Key words: frog, feeding, behavior, kinematics, Rana pipiens.
THE MODULATION OF FEEDING BEHAVIOR IN RESPONSE
TO PREY TYPE IN THE FROG RANA PIPIENS
CURTIS W. ANDERSON
Physiology and Functional Morphology Group, Department of Biological Sciences, Northern Arizona University, Flagstaff, AZ 86011, USA
Accepted 24 February 1993
Summary
Using high-speed video motion analysis, the kinematics of feeding behavior was studied in Rana pipiens. Rana pipiens exhibits differing behavior patterns depending upon prey type. When feeding on small prey such as waxworms, R. pipiens uses tongue protraction to catch prey, minimizing head and body movements. When feeding on the larger earthworm, it arches its body, flexes the head downwards and uses jaw prehension to capture the prey. Time to completion of tongue retraction, time to completion of mouth closing, duration of tongue retraction, duration of mouth closing, total time that the mouth is open and maximum tongue reach were significantly longer during waxworm feedings than during earthworm feedings. The tongue angle and degree of head flexion were significantly greater during earthworm feedings. These different kinematic patterns correspond to differences in feeding behavior among major taxa of anurans. Phylogenetically primitive archaeobatrachian frogs have short tongues which can be protracted only a few millimeters. In contrast, many neobatrachians have long tongues which they project to catch small prey. The similarity between the behavior that neobatrachians use to catch large prey and the movement patterns of archaeobatrachians suggests that Rana pipiens has retained the plesiomorphic archaeobatrachian motor pattern and uses it to catch large prey, while evolving a new motor pattern for catching small prey.
Introduction
The feeding behavior of teleost fishes and amniotes has been shown to vary in response to different prey stimuli. Differences in prey size, shape and mobility may produce differences in the kinematics and motor patterns of feeding. Among centrarchid fishes, feeding behavior has been shown to differ quantitatively when feeding on differing prey items (Lauder, 1981; Wainwright, 1986; Wainwright and Lauder, 1986) and when the locomotor behavior of the prey differs (Elshoud-Oldenhave and Osse, 1976; Liem, 1978, 1980).
Mammalian mastication exhibits diversity both within and between species. Feeding patterns depend not only on prey type and size, but also on the consistency of the prey item. Mammalian mastication is used not only for prey capture, but for prey reduction as well (deVree and Gans, 1974; Gans et al. 1978; Hiiemae and Crompton, 1985).
Among amphibians, no previous studies have, to my knowledge, documented the modulation of feeding behavior in response to different prey items. Urodeles (salamanders) have not been observed to modulate their pattern of feeding behavior (Findeis and Bemis, 1990; Thexton et al. 1977; Lauder and Shaffer, 1985; Reilly and Lauder, 1989; Roth et al. 1990). In urodeles, all of the muscles involved in feeding are activated simultaneously (Thexton et al. 1977; Lauder and Shaffer, 1985). Because muscles such as the main tongue retractors (the m. rectus cervicus/abdominis complex) are activated nearly simultaneously with the tongue protractors (the m. subarcualis rectus), there is little possibility for behavioral modulation once the feeding bout has been initiated (Thexton et al. 1977; Roth et al. 1990). Thus, the pattern of prey capture is more stereotyped than in amniotes or fishes because of morphological and neuroanatomical constraints.
The feeding behavior of anurans also has been considered to be highly stereotyped (Ewert, 1987; Roth, 1987; Roth et al. 1990). The stereotypy of feeding behavior found in anurans has been attributed to a limited repertoire of fixed action patterns and to the animal’s behavioral simplicity (Ewert, 1987). Detailed kinematic or electromyographic (EMG) analyses have been carried out only for a few species (Gans and Gorniak, 1982a,b; Matsushima et al. 1985; Deban and Nishikawa, 1992; Nishikawa and Roth, 1991; Nishikawa and Cannatella, 1991). Most of the EMG studies have been on the genus
Bufo, which prefer small prey (Zug and Zug, 1979; Gans and Gorniak, 1982a; Duellman
and Trueb, 1986).
