutator phenotype in vivo, and they are sensitive to MMS and W irradiation. In addition, we demon- strate that after irradiation with UV light, cdc44 mutants have a higher level of single-stranded DNA breaks than do CDC44 cells. Taken together, these results suggest that Cdc44p plays an essential role in the repair of single-stranded breaks in DNA.
MATERIALS AND METHODS
S t r a i n s and media: The yeast strains used in this study are listed in Table 1. Standard genetic techniques were used in constructing the strains (SHERMAN et al. 1986), and the con- struction of all strains created by integration of plasmid DNA was verified by Southern analysis (SOUTHERN 1975). To create rho" isolates of strains, rho+ strains were first inoculated into liquid W D medium containing 25 yg/ml of ethidium bro- mide and incubated for 2 days at 30" (SHERMAN et ul. 1986). The cultures were then streaked onto W D plates and incu- bated at 30" for a further 3 days. Single colonies were recov- ered from the plates and tested for their ability to grow on YEP medium with glycerol (2%) as a carbon source. Rho- colonies were tested for the presence of mitochondrial DNA by staining the cells with 4',6-diamidino-2-phenylindole and examining the cells by fluorescence microscopy.
Strains CH2213 and CH2214 were constructed by trans- forming strain MKF"o with a DNA fragment containing either the WC44 or cdc44-I gene that was linked to a DNA fragment containing the HIS3 gene. Briefly, for the construction of CH2213, a 1.8-kb BamHI fragment of the HZS3 gene derived from plasmid pJJ215 (JONES and PRAKA~H 1990) was inserted into the SnaBI site downstream of the WC44 gene in plasmid pCH1335 (HOWELL et al. 1994). This created plasmid pCH1541, which contained the HZS3 gene attached down- stream of the CDC44 gene. A 4.8-kb StuI-KpnI WC44.HZS3 fragment from pCH1541 was then used to transform the his3 strain MKF"o, creating strain CH2213 (WC44.HIS3). An iden- tical strategy using the cdc44-I plasmid pCH1350 was used to
create strain CH2214 (cdc44-1.HZS3).
Plasmids pCH1587 (PGALI-WC44) and pCH1588 (pGAL1- cdc44-I) were created by cloning a Fnu4HI-SnuBI WC44 frag- ment from plasmids pCH1335 (WC44) and pCH1350 (cdc44-
I ) into the SmuI site of plasmid pCH1155 (PGALI)
(KRANz
1993). A NotI fragment from pCH1587 or pCH1588 was
cloned into the NotI site of plasmid pCH1099 (URA3) to cre- ate plasmids pCH1523 (PGALI-WC44 URA3) and pCH1524 (pGAL1-cdc44-I URA3). These plasmids were transformed into strain CH1587 (ura3-52 trpl WC44::LEU2 [pCH1289 ( W C 4 4 TRPl)] 1, and the resulting strains were streaked onto SD plus Trp medium. Trp- isolates that had lost the plasmid pCH1289 (WC44 TRPl) were recovered, creating strains CH2088 { WC44::LEUZ uru3-52 trpl [pCH1523(pGALl-WC44 URA3)]] and CH2089 (CDC44::LEUZ ura3-52 trpl [pCH1524(pGAZ,l-
YEPD medium is 1% yeast extract, 2% bacto-peptone and 2% glucose. YEP Raf/Gal medium is 1% yeast extract, 2% bacto-peptone, 2% galactose and 2% raffinose. SD medium is 6.7 g/I yeast nitrogen base without amino acids (Difco), 20 g/l of glucose, and 20 g/1 of bacto-agar (Difco). SC medium is SD medium supplemented with 20 mg each of adenine, histidine, and uracil, 60 mg of leucine, 30 mg of lysine, and 20 mg of tryptophan per liter of medium.
Molecular biology: All standard techniques of molecular biology were performed as described (AUSUBEL et al. 1988). Creation of new cdc44 alleles: New mutant alleles of the CDC44 gene were created by random or sitedirected muta- genesis. For random mutagenesis, the plasmids pCH1530 (WC44.LEU2 URA3) and pCH1531 (CDC44.LEU2 URA3) were propagated in the mutagenic mutD E. colistrain KD1067 (arg his mutD5 Sut) (HENKIN and SONENSHEIN 1987). Muta- genized plasmids were recovered from E. coli and then trans- formed into the yeast strain CH1587. The plasmid shuffle technique (BUDD and CAMPBELL 1987) was used to select for strains bearing only the mutagenized plasmids, and these strains were screened for mutant phenotypes. Sitedirected mutations in the WC44 gene were constructed by the method of KUNKEL et al. (1987). Mutant plasmids were recovered from
E. coli and sequenced to confirm the presence of the muta- tions, and the mutant WC44 fragments were subcloned into plasmid pCH1335. The new mutant WC44 alleles were trans- formed into yeast strains bearing a deletion in the WC44 gene by a plasmid shuffle method (BUDD and CAMPBELL 1987). Transformants were screened for phenotypes including heat and cold sensitivity, and sensitivity to MMS and hydroxyurea. GST-Cdc44p fusion proteins and GTP binding assays: To purify Cdc44p for GTP binding assays, we created GST- Cdc44p fusion constructs and purified the fusion proteins with glutathione agarose beads. Briefly, a 3.4kb BamHI-EcoN WC44 fragment from plasmid pCHl335 (CDC44) was cloned into plasmid pGEX-1X (SMITH and JOHNSON 1988) that was digested with BamHI and EcoN. This created plasmid pCH1484 (GST-WC44). A similar strategy using plasmid pCH1350 (cdc44-1) was used to create plasmid pCHI499 (GST- cdc44-I). In these constructs, the nonessential N-terminal 140 amino acids of the WC44 gene (HOWELL et ul. 1994) were replaced with sequences encoding the GST protein. Plasmids were transformed into E. coli and grown overnight, and ex- pression of the fusion proteins was induced with 1 mM isopro- pyl thiogalactoside for 3 hr. Cells were collected, digested with lysozyme (0.2 mg/ml), and lysed by sonication, and the lysates were incubated with glutathione agarose beads (Sigma). The glutathione beads were washed with buffer A (phosphate-buffered saline with 20% glycerol, 1% NP-40, 1 mM EDTA, 1 mM 1,4dithiothreitol, 1 pg/ml leupeptin, 1 yg/ ml pepstatin and 1 mM phenylmethyl sulfate fluoride) to re- move unbound proteins. The presence of the fusion proteins was confirmed by SDSPAGE. Equal amounts of protein were then incubated with alpha jnP GTP (NEN duPont) for 10 min at 30" in buffer B (50 mM Tris pH 7.5,50 mM KCI, 5% glycerol and 0.5 mM MgCI2). After incubation, the beads were washed
RFC in DNA Replication and Repair
TABLE 1
