The synthesis of trehalose by insect fat body is comparable, in certain respects, to the formation of glucose by mammalian liver. In the liver, glucose synthesis is controlled by the pancreatic hormone glucagon, which shifts liver metabolism from glucose utilization (glycolysis, glycogen synthesis) to glucose production. As a result, glycolytic flux is decreased while glycogen degradation and/or gluconeogenesis are stimulated. The concentration of fructose 2,6-bisphosphate, first discovered in rat liver cells by Van Schaftingen et al. (1980) and subsequently described in various vertebrates and invertebrates (for a review, see Pilkis, 1990), is crucial to the control of glycolysis in liver. Fructose 2,6-bisphosphate is not an intermediate of glycolysis or of any other metabolic pathway, but is a metabolic signal that is formed and degraded by a specific enzyme system (Van Schaftingen and Hers, 1981; Van Schaftingen et al. 1982a). It is a potent allosteric activator of the key glycolytic enzyme phosphofructokinase (PFK) and is an inhibitor of the gluconeogenic enzyme fructose-1,6-bisphosphatase (Van Schaftingen et al. 1980; Van Schaftingen and Hers, 1981). Glucagon decreases the content of fructose 2,6-bisphosphate in the liver, thus causing inhibition (deactivation) of phosphofructokinase and activation
(de-inhibition) of fructose-1,6-bisphosphatase. This is, therefore, a crucial part of the mechanism by which liver metabolism is shifted from glucose consumption to glucose production.
Trehalose synthesis, at least in certain insects such as the American cockroach Periplaneta americana, is stimulated by extracts of the corpus cardiacum, a neurohaemal organ attached to the brain (Steele, 1961). The active factors in corpus cardiacum extract, termed hypertrehalosaemic hormones (HrTHs), have been isolated and shown to be blocked peptides (Scarborough et al. 1984; Gäde and Rinehart, 1986; Hayes et al. 1986). The corpus cardiacum extracts stimulate glycogen phosphorylase (Steele, 1963), thus increasing levels of the hexose phosphates glucose 1-phosphate, glucose 6-phosphate and fructose 6-phosphate (Steele et al. 1984), which are equilibrated in the fat body by phosphoglucomutase and phosphoglucoisomerase. The increase in levels of fructose 6-phosphate, the substrate of phosphofructokinase, suggests that an increase in the rate of glycolytic flux will follow, as it does in many cell types. On the contrary, hypertrehalosaemic hormone decreases glycolytic flux in the fat body, while simultaneously increasing trehalogenesis and the oxidation of fatty acids (Wiens and Gilbert, 1965, 1967; McDougall and Printed in Great Britain © The Company of Biologists Limited 1998
JEB1382
In cockroach fat body, trehalogenesis and glycolysis compete for glucose phosphates as common substrates. During trehalogenesis, glycolysis is inhibited, although the mechanism responsible for this is not known. Incubation of the isolated fat body from the Argentine cockroach Blaptica
dubia with an extract of the corpora cardiaca containing as
little as 0.005 gland equivalents ml−1of incubation medium increased the release of trehalose (anthrone-positive material) from the tissue by more than 100 %. The content of the glycolytic activator fructose 2,6-bisphosphate was decreased by up to 50 %. A decapeptide was isolated from the corpora cardiaca of B. dubia and shown to be identical to the naturally occurring Blaberus discoidalis hypertrehalosaemic peptide (Bld HrTH), which was also found in the corpora cardiaca. Synthetic Bld HrTH at 2 nmol l−1 and above increased trehalose production and decreased the content of fructose 2,6-bisphosphate to the
same extent as did corpus cardiacum extract. The octapeptides Periplaneta americana cardioacceleratory hormones I and II (Pea CAH-I and Pea CAH-II) also had a significant effect on both parameters. Fructose 2,6-bisphosphate is a potent activator of phosphofructokinase from cockroach fat body if the enzyme is assayed at near-physiological concentrations of substrates and effectors. It is suggested that, because of the decrease in fructose 2,6-bisphosphate levels in the fat body, the activity of the key glycolytic enzyme phosphofructokinase is diminished. This can explain the inhibition of glycolytic flux by hypertrehalosaemic peptides which alters the balance of glucose metabolism in favour of trehalose formation.