The goal of this work is to study the feeding behavior of a generalized predator, Rana
pipiens, and to quantify the movement patterns involved during feeding on different prey
types. Rana pipiens, like bufonids, is a neobatrachian, or derived anuran, which possesses a highly protrusible tongue. By studying a neobatrachian that is a dietary generalist, I hope to understand further the behavioral and neuromuscular changes that have accompanied the evolution of a highly protrusible tongue. In addition, if Rana pipiens does modulate its feeding behavior in response to differences in prey type, this study will be the first to document this ability among amphibians. The kinematic studies presented here are part of a long-term study on the evolution and neural control of anuran feeding behavior.
Materials and methods
Adult Rana pipiens were obtained from animal suppliers. The individuals used in this study ranged from 62 to 67mm in snout–vent length (SVL). To study the kinematics of feeding behavior, earthworms (Lumbricus sp.; 7–12cm) and waxworms (Galleria sp.; 1.5–2.0cm) were used as prey. The prey were chosen to provide variation in size, mass and mobility.
synchronized stroboscopic illumination, and were allowed to feed unrestrained on a flat stage covered with a damp paper towel. Filming was carried out at room temperature (approximately 21–23˚C).
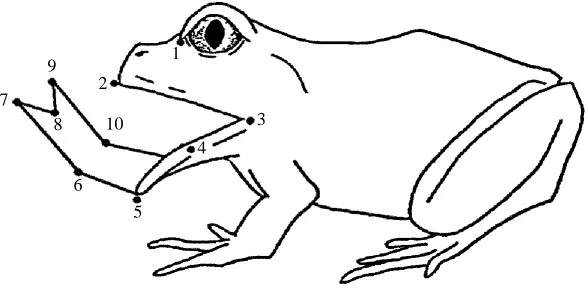
Five feeding sequences per prey type per individual were obtained from five individuals, for a total of 25 waxworm feeding sequences and 25 earthworm sequences. The feeding sequences were analyzed using Peak Performance Technologies 2D motion analysis software on an IBM-compatible computer. For each field of the feeding sequence, the x,y-coordinates of the prey item, a non-moving reference point and 10 points on the frog were digitized from the video monitor (Fig. 1). The video resolution varied between 31.8 and 50.8pixelscm21. A typical feeding sequence consisted of 30–38 fields (250–315ms). All successful feeding attempts were digitized for analysis except those in which the frog or the prey item moved out of view of the camera. Sixty-two sequences were filmed, 50 of which were included in the analysis.
For the kinematic analyses, a series of temporal variables was used to quantify the feeding behaviors. The frame in which the frog began to open its mouth, described as the
onset of mouth opening, was designated as time zero in the analysis. The following
variables were measure in milliseconds (modified from Nishikawa and Cannatella, 1991; Nishikawa and Roth, 1991). (1) Onset of forward head movement: the time at which the first movement towards the prey was observed. (2) Onset of tongue
p r o t r a c t i o n: the time at which the tongue pad first became visible. (3) Time of minimum mandible angle: the time at which the angle subtended by the lower jaw tip, the midpoint
of the lower jaw and the jaw joint was minimal. (4) Time of prey contact: the time at which the tongue made contact with the prey. (5) Completion of tongue protraction: the time at which the tongue was maximally protracted. (6) Onset of tongue retraction: the time at which the tongue first began to be retracted. (7) Time of maximum gape: the time at which the mouth was maximally opened. (8) Completion of tongue retraction: the time at which the tongue became fully retracted. (9) Time to maximum displacement of
1 2
10
5 6 7
8 9
[image:3.612.88.381.445.592.2]3 4
upper jaw tip : the time at which maximum forward movement of the upper jaw tip was
reached during the lunge. (10) Onset of mouth closing: the time at which the mouth began to close. (11) Completion of mouth closing: the time at which the mouth closed. (12) Duration of mouth opening (time zero to variable 7): the time from the onset of mouth opening to the time of maximum gape. (13) Duration of tongue protraction (variable 2 to variable 5): the time from the onset of tongue protraction to the completion of tongue protraction. (14) Duration of tongue retraction (variable 6 to variable 8): the time from the onset of tongue retraction to the completion of tongue retraction. (15) Duration of tongue at target (variable 4 to variable 6): the time from prey contact to the onset of tongue retraction. (16) Duration of mouth closing (variable 7 to variable 11): the time from maximum gape until the mouth was closed. (17) Total time mouth is open (time zero to variable 11): the time from the onset of mouth opening to the completion of mouth closing. Of these timing variables, only the durations (12–17) were used in the statistical analysis.