S. cereuisiQe strains used in this study
67
Strain Genotype" Reference
CH325 CH526 CH533 CH541 CH545 CH756 CH1491 CH 1492 CHI 494 CH1587 CH1589 CH1591 CH1634 CH1806 CH1807 CH2027 CH2088 CH2089 CH2211 CH2212 CH22 13
CH2214
CH22 15 MKP-O
MATa lys2-801 ura3-52 his4-539 top2-4
MATa ade 1 ade2 lys2 ural his7 tyrl gall cdc2-lb MATa ade 1 ade2 lys2 ural his7 tyrl gall cdc9-I MATa ade I ade2 lys2 ural gall cdcl7-1
MATa ade 1 ade2 lys2 ural his7 tyrl gall cdc21-lb MATa ade 1 ade2 lys2 ural his7 tyrl cdc8-1 MATa leu2-3, I 1 2 ura3-52 cdc44-1 MATa ade2 leu2-3,112 ura3-52 MATa ade2 leu2-3,112 ura3-52 cdc44-I
MATa ura3-52 leu2AI trplA63 W44::LEUZ [pCH1289 ( W C 4 4 TRPI)] MATa ura3-52 bu2AI trplA63 W44::LEU2 [pCH1297 (cdc44-2 TRPI)] MATa ura3-52 leu2Al trplA63 W44::LEU2 [pCH1289 (cdc44-4 TRPI)] MATa his4 lys2 ura3-52 rad52::URA3
MATa ura3-52 his4-539 lys2-801 W C 4 4 . URAP MATa ura3-52 his4-539 lys2-801 cdc44-1. URAP MATa ura3-52 leu2 rad9::LEU2
MATa ura3-52 leu2Al trplA63 WC44::LEUZ [pCH1523 (pGAL1-WC44 URA3)] MATa ura3-52 leu2Al trplA63 WC44::LEU2 [pCH1523 (PGALI-cdc44-1 URA3)] MATa ade2 leu2-3,112 ura3-52 W C 4 4 [rho"]
MATa ade2 leu2-3,112 ura3-52 cdc44-I [rho"]
MATa canl-100 ade2-1 lys2-1 ura3-52 leu2-3,112 his3-A200 trpl-A901 WC44.HIS3
MATa canl-100 ade2-l lys2-1 ura3-52 leu2-3,112 his3-A200 trpl-A901 cdc44-l.HIS3
MATa ade2 ura3-52 cdc9 rho"
MATa canl-100 ade2-1 lys2-1 ura3-52 leu2-3,112 his3-A200 trpl-A901 IJ'CpMP21 wCpMP2 SUP4-01
wCpMP2 SUP4-01
HOLM et al. (1985) HARTWELL et al. (1973) HARTWELL et al. (1973) HARTWELL et al. (1973) HARTWELL et al. (1973) HARTWELL et al. (1973) M&.EAR et al. (1994) MCALEAR et al. (1994) This study
HOWELL et al. (1994) This study
This study SPELL et al. (1994) HOWELL et al. (1994) HOWELL et al. (1994) This study
This study This study This study This study This study
This study
KUNZ et al. (1989)
a Extrachromosomal elements and plasmids indicated by [ 1.
'This strain has URA3 integrated next to W C 4 4 or cdc44-1 at the W C 4 4 locus. This strain has additional unidentified auxotrophies.
three times in buffer B to remove any unbound GTP and then counted in a scintillation counter.
Synthetic lethality: Cold-sensitive strain CH1491 (cdc44-1) was crossed with each of the heat-sensitive strains CH526
CH545 (cdc21-I) and CH325 (top2-4). Diploid strains were selected, grown on rich medium, and sporulated. Tetrads from each cross were dissected, and the resulting spores were allowed to germinate on YEPD at 25". This temperature was permissive for both the cold-sensitive (Cs-) and heat-sensitive (Ts-) strains. The viability of the spores and the segregation of markers were determined for 2 1 4 tetrads from each cross. We were able to recover at least eight double mutant (Cs- Ts-) spores from each of the diploids that were sporulated, except for the diploid strain that was derived from the CH1491 (cdc44) X CH526 (cdc2) cross. In that case, we recovered zero out of an expected 15 double mutant (cdc44 cdc2) spores.
Detection of spontaneous SUP& mutants: Spontaneous mutagenesis assays were performed essentially as described previously (KUNZ et al. 1989). We used isogenic strains CH2213 ( W C 4 4 ) and CH2214 (cdc44-1) that carried chromo- somal canl-100, ade2-1, lys2-I and ura3-52 alleles. The canl- 100, ade2-1, lys2-1 alleles were suppressed by a SUP4-0 ochre suppressor tRNA gene that was present on an extrachromo- somal YCp URA3, SUP4-o plasmid. Cells harboring the SUP4- o plasmid were therefore Can', Ade+, Lysf, and Ura+. Cells acquiring a mutation in the SUP4-0 gene become Can', Ade-, Lys-, and Ura'. Strains CH2213 and CH2214 were streaked for single colonies onto SC "ura plates to maintain the URA3
( c d ~ 2 - l ) ~ CH756 ( c d c a - l ) , CH533 (cdc9-l), CH541 (cdcl7-1),
sUP4-o plasmid and then incubated at 30" for 4 days. Indepen- dent colonies from each strain were used to inoculate 5 m l cultures of liquid SC "ura media, and these cultures were incubated overnight at 30". Cells were sonicated, counted, diluted as necessary in water, and plated onto SC "ura, SC "ura plus canavanine (30 mg/ml), and YPD plates. Plates were incubated at 30" for 5 days and then scored for colony growth. For each strain, the data from 15 independent cul- tures were used to determine the mean mutation frequencies. Red (ade-) canavanine-resistant colonies were tested for ly- sine auxotrophy, and Can', Ade-, Lys- clones were retained for sequencing analysis. For sequencing, PCR was used to amplify a DNA fragment containing the SUP4-0 gene directly from yeast cells. The primers used for this amplification flanked the SW4-o gene and were of the following sequence: primer 1,5'-GTGGCGCCGGTGATGCCGGG3'; primer 2,5'- GTTGTGTGGAATTGTGAGCG-3'. The amplified DNA prod- uct was used as a substrate for sequencing by the fmol'" DNA sequencing system (Promega).
Microfluorometric analysis: Strains CH1806 ( W C 4 4 ) and CH1807 (cdc44-1) were grown to early log phase in liquid W D p H 4 at 30" and then arrested with alpha factor (5 pg/
68 M. A. McAlear, K. M. Tuffo and C. Holm
(HUTTER and EIPEL 1979). Each DNA content histogram was based on the analysis of 210,000 cells.
Reciprocal shift experiments: Strains CH1806 ( W C 4 4 ) and CH1807 (cdc44-1) were grown to early log phase in liquid YPD medium at 30" and then treated as indicated. For control experiments, a sample of the 30" culture was plated onto 14" YEPD plates or 3O"YEPD plates containing 0.3 M hydroxyurea. Although control cultures were also plated onto YEPD plates containing 80 pg/ml benomyl, only about half of the cells arrested as doublets, making the results of this shift impossible to interpret; results of shifts to benomyl plates are therefore not reported here. For the experimental cultures, cells were first incubated in YPD containing HU or nocodazole at 30" for 3 hr, or in YPD alone at 14" for 12 hr. The cultures were then chilled, washed, sonicated, and plated onto YEPD plates at 14" or onto 30" YEPD plates containing HU. Cell morphol- ogy was scored immediately by light microscopy (zero time) and then again after either 5 hr at 30" or 12 hr at 14".
