Molecular analysis of the determination of developmental fate in the small intestinal epithelium in the chicken embryo
Full text
Figure
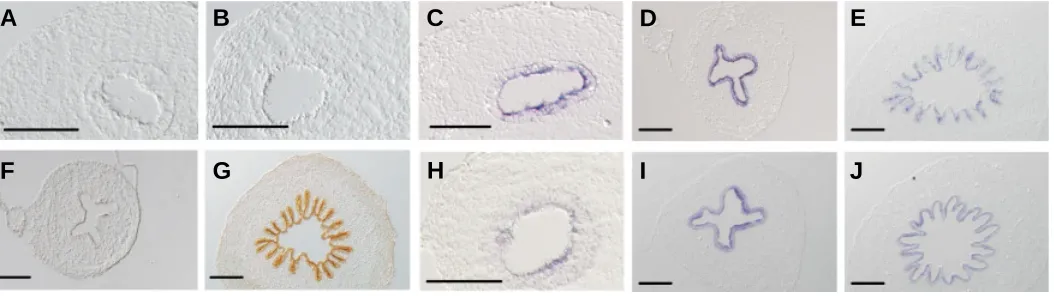
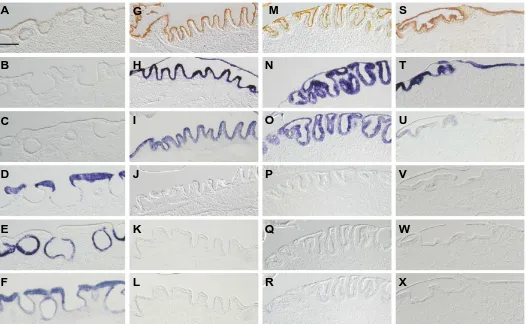
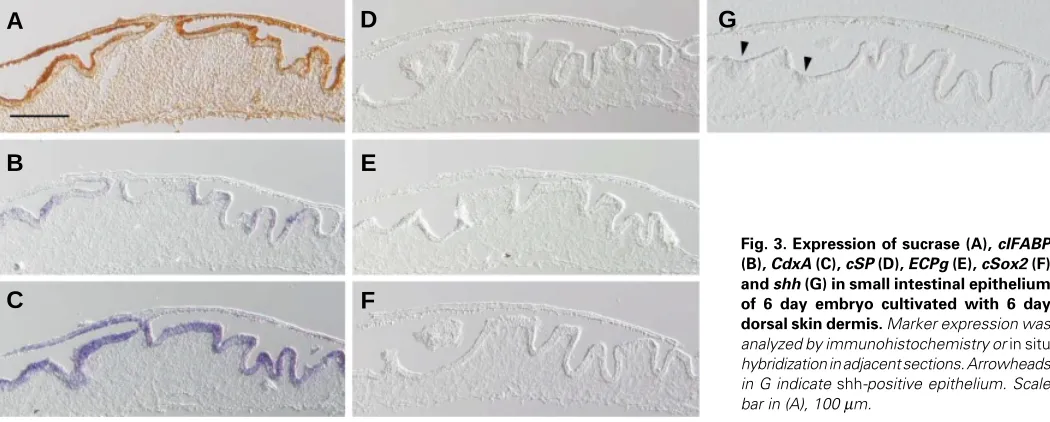
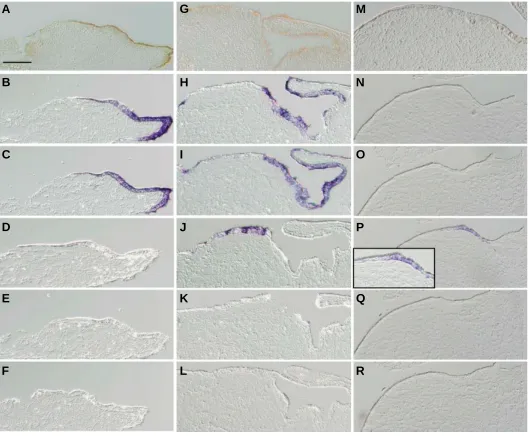

Related documents
Dynamics of fungal invasion, epithelial damage, and loss of epithelial barrier integrity during translocation of Candida albicans through intestinal epithelial layers.. A combination
AIRWAYS ICPs: integrated care pathways for airway diseases; ARIA: Allergic Rhinitis and its Impact on Asthma; COPD: chronic obstructive pulmonary disease; DG: Directorate General;
It was decided that with the presence of such significant red flag signs that she should undergo advanced imaging, in this case an MRI, that revealed an underlying malignancy, which
Also, both diabetic groups there were a positive immunoreactivity of the photoreceptor inner segment, and this was also seen among control ani- mals treated with a
The main purpose of the study is to evaluate the impact of knowledge sharing towards teaching innovation; through technological pedagogical content knowledge and job
ings were observed in the renal tissue of the Cis group with significantly increased levels of oxidants and IL-1β, such as wide hemorrhage areas, dilated and congested blood
19% serve a county. Fourteen per cent of the centers provide service for adjoining states in addition to the states in which they are located; usually these adjoining states have
Field experiments were conducted at Ebonyi State University Research Farm during 2009 and 2010 farming seasons to evaluate the effect of intercropping maize with