In addition, the following variables were calculated from the digitized points: (1) maximum gape angle, the maximum angle subtended by the upper and lower jaw tips with the jaw joint at the vertex (Fig. 1, points 2, 3, 5); (2) minimum mandible angle, the angle subtended by the jaw joint, the midpoint of the lower jaw and the lower jaw tip (Fig. 1, points 3, 4, 5); (3) maximum head flexion, the degree to which the head is ventroflexed downward (Fig. 1, points 2 and 3 plotted against the horizon line); (4) maximum tongue reach, the distance from the tip of the anteriormost point on the protracted tongue to the line connecting the upper and lower jaw tips; (5) tongue angle, the angle subtended by the tip of the tongue, the lower jaw tip and the jaw joint (Fig. 1, points 7, 5, 3); (6) absolute gape (cm), the distance between the upper and lower jaw tips at maximum gape; and (7) lunge length (cm), the distance between the tip of the snout at rest and at maximum forward excursion.
Data were analyzed with Statview 512+ statistical software on a Macintosh IIci computer. Paired t-tests were used to test the significance of differences between individual means for the two prey types (a=0.05, two-tailed). To analyze individual differences among frogs, a separate one-way analysis of variance was run for each prey type. A two-way analysis of variance with prey type and individual as main effects was not utilized because the F-test (with individual 3 treatment mean square in the denominator) lacks statistical power and there is a large probability of making a type II error. Values are presented as the mean ± S.E.M.
Results
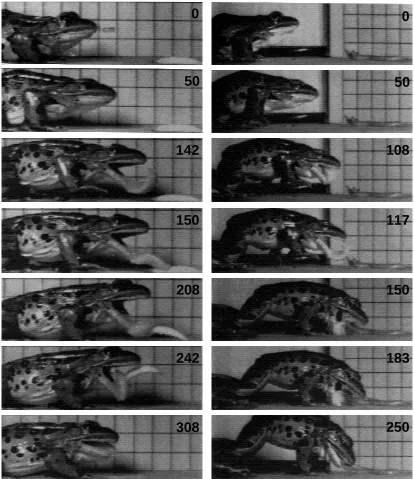
Prey capture in Rana pipiens is composed of four phases: (1) approach/mouth opening; (2) tongue protraction; (3) prey contact/tongue retraction; and (4) mouth closing/body recovery (modified from Nishikawa and Cannatella, 1991; Nishikawa and Roth, 1991; Fig. 2).
Rana pipiens exhibits different patterns of feeding behavior depending upon the type of
are positioned for capturing the prey. When feeding on small prey, the tongue is sufficient for capturing the prey. The movement of the body is minimized, and the jaws do not function for retrieving the prey (Fig. 2).
0
50
142
150
208
242
308
250
[image:5.612.34.448.128.607.2]183
150
117
108
50
0
Approach/mouth opening
The feeding bout begins at the onset of forward head movement. For both prey types, the onset of forward movement occurs 55–57ms before mouth opening. As the mouth opens, the mandibles are bent ventrally and the tongue is projected from the m o u t h .