Sensitivity to DNA damaging agents: For assaying sensitivity to DNA damaging agents, yeast strains were first grown in liquid YPD medium at 30" to early log phase, sonicated, and diluted in water as necessary. For assaying UV sensitivity, the cells were plated onto YPD plates and exposed to u p to a maximum of 200 J/m' of 254 nm UV radiation from a model 1800 Stratalinker (Stratagene). For assaying MMS sensitivity, cells were incubated in 0.3% MMS for up to 40 min and then plated onto YPD plates. For assaying sensitivity to gamma rays, cells were diluted in water, exposed to u to a maximum of 20 krads of gamma radiation from a Cs'
'
source, and then plated onto YPD plates. Plates were incubated at 30" for 3 days and then scored for viability. The plots represent the average viabilities determined from two independent experi- ments.Velocity sedimentation gradients: Cell culture and chro- mosomal DNA isolation was performed by a modification of a previously published protocol (BUDD et al. 1989). Briefly, strains CH2211 [WC44(rhoo)] and CH2212 [cdc44(rhoo)] were grown in liquid SC at 30" to a density of -5 X
lo6
cells/ ml. Sixty millicuries of 5,G'H uracil (35 Ci/mmol) (NEN Dupont) or 10 mCi of I4C uracil (60 mCi/mmol) (NEN Du- pont) were added to 500 p1 aliquots of cells, and aliquots were incubated for 6-8 hr at 30". Where indicated, cultures were irradiated with 100 J/m' W radiation or shifted to 14". (At 100 J/m' of UV irradiation, the viability of the cDC44 and cdc44 cells was 80 and 30%, respectively.) After incubation, the cells were pelleted, resuspended in 0.5 ml of cold 0.1 M Tris pH 8.5, 0.1 M EDTA, 2% 2-mercaptoethanol, and incubated on ice for 15 min. Cells were washed, resuspended in cold 0.01 M KP04 pH 9.72, 0.01 M EDTA and then transferred to the bottom of a 5-ml polyallomer centrifuge tube (Beckman). Twenty microliters of 10% zymolyase and 10 pl of RNAse A (10 mg/ml) were added to the cells, and the samples were incubated at 37" for 15 min. Nonidet P40 and sarkosyl were added to a final concentration of 2%, and for the alkaline gradients, NaOH was added to 0.5 M. A 4 m l 15-30% sucrose gradient with 0.7 M NaCI, 0.03 M EDTA (and 0.3 M NaOH for alkaline gradients) was pumped into the bottom of the tube. Alkaline gradients were centrifuged for 17 hr at 12,000 rpm in a Beckman SW55 rotor at 20"; neutral gradients were centrifuged at 12,000 rpm for 13 hr. After centrifugation, the bottoms of the tubes were punctured, and 20 200-pl fractions were collected. For the neutral gradients, residual RNA was hydrolyzed by incubation overnight with the addition ofNaOH to 0.5 M. Refractive indices of each fraction were deter- mined to confirm the integrity of each gradient. Aliquots (150 pl) from each fraction were precipitated with 3 ml of 5% trichloroacetic acid, 0.1 M Na pyrophosphate. Precipitates
r
were collected onto GF/C filters (Whatman), and the filters were washed with 25 ml 5% trichloroacetic acid, 0.1 M Na pyrophosphate, and 10 ml ethanol. Filters were counted for H and 14C in 5 ml Ecoscint (National Diagnostics) for 5 min each.
To eliminate the contributions of mitochondrial DNA from this type of analysis, we used rho" isolates from each strain tested (SHERMAN et al. 1986). To facilitate the comparison between the profiles of different strains, two chromosomal DNA preps (ie., one from W C 4 4 cells labeled with 'H uracil and one from cdc44 cells labeled with 14C uracil) were run together within the same gradient. The resulting profiles were confirmed by repeating the experiments with the strains la- beled in the opposite orientation. The profiles are plotted with fraction 1 representing the top of the gradient and frac- tion 20 representing the bottom of the gradient. DNA from bacteriophage T4 was sedimented in parallel gradients and used as a molecular weight marker. Under neutral gradients, T4 DNA migrated to fraction 5. Under alkaline conditions, T4 DNA migrated to fraction 6.
To calculate the number-average molecular weight (M,) of the gradient profiles, we used the formula (GREEN et al. 1974; BUDD et al. 1989)
M, = - C,
C,
x-
t Mt
where C, is the activity in the ith fraction. M, is computed by the equation (FRIEFELDER 1970)
MZ/M*4 = ( d,/ d ~ 4 ) ''69,
where d, is the distance of the ith fraction and h4 is the distance sedimented by T4 DNA (MT4 = 160 kb). Since the very top of the gradients do not resolve DNA fragments well, the contributions of the first three fractions from each profile were omitted from the calculation of the number-average mc- lecular weight (PETES and FANCMAN 1972).
RESULTS
Genetics of cdc44 mutations: To characterize essential domains and activities of the large subunit of RF-C, we used a genetic approach to isolate and characterize
RFC in DNA Replication and Repair 69
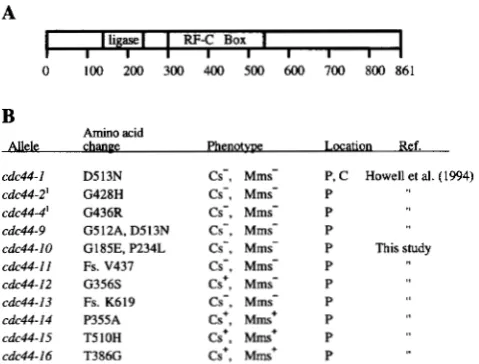
A
I
IligaseI I RF-C Box II I I I I I I I
0 100 200 300 400 500 600 700 800 861
B
Amino acid
Allele chanee Phenotwe Location Ref. cdc44-1 D513N Cs-, Mms- P, C Howell et al. (1994) cdc44-2' G428H Cs-, Mms- P
cdc44-4' G436R Cs-, Mms- P cdc44-9 G512A, D513N Cs-, Mms- P
cdc44-10 G185E. P234L Cs-, Mms- P This study cdc44-11 Fs. V437 Cs-, Mms- P
cdc44-12 G356S Cs', Mms- P cdc44-13 Fs. K619 Cs-, Mms- P cdc44-14 P355A Cs', Mms* P
cdc44-15 T510H Cs*, Mms' P cdc44-16 T386G Cs', Mms' P
FIGURE 1.-Mutant W C 4 4 alleles. (A) Map of the CDC44 gene indicating the regions of homology to the smaller sub-
units of replication factor C (RF-C box) and to E. coli DNA ligase (ligase). (B) List of cdc44 alleles and their properties.