Key words: hypertrehalosaemic peptide, Blaberus discoidalis hypertrehalosaemic hormone, Bld HrTH, trehalose, fructose 2,6-bisphosphate, glycolysis, fat body, cockroach, Blaptica dubia. Summary
Introduction
HYPERTREHALOSAEMIC NEUROPEPTIDES DECREASE LEVELS OF THE
GLYCOLYTIC SIGNAL FRUCTOSE 2,6-BISPHOSPHATE IN COCKROACH FAT BODY
ANDREAS BECKER ANDGERHARD WEGENER*
Institut für Zoologie, Johannes Gutenberg-Universität, Saarstrasse 21, D-55099 Mainz, Germany
*Author for correspondence (e-mail: [email protected])
Steele, 1988). In many cells and tissues, phosphofructokinase has been established as a key enzyme in the control of glycolytic flux. However, the mechanism by which glycolysis is inhibited during trehalogenesis in fat body is not understood.
The analogy that can be drawn between insect fat body and vertebrate liver suggested that it might be useful to investigate the mechanism by which glycolysis is inhibited during trehalogenesis to determine what differences and similarities might exist between the two systems. In this study, we show that the hypertrehalosaemic hormones bring about an increase in trehalose production by the fat body in concert with a decrease in the content of fructose 2,6-bisphosphate, and that the decrease in levels of this signal metabolite diminishes the activity of fat body phosphofructokinase in vitro.
Materials and methods Chemicals
Enzymes and substrates were obtained from Boehringer-Mannheim (Boehringer-Mannheim, Germany), fructose 2,6-bisphosphate from Sigma Chemie (Deisenhofen, Germany) and liquid nitrogen from Linde AG (Mainz-Kostheim, Germany). Other chemicals were purchased from E. Merck (Darmstadt, Germany), Roth (Karlsruhe, Germany) and Serva Feinbiochemica (Heidelberg, Germany) and were of analytical grade. Synthetic hypertrehalosaemic peptides were purchased from Bachem (Heidelberg, Germany; Bld HrTH) or Peninsula (Merseyside, England; Pea CAH-I and Pea CAH-II). Stock solutions of 1 mg ml−1(approximately 1 mmol l−1) were prepared
in 20 % acetic acid. For use, the stock solutions were diluted with incubation medium. The incubation medium for isolated fat bodies contained 140 mmol l−1NaCl, 5 mmol l−1 KCl, 2 mmol l−1
CaCl2, 2 mmol l−1MgCl2and 40 mmol l−1 Hepes/NaOH, pH 7.4.
Experimental animals
Adult male Argentine cockroaches Blaptica dubia (Blaberidae) were used for the experiments. The cockroaches were reared at 25–30 °C, approximately 60 % relative humidity and in a 12 h:12 h light:dark cycle. The cockroaches were fed Chappi dog chow (Effem, Verden/Aller, Germany) and water ad libitum.
Preparation of corpus cardiacum extract
Corpora cardiaca were transferred to incubation medium (10 corpora cardiaca ml−1) and sonicated for 30 s (Branson
Sonifier; microtip, position 3). The homogenate was heated in a waterbath at 80 °C for 5 min, followed by centrifugation at 10 000 g for 5 min. The supernatant was collected and stored at −20 °C. For use, the concentrated stock solution was diluted with incubation medium as required.
Incubation of fat bodies
The fat body was removed under incubation medium and separated into left and right lobes. Each lobe was preincubated as described by Steele et al. (1988) at 30 °C for 10 min in 20 ml glass vials containing 2 ml of incubation medium. Each lobe
was then transferred to a clean vial containing 2 ml of fresh medium, either with or without the test reagent. All incubations were performed in a shaking, thermostatted waterbath. For time course experiments, the fat bodies were incubated for 10, 20, 30 and 60 min. The incubation period for dose–response studies was 30 min.
Determination of trehalose levels
The reaction for anthrone-positive material was used to estimate the trehalose content of incubation media. More than 95 % of the carbohydrate released from fat body in vitro is trehalose (Steele et al. 1984; A. Becker and G. Wegener, unpublished data). Anthrone reagent, which reacts specifically with carbohydrate, was prepared as described by Roe (1955) and modified by Carroll et al. (1956). Samples of medium were stored at −20 °C prior to analysis.
Determination of fructose 2,6-bisphosphate levels After incubation, the fat body was blotted with filter paper to remove adhering medium and then frozen in liquid nitrogen. The tissue was weighed while still frozen and sonicated in 10 parts (v/w) of 200 mmol l−1 NaOH. The homogenate was
incubated at 80 °C for 5 min and then centrifuged at 10 000 g for 15 min. The clear infranatant, beneath a thick layer of fat, was collected with a syringe fitted with a thin needle, diluted with 9 volumes of 10 mmol l−1NaOH and stored for future use
at −20 °C. Fructose 2,6-bisphosphate was assayed by its ability to activate the pyrophosphate-dependent phosphofructokinase (PPi-PFK) from potato tubers (Van Schaftingen et al. 1982b;
Van Schaftingen, 1984) and identified by its high lability at low pH (10 min at pH 2).