Although many of the characteristics of the lunge differed between waxworm and earthworm feedings, lunge length did not differ significantly between prey types. However, there were significant inter-individual differences (Tables 1, 2). Waxworm feedings especially illustrated significant inter-individual differences. Many of these differences can be attributed to the location of the prey before the lunge is initiated. Some frogs have a tendency to move closer to the prey before beginning tongue protraction, whereas other individuals exhibit a longer lunge prior to prey capture. There is less room for inter-individual differences when feeding on earthworms. The prey capture must be made with the jaws, so lunge length and variables such as tongue length become less important.
40
0 80
60
20 A
83 166 249
0
Time (ms)
B 150
100
50
0 200
83 166 249
0
Time (ms) C
60 40 20 0 −20 80
83 166 249
0
[image:6.612.61.432.300.599.2]Time (ms)
Tongue protraction
[image:7.612.55.427.201.408.2]Tongue protraction occurs for both prey types, although the timing and degree of protraction differ significantly between prey items. When feeding on waxworms, the body remains stable throughout the feeding bout, with the exception of the forward excursion of the body due to the lunge. The head is moved forward as a result of the rotation of the body over the front limbs, but there is little head flexion (Figs 2 and 3,
Table 1. Mean, standard error, t-value and P-value from a paired-sample t-test
comparing waxworm feedings and earthworm feedings
Earthworm Waxworm
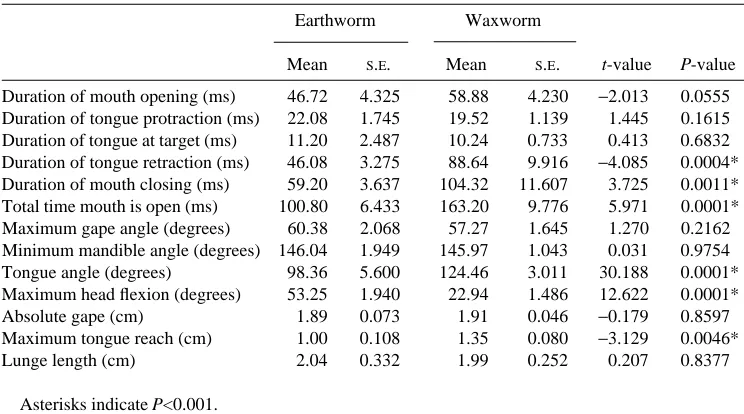
Mean S.E. Mean S.E. t-value P-value Duration of mouth opening (ms) 46.72 4.325 58.88 4.230 −2.013 0.0555 Duration of tongue protraction (ms) 22.08 1.745 19.52 1.139 1.445 0.1615 Duration of tongue at target (ms) 11.20 2.487 10.24 0.733 0.413 0.6832 Duration of tongue retraction (ms) 46.08 3.275 88.64 9.916 −4.085 0.0004* Duration of mouth closing (ms) 59.20 3.637 104.32 11.607 3.725 0.0011* Total time mouth is open (ms) 100.80 6.433 163.20 9.776 5.971 0.0001* Maximum gape angle (degrees) 60.38 2.068 57.27 1.645 1.270 0.2162 Minimum mandible angle (degrees) 146.04 1.949 145.97 1.043 0.031 0.9754 Tongue angle (degrees) 98.36 5.600 124.46 3.011 30.188 0.0001* Maximum head flexion (degrees) 53.25 1.940 22.94 1.486 12.622 0.0001* Absolute gape (cm) 1.89 0.073 1.91 0.046 −0.179 0.8597 Maximum tongue reach (cm) 1.00 0.108 1.35 0.080 −3.129 0.0046*
Lunge length (cm) 2.04 0.332 1.99 0.252 0.207 0.8377
Asterisks indicate P<0.001.