Fs., frameshift mutation; Cs-, cold sensitive; Mms-, methyl-
methane sulfonate sensitive; P, phenotype detected in a strain carrying a plasmid-borne mutation; C, phenotype detected in a strain carrying a chromosomal allele; I , mutations in trun-
cated alleles.
We also tested the significance of a putative nucleo- tide-binding domain that contains sequence motifs that are similar to GTP-binding consensus sequences. It had appeared that these motifs might be significant, be- cause three previously isolated cdc44 alleles contained mutations in or near the putative consensus sequences. In particular, a p2lRAS mutation analogous to the cold sensitive cdc44-1 mutation decreases p21- binding of guanine nucleotides by 100-fold (CHANNING et al. 1986). First, we tested Cdc44p for GTP binding directly. We expressed GST-tagged Cdc44p in E. coli, purified the recombinant protein, and assayed it for GTP binding in vitro. Although we could detect GTP binding in a GST-p21RAS control protein, we could not detect GTP binding in either GST-Cdc44p or GST-Cdc44lp fusions (data not shown). Second, we constructed site-directed mutations within the putative GTP-binding domain and tested the resulting alleles for mutant phenotypes. We created three new cdc44 alleles, each of which was based on a known mutation that disrupts the activity of the GTP-binding protein ~ 2 1 ~ . These alleles included mu- tations in the putative G1 region (P355A), G2 region
(T386G) and G4 region (T510H) of Cdc44p. The anal-
ogous mutations in p2lW affect GTP hydrolysis, ~ 2 1 ~ activation, and nucleotide binding, respectively (VALEN-
CIA et al. 1991). Although we looked for sensitivities to heat, cold, hydroxyurea, MMS and UV irradiation, none of these alleles gave rise to any detectable mutant phe- notype. Taken together, these results suggest that it is unlikely that the essential activity of Cdc44p requires GTP-binding.
Finally, we used synthetic lethality to identify proteins
involved in DNA metabolism with which Cdc44p might interact in vivo. We crossed a cdc44 mutant strain with several mutants defective in DNA metabolism. We found that cdc44-1 exhibited synthetic lethality with cdc2-1 (DNA polymerase
6)
but not with cdc8-1 (thymi- dylate kinase), cdc9-l (DNA ligase), cdcl7-1 (DNA poly- merase a), cdc21-1 (thymidylate synthase) or t@2-4 (DNA topoisomerase 11). Although other interpreta- tions are possible, these results are consistent with the hypothesis that Cdc44p interacts directly with DNA polymerase6
in vivo, and they are consistent with the biochemical evidence that RF-C interacts with DNA polymerase6
in vitro (TSURIMOTO and STILLMAN 1989;BURGERS 1991).
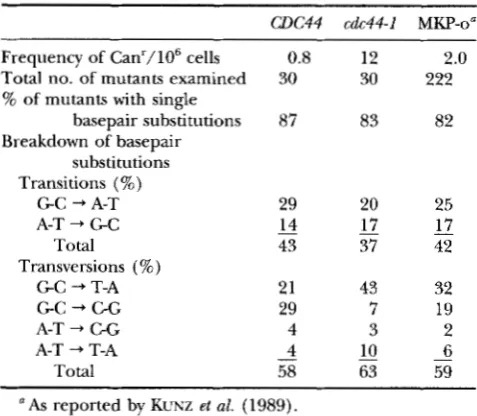
cdc44 mutants exhibit a nonspecific elevation in muta- tion frequency: Although cdc44 mutants have an ele- vated mutation rate (MCALEAR et al. 1994), it was not clear whether this mutator phenotype is of a general nature, or if cdc44 mutants have increased levels of a specific type of DNA mutation. To address this question, we took advantage of an assay that has been developed to identify and quantitate spontaneous mutations that arise within a small plasmid-borne reporter gene (KUNZ
et al. 1989). Isogenic strains CH2213 (CDC44) and CH2214 (cdc44-1) were constructed bearing the chro- mosomal canl-100, ade2-1, lys2-1 and ura3-52 alleles, as well as a nonessential, extrachromosomal, centromere plasmid-bearing URA3 and SUP4-o. In these strains, SUP4-o supresses the canl, ade2, and lys2alleles, resulting in cells that are phenotypically Can', Ade+, and Lys+. Mutations arising within the 89-bp s i P 4 - o gene produce cells that are Can', Ade-, and Lys-, and they can be selected by plating the cultures on plates containing canavanine (KUNZ et al. 1989). We observed that for the CDC44 strain, the mean frequency of canavanine- resistant mutants was 0.8/106 cells (Table 2). This fre- quency is similar to the value (2.0/106 cells) that was previously reported as the mean mutation frequency for the S W 4 - o gene in the parental wild-type strain MKP-o
(KUNZ et al. 1989). In contrast, the cdc44-1 strain exhib- ited a mean mutation frequency of 12/106 cells. This 15-fold increase is consistent with our previous analysis of mutation frequencies in the 1.8-kb CAN1 gene in
CDC44 and cdc44 strains (MCALEAR et al. 1994). To examine the sequence-specificity of the mutator phenotype associated with cdc44 mutations, we se- quenced 30 independent s i P 4 - o mutations that arose from either CDC44 or cdc44-I strains.
As
had been ob- served previously with the wild-type parent (MKP-o)(KUNZ et al. 1989), the majority of the mutations in both the CDC44 and cdc44-1 cultures were single bp substitutions (87 and 83%, respectively) (Table 2).
70 M. A. McAlear, K. M. Tuffo and C. Holm
TABLE 2
Spectrum of nucleotide changes in cd& and CDC44 strains
W C 4 4 cdc44-1 MKP-0"
Frequency of Canr/106 cells 0.8 12 2.0
Total no. of mutants examined 30 30 222
% of mutants with single
basepair substitutions 87 83 82
Breakdown of basepair substitutions Transitions (%\
G C -+ A-?: 29 20
Total 43 37
A-T -+ G C 14 17
Transversions (%)
G C -+ T-A 21 43
G C -+ C-G 29 7
A-T + C-G 4 3
A-T -+ T-A - 4 10
58 63
Total -
25 - 17 42
32 19 2 - 6 59
a As reported by KUNZ et al. (1989).