Identification of hypertrehalosaemic peptide Extraction of corpora cardiaca
Corpora cardiaca from 20 adult male Blaptica dubia were collected in 500µl of distilled water, centrifuged at 10 000 g for 5 min, the supernatant was collected and the pellet was extracted by sonication in 200µl of 80 % methanol. After centrifugation, the methanolic and aqueous supernatants were combined and dried under nitrogen. The dried material was resuspended in methanol and, after further sonication and centrifugation, the supernatant was dried by vacuum centrifugation.
Reverse-phase high-performance liquid chromatography The sample was dissolved in 40µl of solvent A (95 % H2O,
5 % acetonitrile and 0.08 % trifluoroacetic acid) and resolved on an Applied Biosystems (Munich, Germany) HPLC fitted with a Vydac column (250 mm×1.6 mm, 218 TP, 5µm) at a flow rate of 150µl min−1and 37 °C. The elution was performed
by mixing solvent A with solvent B (20 % H2O, 80 %
recorded at 214 nm and the fractions containing peaks were collected.
Deblocking and sequencing of peptides
To deblock the peptide by removal of the pyroglutamate residue, half of the fraction which had the same elution time as synthetic Bld HrTH was dried by vacuum centrifugation, dissolved in 15µl of digestion buffer (100 mmol l−1 sodium phosphate buffer, pH 8.0, 10 mmol l−1Na2-EDTA, 5 mmol l−1
dithiothreitol, 5 % glycerol) containing 0.5µg of pyroglutamate aminopeptidase (E.C. 3.4.19.3, Boehringer-Mannheim). Synthetic Bld HrTH (50 pmol) was also carried through the procedure as a control. The samples were incubated for 18 h at 4 °C and then for 4 h at 25 °C. The peptides were freed of pyroglutamate aminopeptidase by HPLC, and the fractions of interest were sequenced using an Applied Biosystems 477A sequencer.
Purification and assay of phosphofructokinase Fat body was homogenised using a glass homogeniser with a Teflon pestle (Braun, Melsungen) in 5 volumes of medium containing (in mmol l−1): 30 imidazole buffer, pH 7.6, 250 sucrose, 2 EDTA, 30 β-mercaptoethanol, 1 fructose 6-phosphate, 3.5 glucose 6-phosphate and 1 phenylmethylsulphonylfluoride (PMSF). The tissue extract was fractionated with saturated (NH4)2SO4and further purified
by ion-exchange chromatography on Q Sepharose and by affinity chromatography on Blue Sepharose (Pharmacia) according to Wegener et al. (1990). The enzyme proved stable with respect to activity and kinetic properties when kept in 50 % glycerol at −80 °C. Enzyme activity was assayed at 25 °C as described by Wegener et al. (1987). One unit (U) of activity is equivalent to 1µmol of substrate transformed per minute. Protein was determined using the Bio-Rad assay based on the method of Bradford (1976) with bovine serum albumin as the standard.
Statistics
Data are expressed as the means ± standard error (S.E.M.). Significance was analysed using a two-sided Student’s t-test for paired samples, as described by Sokal and Rohlf (1973).
Results
Fructose 2,6-bisphosphate and trehalose production in vitro Preliminary experiments showed considerable variation between individual cockroaches with respect to the content of fructose 2,6-bisphosphate in the fat body and the rate of trehalose production by the tissue. To avoid the difficulties associated with this variation, all in vitro studies employed the paired tissue technique as described by Steele et al. (1988). In one set of experiments, in which both the lateral and contralateral fat body lobes were incubated under control conditions for 30 min, the concentrations of fructose 2,6-bisphosphate in the two lobes were 2.14±0.26 and 2.06±0.23 nmol g−1, respectively. The corresponding values for trehalose production were 1.04±0.06 and 0.99±0.07µmol min−1g−1, respectively. For neither group (N=10) is the difference significant.
Effect of corpus cardiacum extract on trehalose and fructose 2,6-bisphosphate levels
Treatment of the fat body with extract of the corpus cardiacum resulted in an increase in trehalose production and a concomitant decrease in the content of fructose 2,6-bisphosphate (Table 1). The data suggested that a significant effect on both metabolites was obtained with a concentration of 0.005 corpus cardiacum equivalents per millilitre. The results also suggested that higher concentrations of the extract do not augment either of the responses.