Table 2. Results of a one-way analysis of variance using the individual frog as the main
effect
Earthworm feedings Waxworm feedings
F-value P-value F-value P-value
Duration of mouth opening (ms) 3.698 0.0207 6.983 0.0011 Duration of tongue protraction (ms) 2.286 0.0959 5.857 0.0027 Duration of tongue at target (ms) 1.416 0.2650 10.750 0.0001 Duration of tongue retraction (ms) 2.308 0.0934 9.926 0.0001 Duration of mouth closing (ms) 0.762 0.5622 4.522 0.0092 Total time mouth is open (ms) 1.671 0.1960 2.876 0.0494 Maximum gape angle (degrees) 2.371 0.0870 19.957 0.0001 Minimum mandible angle (degrees) 3.679 0.0211 1.073 0.3961
Tongue angle (degrees) 4.371 0.0106 6.594 0.0015
Maximum head flexion (degrees) 1.374 0.2783 3.437 0.0271
Absolute gape (cm) 3.154 0.0366 6.969 0.0011
Maximum tongue reach (cm) 6.245 0.0020 9.734 0.0002
[image:7.612.58.430.458.646.2]22.9±1.5˚, S.E.M.). The tongue is protracted through a mean tongue angle of 124.5±3.0 ˚ (S.E.M.).
When feeding on earthworms, the body is arched forward, and the head moves forward slightly, as with waxworms, but is ventroflexed downward to a significantly larger degree (Figs 2 and 3, 53.3±1.9˚, t=12.622, P<0.0001). The tongue is protracted, but significantly less than when feeding on waxworms (Table 1, t=23.129, P=0.0046). Because of the arching of the body and larger degree of ventroflexion of the head, the gape angle is directed downward and the tongue angle is significantly smaller (t=30.188, P<0.0001).
Prey contact/tongue retraction
When feeding on waxworms, the tongue contacts the prey, which adheres to the sticky tongue pad, and the prey is returned to the mouth. The duration of tongue retraction (t=24.085, P=0.0004) was significantly longer during waxworm feedings than during earthworm feedings (Table 1).
Mouth closing/body recovery
The duration of mouth closing (t=23.725, P=0.001) was significantly longer during waxworm feedings than during earthworm feedings.
Discussion
The purpose of this study was to compare the feeding behavior of Rana pipiens in response to differing species of prey. When feeding on waxworms, Rana pipiens uses tongue protraction to capture the prey and return it to the mouth. Larger prey, such as earthworms, are not as easily captured by tongue prehension, and R. pipiens arches the body, ventroflexes the head downwards and uses jaw prehension to capture the prey. In both cases, the feeding behavior is a combination of body movement, mouth opening and closing, and tongue protraction and retraction.
Two lines of evidence suggest that R. pipiens uses visual, rather than olfactory or other stimuli, to modulate feeding behavior. First, regardless of prey type, R. pipiens does not strike prey unless it is moving. Second, when a small piece of earthworm was offered, the frogs captured the prey using the feeding behavior characteristic of that for waxworms rather than that for whole earthworms. These observations suggest that visual, rather than olfactory, stimuli are used to modulate feeding behavior in response to different prey. Further experiments are needed to test this hypothesis.
Previous studies of feeding kinematics in anurans (Bufo marinus, Gans and Gorniak, 1982a; Hyla cinerea, Deban and Nishikawa, 1990, 1992) have not addressed the question of whether these anurans modulate their patterns of feeding behavior in response to prey type. This study is the first, to my knowledge, to identify behavioral modulation during feeding in anurans. Among those anurans that do not seem to modulate their behavior, one possible explanation is dietary specialization. Bufonids tend to be small-prey specialists (Toft, 1980; Emerson, 1985) and may have reduced their ability to feed on larger prey, which do not adhere well to the tongue.
The plesiomorphic condition of tongue morphology found in archaeobatrachian anurans is the possession of a muscularized tongue of limited protrusibility (Regal and Gans, 1976; Noble, 1931; Nishikawa and Cannatella, 1991). Because of limited tongue protrusibility, archaeobatrachians such as Discoglossus pictus and Ascaphus truei lunge forward with the body, and the jaws close on the prey to aid in prey capture (Nishikawa and Cannatella, 1991; Nishikawa and Roth, 1991).