cdc44-1 strains ( P
<
0.05). We did, however, observe that the cdc44-1 strain exhibited a doubling in the fre- quency of G C to T-A mutations, suggesting that the mutator phenotype might be related, in part, to the metabolism of G C base pairs. In both W C 4 4 and cdc44 strains, the positions of the mutations were distributed throughout the SUP4-o gene and were reminiscent of the positions previously reported for the wild-type MKP- o strain (Figure 2) (KUNZ et al. 1989). Thus, although cdc44-1 mutants exhibit an increased frequency of spon- taneous mutations, these results show that the spectrum of the mutations is not dramatically different than those observed in W C 4 4 strains.cdc44 mutants are delayed in progressing through S
phase: Since Cdc44p is an integral component of the DNA replication apparatus, an increased mutation fre- quency could arise from defects in DNA replication or DNA repair. Although RF-C has been shown to be required for processive DNA replication in vitro, the role it plays in DNA synthesis in vivo has not been deter- mined. To determine whether RF-C is required for DNA replication in vivo, we monitored the progression of DNA synthesis in synchronized CDC44 (strain CH1806)
"
6 hours
8 hours
1 2 hours
FIGURE 3.-"icrofluorometric analysis of CDC44 and cdc44 strains. Strains CH1806 (CDC44) and CH1807 (cdc44) were grown to early log phase at 30", arrested in G1 with alpha factor, and then released from arrest at either 30" or 14". Samples were fixed, stained with propidium iodide and counted by fluorescence activated cell sorter.
and cdc44-1 (strain CH1807) cultures (Figure 3 ) . Mu- tant and wild-type cells were arrested in G1 with alpha factor, released into fresh medium at the permissive or restrictive temperature, and monitored for DNA synthe- sis by microfluorometric analysis. When CDC44 cells are released from alpha factor arrest at 30°, they proceed into S phase, and by 30 min one can detect cells with a G2/M DNA content (Figure 3A). When cdc44 cells are released at 30", they progress through the cell cycle with similar kinetics (Figure 3B). However, when these strains are released from alpha factor arrest into me-
CUI244 A
T
A T T T
C A TCC A GA C
T
A T T A A A G TA AG AAC
3 ' GAGAGCCATC GGTTCAACCA ATTCCGCGT TCTGAAATTA AATAGTZA'E CTTTAGAACT CTAGCCCGCA AGCTGAGCGG GGGCCCTCT 5 '
1 10 20 3 0 4 0 50 60 70 80 89
C G T T C AA C TC C C A T G C T G TC
A T T C A T C
T
CDC4 4
RFC in DNA Replication and Repair 71
dium at 14", the cdc44 mutants progress through S
phase very slowly. Whereas the majority of the CDC44
cells have a G2/M DNA content
2.5
hr after release from alpha factor (Figure 3C), cdc44 mutants only begin to accumulate cells with a G2/M DNA content 8 hr after release from alpha factor (Figure 3D). The increase in DNA content in cdc44 cells is not due to an accumula- tion of mitochondrial DNA, because similar results were obtained with strains CH2211 (CDC44) and CH2212(cdc44), which lack mitochondrial DNA (data not shown). These results demonstrate that cdc44-I mutants experience a delay in traversing S phase at the restric- tive temperature.
To rule out the possibility that cdc44 mutants are able to traverse S phase because of an aberrant function of Cdc44lp at the restrictive temperature, we depleted Cdc44p from cells using transcriptional inactivation in cells grown only at the permissive temperature. We en- gineered strains that could be depleted of Cdc44p by regulating its expression with the repressible GAL1 pro- moter (PGAL). The plasmids pCH1523 (PGALI-CDC44)
or pCH1524 (pGAL1-cdc44-I) were transformed into a CDC44 deletion strain, and the transformants were maintained on galactose (inducing) media. Unfortu- nately, the depletion experiment could not be per- formed with the strain (CH2088) bearing the PGAL-
CDC44 construct, because the residual transcription of the pGAL-CDC44 construct allowed the cells to grow even in the presence of glucose. However, the CDC44
deletion strain carrying the PGAL-cdc44-1 plasmid (CH2089) grew on galactose medium but not on glu- cose medium, indicating that Cdc44p could be de- pleted from the cells by growing them on glucose.
PGAL-cdc44-1 cells were grown to early log phase under inducing conditions (in W raffinose containing galac- tose) and then shifted to medium containing glucose to repress the expression of the cdc44-1 gene. Consistent with the earlier temperature-shift results, PGAL-cdc44-1
cells arrested with a G2/M DNA content before mitosis (data not shown). Thus it appears that although Cdc44p is normally involved in DNA replication, it is nonetheless possible for the cell to complete the bulk of DNA replication even after Cdc44p activity drops below a critical level.
Cdc44p activity is required to traverse the G2/M boundary: Regardless of the method used for reducing Cdc44p activity in the cells, cells with reduced Cdc44p activity always undergo a cell cycle arrest with a G2/M DNA content (Figure 3 ) (HOWELL et al. 1994). This result demonstrates that although normal levels of Cdc44p activity are not essential for the bulk of DNA synthesis, they are required for some process that allows the cells to continue with the cell cycle. To determine whether Cdc44p has an activity that is required at the end of S phase, we examined the time of action of the Cdc44p function that is inactivated by the cdc44-I muta-
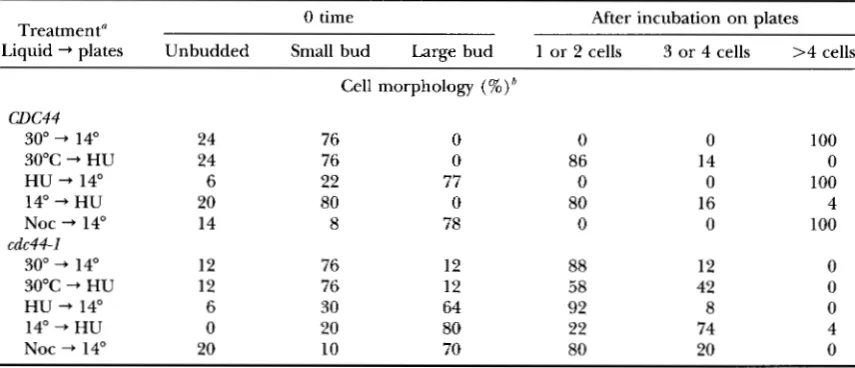
tion. We used reciprocal shift experiments to determine the order of the Cdc44p execution point with respect to other defined points within the cell cycle. In these experiments, cdc44 cells are alternately arrested and re- leased from arrest under various conditions, and the progression of the cells through the cell cycle is deter- mined by monitoring cell morphology
UARVIK
and BOTSTEIN 1973; HEREFORD and HARTWELL 1974).First, we investigated the relationship between the execution points of Cdc44p and the drug hydroxyurea (Table 3). Hydroxyurea is a reversible inhibitor of the enzyme ribonucleotide reductase, and treatment of cells with hydroxyurea leads to a depletion of nucleo- tide pools, causing cells to arrest in S phase (SLATER
1973). When cdc44-I mutants are first arrested in hy- droxyurea at 30" and then washed free of hydroxyurea and plated on W D plates at 14", they fail to progress through the cell cycle, remaining arrested as large bud- ded cells. In contrast, if cdc44-I cells are first arrested at 14" and then placed onto hydroxyurea plates at 30",
the cells progress through the first cell cycle and then arrest in the second cell cycle. Formally, these results demonstrate that the Cdc44prequiring step is depen- dent upon the HU-sensitive step. Thus, it appears that when cdc44 cells are arrested at 14", they are at a point in the cell cycle after the hydroxyurea-sensitive step, which is usually thought of as the bulk of DNA replica- tion (SLATER 1973). This interpretation is consistent with the results of the microfluorometric analysis of synchronized cells.