Isolation of hypertrehalosaemic neuropeptide
[image:3.609.50.566.551.653.2]The hypertrehalosaemic action of the corpus cardiacum
Table 1. Effect of aqueous corpora cardiaca extract on trehalose production and the content of fructose 2,6-bisphosphate of isolated fat body lobes from Blaptica dubia
Corpora cardiaca Rate of trehalose production Fructose 2,6-bisphosphate content
extract (gland (µmol min−1g−1) (nmol g−1)
equivalents ml−1) N Control Experimental Change P Control Experimental Change P
0.001 5 0.75±0.23 0.99±0.30 +0.24±0.10 NS 1.22±0.14 0.93±0.09 −0.29±0.19 NS
0.005 5 0.63±0.06 1.38±0.08 +0.75±0.10 0.01 1.39±0.13 0.79±0.09 −0.60±0.14 0.02
0.01 8 0.72±0.06 1.49±0.13 +0.80±0.14 0.001 1.91±0.34 1.15±0.25 −0.76±0.17 0.01
0.05 5 0.51±0.10 1.08±0.18 +0.57±0.09 0.01 2.39±0.48 1.68±0.33 −0.71±0.17 0.02
0.1 6 0.62±0.05 1.34±0.15 +0.73±0.11 0.01 1.62±0.07 0.79±0.11 −0.84±0.13 0.01
Fat body lobes were incubated in 2 ml of medium as described in Materials and methods. One fat body lobe was incubated in the presence of various doses of corpora cardiaca extract (experimental), while the other lobe from the same cockroach was incubated in medium only (control). After 30 min of incubation, the trehalose (anthrone-positive material) released by the fat body into the medium was measured, as was the content of fructose 2,6-bisphosphate in the tissue.
Values are given as of µmol trehalose per gram of fat body tissue per minute of incubation and as nmol of fructose 2,6-bisphosphate per gram of fat body and are means ±S.E.M. of N independent observations.
extract posed questions about the identity of the active factor. Furthermore, it was of interest to know whether the same factor was responsible for both the increase in trehalose release and the decrease in fructose 2,6-bisphosphate concentration. Reverse-phase HPLC of a methanolic extract of the corpora cardiaca of Blaptica dubia produced a prominent peak corresponding to the peak generated when synthetic Bld HrTH was run under identical conditions (Fig. 1). The peptide fraction corresponding to the Bld HrTH peak was isolated, treated with pyroglutamate aminopeptidase and sequenced. The sequence was found to be identical to that of Bld HrTH.
Effect of synthetic hypertrehalosaemic peptides on trehalose release and fructose 2,6-bisphosphate levels
It is well established that the hypertrehalosaemic effect of corpus cardiacum extract is due to the action of neuropeptides in the extract. The important question is whether the decrease in the fat body fructose 2,6-bisphosphate content is also
induced by these peptides. This was tested by incubating fat bodies with three synthetic hypertrehalosaemic neuropeptides: Bld HrTH, the naturally occurring peptide of Blaptica dubia, and the two octapeptides Pea CAH-I and Pea CAH-II, first identified in Periplaneta americana (Scarborough et al. 1984; for a review, see Becker et al. 1996).
Each of the peptides stimulated trehalose production and decreased the content of fructose 2,6-bisphosphate, although they differed with respect to their maximal effects and efficiency (Table 2). The decapeptide Bld HrTH was more potent than the octapeptides. While trehalose production was increased by 130±45 % (mean of the percentage changes ±
S.E.M., N=5) by 10 nmol of Bld HrTH, the increase due to 10 nmol of Pea CAH-I was only 91±21 % (N=6) and that due to 10 nmol of Pea CAH-II was only 26±2 % (N=6, not significant). At 10 nmol, each peptide also significantly decreased the content of fructose 2,6-bisphosphate in the isolated fat body by 37±4 % (N=5) for Bld HrTH, by 28±7 % (N=6) for Pea CAH-I and by 15±3 % (N=6) for Pea CAH-II.