The evolution of a derived tongue morphology, such as the highly protrusible tongue found in Bufo marinus, allows for a different behavioral pattern. The head is moved forward slightly by rotation of the body over the forelimbs, the mouth opens but the head remains relatively stationary, the highly protrusible tongue is flipped out of the mouth and contacts the prey item. The prey is then returned to the mouth without touching the jaws (Gans and Gorniak, 1982a; Nishikawa and Gans, 1990, 1992).
The patterns of feeding behavior found in Rana pipiens suggest that the plesiomorphic condition was not lost during the evolution of a highly protrusible tongue. The different behavior patterns that R. pipiens uses for different prey types correspond to differences in feeding behavior between derived and more primitive frogs. The behavioral pattern that
R. pipiens uses to catch large prey is similar to the feeding behavior of
archaeobatrachians. This suggests that Rana pipiens has retained the primitive archaeobatrachian motor pattern and uses it when feeding on large prey. A new motor pattern has apparently evolved for catching small prey.
In contrast to many studies of amphibian feeding behavior, this study illustrates that the feeding behavior of frogs is less stereotypical than once thought, and that frogs may modify their feeding behavior in response to different prey stimuli. This suggests that, during the evolution of a highly protrusible tongue in Rana pipiens, the plesiomorphic neural circuitry was conserved. Additional studies, using an electromyographic analysis of feeding behavior in Rana pipiens synchronized with high-speed video studies, will allow measurement of the motor patterns generated during feeding on different prey types.
References
ANDERSON, C. W. (1990). The effect of prey size on feeding kinematics in two species of ranid frogs. Am. Zool. 30, 140A.
ANDERSON, C. W. (1991). The modulation of feeding behavior in response to visual stimuli in Rana pipiens. Arizona-Nevada Acad. Sci. 26, 14.
DEBAN, S. ANDNISHIKAWA, K. C. (1990). The mechanism of tongue protrusion in Hyla cinerea and its evolutionary implications. Am. Zool. 30, 141A.
DEBAN, S. ANDNISHIKAWA, K. C. (1992). The kinematics of prey capture and the mechanism of tongue
protraction in the green tree frog, Hyla cinerea. J. exp. Biol. 170, 235–256.
DEVREE, F. ANDGANS, C.(1974). Mastication in pygmy goats, Capra hircus. Ann. soc. r. zool. Belg. 105, 255–306.
DUELLMAN, W. E. ANDTRUEB, L.(1986). Biology of Amphibians. New York: McGraw-Hill.
ELSHOUD-OLDENHAVE, M. J. W. ANDOSSE, J. W. M. (1976). Functional morphology of the feeding
system in the ruff – Gymnocephalus cernua (L. 1758) – (Teleostei, Percidae). J. Morph. 150, 399–422.
EMERSON, S. B.(1985). Skull shape in frogs – correlations with diet. Herpetologica 41, 177–188. EWERT, J. P. (1987). Neuroethology of releasing mechanisms: Prey-catching in toads. Behav. Brain Sci.
10, 337–405.
FINDEIS, E. K. ANDBEMIS, W. E. (1990). Functional morphology of tongue projection in Taricha torosa (Urodela: Salamandridae). Zool. J. Linn. Soc. 99, 129–157.
GANS, C., DEVREE, F. ANDGORNIAK, G. C. (1978). Analysis of mammalian masticatory mechanisms: progress and problems. Zentbl. Vet. Med. C. Anat. Histol. Embryol. 7, 226–244.
GANS, C. ANDGORNIAK, G. C. (1982a). Functional morphology of lingual protrusion in marine toads
(Bufo marinus). Am. J. Anat. 163, 195–222.
GANS, C. ANDGORNIAK, G. C. (1982b). How does the toad flip its tongue? Test of two hypotheses. Science 216, 1335–1337.