Second, we investigated the relationship between the execution points of Cdc44p and the drug nocodazole (Table 3). Nocodazole causes reversible depolymeriza- tion of microtubules, and cells treated with nocodazole arrest after the completion of S phase and at the begin- ning of mitosis (JACOBS et al. 1988). Unexpectedly, when cdc44-1 cells are first arrested at 30" in nocodazole and then shifted into nocodazole-free medium at 14", the cells fail to progress through the cell cycle. Because this result is unchanged if the initial growth and noco- dazole arrest are performed at 35", [a temperature at which cdc44-1 mutants exhibit a normal FACS profile
(data not shown)], it is unlikely that the choice of per- missive temperature has any large influence on the ex- periment. This result suggests that nocodazole-arrested cells still have a requirement for Cdc44p activity before they can continue through the cell cycle. However, only the cdc44-1 allele revealed this unusual execution point for Cdc44p activity; cells bearing the cdc44-2, cdc44-4,
72 M. A. McAlear, K. M. Tuffo and C. Holm
TABLE 3
Relationship between cdc44-1, hydroxyurea, and nocodazole execution points
Treatment" 0 time After incubation on plates
Liquid -+ plates Unbudded Small bud Large bud 1 or 2 cells 3 or 4 cells >4 cells
w c 4 4
30" -+ 14"
30°C -+ HU
HU -+ 14"
14" -+ HU
Noc -+ 14"
30" "* 14"
30°C -+ HU
HU -+ 14"
14" -+ HU
Noc -+ 14"
cdc44-1
24
24
6 20 14
12
12
6
0
20
Cell morphology (%)*
76 0 0
76 0 86
22 77 0
80 0 80
8 78 0
76 12
88
76 58 12 30
64 92
20 80 22
10 80 70
0
14
0
16
0
12
42
8 74 20
100 0 100 4 100
0 0 0 4 0
a Strains CH1806 ( W C 4 4 ) and Ch1807 (cdc44-1) were grown exponentially in liquid medium at 30°, treated
for 1.5 generation times as indicated under liquid, sonicated, and then plated onto W D plates or W D plates containing HU. The morphologies of the cells were determined immediately before plating (0 time) and again after 5 hr at 30" or 16 hr at 14". Unless indicated otherwise, the cells were incubated at a temperature
of 30" and in the absence of any drug. Each regimen was repeated at least twice; representative results are shown.
"The number of cell bodies in each microcolony was determined by phasecontrast microscopy of cells on
plates.
cdc44
mutants are defectivein
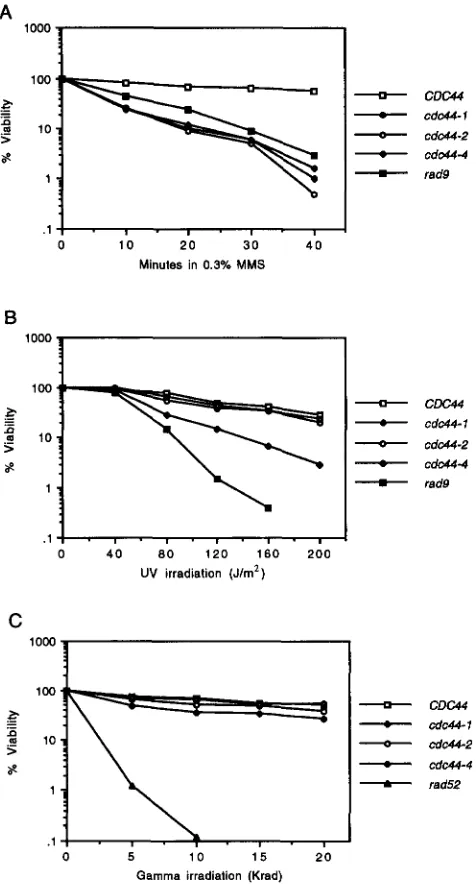
repairing single-stranded DNA breaks: If Cdc44p plays a role in DNA repair in addition to its role in DNA replication, then cdc44 mutants might be sensitive to one or more geno- toxic agents. To test this hypothesis, we assayed whether cold-sensitive cdc44 mutations were associated with an increased sensitivity to the DNA-damaging agent MMS or to exposure to UV or gamma radiation. These genotoxic agents were chosen for their ability to induce a range of DNA damage, including chemically damaged DNA bases (MMS and W), and double-stranded DNA breaks (gamma radiation). Each of the cdc44 mutants tested had a 10- to 100-fold greater sensitivity to MMS exposure than did the CDC44 strain (Figure 4A). Sensitivity to
UV irradiation was milder: the cdc44-1 allele caused the greatest sensitivity, -10 times greater than W C 4 4 ; cdc44-
2 and cdc44-4 exhibited no significant increase in UV
sensitivity (Figure 4B). None of the cdc44 strains were particularly sensitive to gamma irradiation (Figure 4C). Taken together, these results suggest that Cdc44p may play a role in the repair of damaged DNA bases but not in the repair of double-stranded DNA breaks.
During both DNA replication and the repair of dam- aged DNA bases, single-stranded gaps are produced in the DNA, and these gaps must be processed to yield intact double-stranded DNA. cdc44 mutants might therefore be defective in the processing of single- stranded regions of DNA. To test this hypothesis, we assayed for single- and double-stranded breaks in chro- mosomal DNA by using velocity sedimentation analysis
with neutral and alkaline sucrose gradients. We used strains lacking mitochondrial DNA for this analysis to ensure that the effects we observed were due entirely to shifts in the state of chromosomal DNA. Strain CH2211 (CDC44) was labeled with 3H uracil, and strain CH2212 (cdc44-I) was labeled with 14C uracil. To facilitate the comparison between the profiles of W C 4 4 and cdc44 strains, DNA from both strains was sedimented within the same gradient. Chromosomal DNA was isolated from both strains, and the samples were sedimented on 15-30% neutral or alkaline sucrose gradients. Double- stranded breaks in the DNA would be revealed by al- tered sedimentation on both neutral and alkaline su- crose gradients. Single-stranded breaks would be re- vealed by altered sedimentation on alkaline gradients.
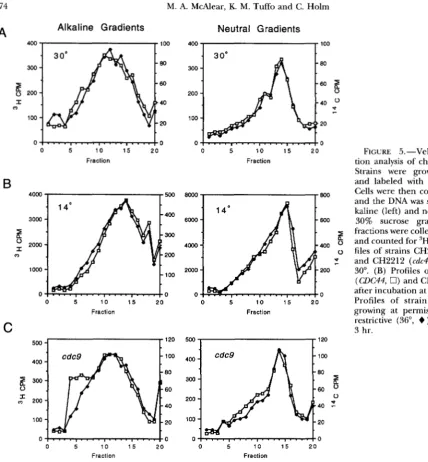
We first compared chromosomal DNA from expo- nentially growing CDC44 and cdc44 strains at permissive or restrictive temperature. When the strains were la- beled at the permissive temperature of 30°, the sedi- mentation profiles of the DNA from the two strains were indistinguishable in both neutral and alkaline gradients (Figure 5A). For the alkaline gradient profiles, the num- ber-average molecular weight was calculated to be 3.1
X 10'. For the neutral gradient profiles, the number- average molecular weight was 6.9 X 10'. This value for the neutral gradients is consistent with the known sizes of yeast chromosomes and is similar to the value (6.2
RFC in DNA Replication and Repair 73
A
loo0
A
-
C W-
cdc44-15
-
cdc44-2ae
-
-
cdc44-4rad9
. 1 : .
,
.
,
.
,
.
, I0 1 0 2 0 3 0 4 0 Minutes in 0.3% MMS
B
loo0
I
1
-
C W-
Cdc44-1-
cdw-2-
cdc44-4-
rad91 ' I
. , , . , . , . , ~ , ~ , ~
0 4 0 80 1 2 0 160 2 0 0
UV irradiation (Jim')
C
lorn
I
C D W
._
-
cdc44-1n
5 a 10
-
cdc44-2-
rad52.. , -
0
;
1 0 1 5 20~ I ~ , ~ ,
Gamma irradiation (Krad)
FIGURE 4.-Sensitivity of strains to DNAdamaging agents.
Strains CH1806 (CDC44, CH1807 (cdc44-I), CH1589 (cdc44-2),
CH1591 (cdc44-4), CH1634 (rad52), and CH2027 ( r a d 3 were
grown to early log phase and exposed to the DNAdamaging agent as indicated. The cells were diluted as necessary, plated onto WD plates, incubated at 30" for 3 days and scored for growth of colonies. (A) Sensitivity to MMS exposure. (B) Sensi-
tivity to UV radiation. (C) Sensitivity to gamma ray radiation.
we cultured D C 4 4 and cdc44 strains at the permissive temperature, shifted them to the restrictive tempera- ture (14") for 10 hr, and then processed the samples (Figure 5B). Once again, we found that the CDC44 and
cdc44 profiles were indistinguishable. Because we de- tected no difference between cdc44 and CDC44 strains, we examined a cdc9 (DNA ligase) mutant as a control for our sedimentation analysis. As expected, the sedi- mentation profile of strain CH2115 (cdc9) grown at the restrictive temperature (36") for 3 hr revealed a shift
in the alkaline gradient profile toward the top of the gradient (equivalent to a decrease in the number-aver- age molecular weight by 30%, which corresponds to a 43% increase in the level of single-stranded DNA
breaks). The neutral sedimentation profile of the cdc9
mutant at the restrictive temperature was similar to the profile at the permissive temperature. Thus, although we could easily visualize single-stranded DNA breaks in a cdc9 mutant, much less breakage must be present in
cdc44 mutants.
Although any difference between the levels of DNA
breaks in unperturbed c1DC44 and cdc44 cells must be low, we reasoned that it might be possible to visualize increased levels of DNA breaks in cdc44 cells following exposure to a genotoxic agent if Cdc44p plays an essen- tial role in DNA repair. To examine this possibility, we determined whether cdc44 mutants had elevated levels of DNA breaks after exposure to UV light. When strains CH2211 (CDC44) and CH2212 (cdc44) were treated with 100 J/m' of UV irradiation, allowed to recover at 23" for 5 min, and then collected for sedimentation analysis, the alkaline DNA sedimentation profiles of both strains were shifted toward the top of the gradient (Figure 6A). In separate gradients in which irradiated and unirradiated samples from a given strain were run together (data not shown), we found that the number average molecular weight of the DNA decreased by only 10% for O C 4 4 cells. In contrast, the number-average molecular weight of the DNA decreased by 30% in the
cdc44 samples. This result may reflect the rapidity with which CDC44 cells can repair W damage, and it sug- gests that cdc44 mutants may be deficient in W repair. To determine whether cdc44 mutants are deficient in W repair at the restrictive temperature, we examined the kinetics of recovery at both permissive and restric- tive temperatures. DNA breaks induced by UV irradia- tion were repaired if the cultures were irradiated and then allowed to recover at permissive temperature (30")
before collecting for sedimentation analysis. For the
74 M. A. McAlear, K. M. Tuffo and C. Holm
A
Alkaline Gradients Neutral Gradients
0 '
0
0 5 1 0 1 5 20
Fraction
B
4000
3000
5
n
;
20000
1000
0
0 5 1 0 1 5 2 0
Fraction
C
400
-
100
300
-
80
0 60
2
200-
40
100
-
20o ! ....,....,....I....
!
~
0 5 1 0 1 5 2 0
0
0 5 1 0 1 5 2 0 Fraction
800
600
I
400 P
0 P
r 200
0
0 5 10 1 5 2 0
Fraction
FIGURE 5 . -Velocity sedimenta- tion analysis of chromosomal DNA.
Strains were grown exponentially
and labeled with 'H or I4C uracil. Cells were then collected and lysed, and the DNA was sedimented on al- kaline (left) and neutral (right) 15- 30% sucrose gradients. Gradient
fractions were collected, reci itated and counted for 'H and C. (A) Pro- files of strains CH2211 (CDC44, 0) and CH2212 (cdc44,
+
) growing at30". (B) Profiles of strains CH2211 (CDC44, 0) and CH2212 (cdc44,
+
)after incubation at 14" for 10 hr. (C)
Profiles of strain CH2215 ( c d c 9 )
growing at permissive (23", 0) or restrictive (36",
+
) temperature for3 hr.
R P
500
,
,
120400
-
300
-
200
-
100-
0 .
. . .
. . .
:
0 5 1 0 1 5 20
100
80
60
0
40 2
20
0
Fraction Fraction
shown); the cdc44 mutant maintained a 20% reduction in number average molecular weight even after 21 hr at 14". This failure of cdc44 mutants to process UV-
induced single-stranded DNA breaks at the restrictive temperature is consistent with the hypothesis that Cdc44p plays a role in the DNA excision repair pathway.
DISCUSSION
We report that the large subunit of RF-C (Cdc44p or Rfclp) is involved in both DNA replication and DNA repair in S. cereuisiae. We have characterized cold-sensi- tive mutant alleles of the CDC44 gene and demonstrate that when cdc44 mutants are released from an alpha factor arrest to the restrictive temperature, the mutant cells are delayed in progressing through S phase. cdc44
mutants also exhibit a general mutator phenotype, and
to the methylating agent MMS. When strains are ex- posed to U V irradiation and then allowed to recover at 14", cdc44 mutant cells retain higher levels of single- stranded DNA breaks than do wild-type cells. These results, coupled with the known biochemical activities of RF-C, are consistent with the hypothesis that RF-C plays a role DNA synthesis and is involved in the metab- olism of single-stranded DNA gaps in both DNA replica- tion and DNA repair in vivo.