GORNIAK, G. C., ROSENBERG, H. I. AND GANS, C. (1982). Mastication in the tuatara, Sphenodon punctatus (Reptilia: Rhynchocephalia): structure and activity of the motor system. J. Morph . 171, 321–353.
HIIEMAE, K. M. AND CROMPTON, A. W. (1985). Mastication, food transport and swallowing. In
Functional Vertebrate Morphology (ed. M. Hildebrand, D. M. Bramble, K. F. Liem and D. B. Wake), pp. 262–296. Cambridge, MA: Harvard University Press.
LAUDER, G. V. (1981). Intraspecific functional repertoires in the feeding mechanism of the characoid fishes Lebiasina, Hoplias and Chalceus. Copeia 1981, 154–168.
LAUDER, G. V. ANDSHAFFER, H. B.(1985). Functional morphology of the feeding mechanism in aquatic ambystomatid salamanders. J. Morph. 185, 297–326.
LIEM, K. F. (1978). Modulatory multiplicity in the functional repertoire of the feeding mechanism in cichlid fishes. I. Piscivores. J. Morph. 158, 323–360.
LIEM, K. F.(1980). Adaptive significance of intra- and interspecific differences in the feeding repertoires
of cichlid fishes. Am. Zool. 20, 295–314.
MATSUSHIMA, T., SATOU, M. AND UEDA, K. (1985). An electromyographic analysis of electrically-evoked prey-catching behavior by means of stimuli applied to the optic tectum in the Japanese toad. Neurosci. Res. 3, 154–161.
NISHIKAWA, K. C. AND CANNATELLA, D. C. (1991). Kinematics of prey capture in the tailed frog,
Ascaphus truei. Zool. J. Linn. Soc. 103, 289–307.
NISHIKAWA, K. C. ANDGANS, C.(1990). Neuromuscular control of prey capture in the marine toad, Bufo marinus. Am. Zool. 30, 141A.
NISHIKAWA, K. C. ANDGANS, C.(1992). The role of hypoglossal sensory feedback during feeding in the marine toad, Bufo marinus. J. exp. Zool. 264, 245–252.
NISHIKAWA, K. C. ANDROTH, G.(1991). The mechanism of tongue protraction during prey capture in the frog Discoglossus pictus. J. exp. Biol. 159, 217–234.
NOBLE, G. K. (1931). The Biology of the Amphibia. New York: McGraw-Hill.
REGAL, P. J. ANDGANS, C. (1976). Functional aspects of the evolution of frog tongues. Evolution 30, 718–734.
REILLY, S. M. AND LAUDER, G. V. (1989). Kinetics of tongue projection in Ambystoma tigrinum:
ROTH, G. (1987). Visual Behavior of Salamanders. Berlin: Springer-Verlag.
ROTH, G., NISHIKAWA, K. C., WAKE, D. B., DICKE, U. ANDMATSUSHIMA, T. (1990). Mechanics and
neuromorphology of feeding in amphibians. Neth. J. Zool. 40, 115–135.
THEXTON, A. J., WAKE, D. B. ANDWAKE, M. H. (1977). Tongue function in the salamander Bolitiglossa
occidentalis. Archs oral Biol. 22, 361–366.
TOFT, C. A. (1980). Feeding ecology of thirteen synoptic species of anurans in a seasonal tropical
environment. Oecologia 45, 131–141.
WAINWRIGHT, P. C. (1986). Motor correlates of learning behaviour: feeding on novel prey by
pumpkinseed sunfish (Lepomis gibbosus). J. exp. Biol. 126, 237–247.
WAINWRIGHT, P. C. ANDLAUDER, G. V.(1986). Feeding biology of sunfishes: patterns of variation in the
feeding mechanism. Zool. J. Linn. Soc. 126, 237–247.
ZUG, G. R. ANDZUG, P. B. (1979). The marine toad, Bufo marinus: a natural history resume of native