RFC in DNA Replication and Repair
A
75
B
80
60
B
0 40 0
P
-
20
0 0 5 1 5 1 0 20
Fraction
C
0 5 1 0 1 5 2 0
Fraction
D
.""..
,CDC44 cdc44
250 FIGURE 6.-Velocity sedimentation
200
150
E
were grown at 30°, irradiated with 100 50 analysis of chromosomal DNA after UV irradiation. Strains CH2211 40 (CDC44, 0) and CH2212 (cdc44,+
) 3o 0 J/mz of UV irradiation, and then al-30 min at 30" (B), 1 hr at 30" (C), 6
Samples were collected and lysed, 10 and the DNA was sedimented on al-
kaline 15-30% sucrose gradients. 100 p lowed to recover for 5 min at 23" (A),
50 2o hr at 14" (D), or 21 hr at 14" (E).
0
0 5 1 0 1 5 20 Fraction
E
1 1 0 0 120
600
-
6 hrs. 100 8080 60
0
60 0
40
20
40
-
20
0 o !
. . . . , . . . . , . . . . , . . . .
!0 5 1 5 1 0 2 0 0
Fraction
0 5 1 5 1 0 20 Fraction
an exponentially growing culture of cdc44 cells is shifted from the permissive temperature to the restrictive tem- perature, the cells also arrest with a G2/M DNA con- tent, and that this cell cycle arrest is mediated by the m y d e p e n d e n t checkpoint (HOWELL et al. 1994). Thus, although the bulk of DNA replication can eventu- ally proceed in cdc44 mutants at the restrictive tempera- ture, it appears that unreplicated or damaged regions of the genome persist and prevent the cell cycle from progressing into mitosis. Cdc44p might therefore play a critical role in a final stage of DNA metabolism that ensures that the genome is in a state suitable for the initiation of mitosis.
The sensitivity of cdc44 mutants to both MMS expo- sure and UV irradiation suggests that RF-C is involved in the DNA excision repair process. The alkylating agent MMS produces 7-methylguanine (7MeG) and 3-
76 M. A. McAlear, K. M. Tuffo and C. Holm
propose that after the excision of the damaged DNA
strand, RF-C binds to the single-stranded gap and through its association with PCNA stimulates DNA poly- merase activity. That RF-C functions in DNA repair through its interaction with PCNA is supported by the observation that PCNA (poZ30) mutations can suppress the DNA repair defects of cdc44 mutations (M. A. Mc-
ALEAR and C. HOLM, unpublished data). What is not clear, however, is why the processivity factor PCNA is used in the polymerization of very short stretches of
DNA. Perhaps in this context, the loading of PCNA
onto the double helix is required for recruiting DNA
polymerase to the site of DNA repair, rather than for increasing its processivity (SHWJI et al. 1992; LI et aZ.
1994; ~ O U S S E K H R A et al. 1995). That cdc44 mutants are not particularly sensitive to gamma ray radiation suggests that RF-C is not involved in the repair of dou- ble-stranded DNA breaks.
While we have demonstrated that Cdc44p plays an important role in the repair of UV damage, the essential role of the protein in unirradiated cells is less clear. Velocity sedimentation analysis indicates that cdc44 mu- tants are compromised in their ability to process single- stranded DNA breaks at the restrictive temperature. When W C 4 4 and cdc44 cells are irradiated with UV
light and then allowed to recover at 14"C, the cdc44
mutants exhibit a higher level of single-stranded DNA
breaks than do W C 4 4 cells. Thus, Cdc44p probably plays a role in the DNA excision repair process after the damaged bases are excised from the DNA duplex. It is puzzling, however, that unirradiated cdc44 mutants arrest before mitosis at 14". Since DNA replication pro- ceeds slowly at 14" in cdc44 mutants, it would not be surprising if the integrity of the DNA were somehow compromised. Nevertheless, we were unable to detect a significant difference in the levels of DNA breaks in unirradiated CDC44 and cdc44 strains at 14". One possi- bility is that low levels of DNA breakage may be present in cdc44 mutants, and that this damage precludes cells from entering into mitosis. It has recently been re- ported that spontaneous DNA damage can occur in highly transcribed regions of DNA (DATTA and JINKS- ROBINSON 1995). Furthermore, it has been demon- strated that as little as one double-stranded DNA break per genome is sufficient to stop the cell cycle in S.
cmeuisiae (BENNETT et al. 1993). Perhaps at the restrictive temperature of 14", the activity of the mutant Cdc44
l p
is insufficient to deal with the final DNA gap-filling requirements created by the processes of DNA replica- tion and DNA repair.Since RF-C is involved in both DNA replication and
DNA repair, what is the source of the mutator pheno- type associated with cdc44? Our results suggest that de- fects in both DNA repair and DNA replication could contribute to this phenotype. The spectrum of sponta- neous mutations observed in cdc44 mutants suggests
that the mutations do not arise from a gross defect in
DNA polymerization (i.e., DNA polymerase slippage,
DNA insertions or deletions). Rather, the majority of the spontaneous mutations produced in cdc44 mutants are single base pair substitutions. Although cdc44 mu- tants have a 15-fold increase in the frequency of sponta- neous mutations over CDC44 cells, the spectra of muta- tions found in the two strains are generally similar to one another. One potentially informative exception, however, is the increase in G C to T-A transversions in
cdc44 mutants. When alkylated bases are removed from the DNA duplex and the subsequent abasic sites escape
DNA repair, there is a preferential incorporation of adenine opposite the abasic site during DNA replica- tion (LOEB and PRESTON 1986). Thus, G C to T-A trans- versions could arise from the incomplete repair of dam- aged G residues (ROMOTAR et al. 1991). Spontaneous alkylation is a known source of 7MeG in
S.
cereuisiae,and mutations in the enzymes that process 7MeG in- crease the frequency of spontaneous mutation by three- to fivefold (XIAO and SAMSON 1993). These results, cou- pled with the observation that the major form of dam- age induced by MMS is 7MeG, support the hypothesis that the cdc44-associated mutator phenotype may be due in part to a defect in the base excision repair path- way. However, since mutants completely defective in 7MeG processing only have a three-to fivefold increase in mutation frequency (XIAO and SAMSON 1993), there must be an additional cause for the elevated mutation rate in cdc44 strains.
Another source for the increased rate of mutation seen in cdc44 mutants might be errors produced during
DNA synthesis due to disruptions in the usage of the
DNA polymerases. In vitro biochemical analysis indi- cates that RF-C and PCNA stimulate the activities of
DNA polymerases
S
and E (LEE and HURWITZ 1990; BURGERS 1991; LEE et al. 1991; FIEN and STILLMAN 1992;PODUST et al. 1992). Of the five DNA polymerases in S.
cereuisiae, these are the only two that have an intrinsic 3'-5' exonuclease proofreading activity (MORRISON and
SUCINO 1994). Therefore, if the DNA polymerase stimu- lating activities of RF-C and PCNA were compromised in cdc44 mutants, those processes that would normally be carried out by DNA polymerase