The time-dependent effects of Bld HrTH on trehalose production and on the content of fructose 2,6-phosphate in isolated fat body were determined using the lowest concentration of hormone (10 nmol) that had been found to give a maximal response in the dose–response study. A significant effect on both metabolites occurred after 10 min of incubation. The increase in trehalose release due to the hormone was
Retention time (min)
0 10 20 30 40 50 60 70 80
Absorbance at 214 nm
(arbitrary units)
% Solvent B
0 10 20 30 40 50 60 70 80 90 100 0 10 20 30 40 50 60 70 80 90 100
50 pmol of Bld HrTH % Solvent B A
B
[image:4.609.318.549.75.286.2]Corpora cardiaca extract (5 CC in 10 µl) % Solvent B
Fig. 1. Reverse-phase HPLC of (A) the synthetic hypertrehalosaemic neuropeptide Bld HrTH and (B) an extract of corpora cardiaca (CC) from Blaptica dubia. Both fractionations were run under identical conditions as described in Materials and methods. The arrow in B marks the position corrresponding to the Bld HrTH peak in A.
Time (min)
0 10 20 30 40 50 60
Trehalose production (
µ
mo
l g
−
1)
0 20 40 60 80
***
***
***
***
10−8 mol l−1 Bld HrTH
Control
[image:4.609.55.284.75.438.2]79±17 % (N=9) at 10 min, increasing to 124±22 % (N=9) after 30 min of incubation (Fig. 2). In both the control tissue and tissue treated with hormone, 50 % of the total trehalose produced during incubation for 60 min was produced during the first 20 min. The non-linearity of the response and the relatively high rate of trehalose production by the control tissues reflect the fact that there is no feedback inhibition of trehalose synthesis at the beginning of incubation in the in vitro system as there is in vivo, where the concentration of trehalose in the haemolymph is high. The hormone decreased fructose 2,6-bisphosphate levels in the tissue by as much as 40 % during the first 10 min of incubation, and this difference was maintained during the remainder of the incubation period (Fig. 3).
Modulation of phosphofructokinase activity by fructose 2,6-bisphosphate
The optimal activity of phosphofructokinase in Blaptica dubia fat body was 1.36±0.06µmol min−1g−1at 25 °C (N=10). The enzyme was purified approximately 1000-fold to a specific activity of 24 U mg−1protein. Optimal activity of the enzyme was obtained using 2 mmol l−1 fructose 6-phosphate, 2 mmol l−1 ATP, 1 mmol l−1 AMP and 10 mmol l−1inorganic phosphate (Pi) at pH 7.2. To evaluate the effect of fructose
[image:5.609.46.568.96.338.2]2,6-bisphosphate on the activity of phosphofructokinase, the enzyme was assayed at near-physiological concentrations of substrates and effectors. Under these conditions, the enzyme proved very sensitive to changes in fructose 2,6-bisphosphate level in the physiological range (Fig. 4).
Table 2. Effect of various concentrations of three synthetic hypertrehalosaemic neuropeptides on trehalose production and fructose 2,6-bisphosphate content of fat body isolated from Blaptica dubia
Peptide Rate of trehalose production Fructose 2,6-bisphosphate content
concentration (µmol min−1g−1) (nmol g−1)
(mol l−1) N Control Experimental Change P Control Experimental Change P
Bld HrTH
10−10 5 0.71±0.02 0.72±0.09 +0.01±0.09 NS 2.99±0.49 2.75±0.39 −0.24±0.12 NS
5×10−10 5 1.16±0.18 1.34±0.22 +0.17±0.08 NS 3.18±0.28 2.66±0.21 −0.52±0.21 NS
10−9 5 0.78±0.08 0.97±0.08 +0.10±0.07 NS 1.28±0.10 1.01±0.08 −0.26±0.04 0.001
2×10−9 5 0.86±0.10 1.38±0.18 +0.52±0.09 0.01 2.86±0.29 1.98±0.39 −0.88±0.19 0.01 5×10−9 5 0.90±0.06 1.92±0.18 +1.02±0.22 0.01 2.49±0.16 1.68±0.27 −0.80±0.14 0.01
10−8 5 0.55±0.14 1.06±0.14 +0.50±0.03 0.001 1.80±0.37 1.12±0.23 −0.68±0.17 0.02
10−7 5 0.43±0.06 0.95±0.09 +0.52±0.06 0.001 1.53±0.28 0.89±0.27 −0.64±0.16 0.02
Pea CAH-I
10−9 4 1.09±0.35 1.01±0.28 −0.08±0.22 NS 1.65±0.49 1.85±0.56 +0.20±0.09 NS
10−8 6 0.73±0.06 1.35±0.20 +0.69±0.16 0.01 1.67±0.08 1.15±0.13 −0.53±0.11 0.01
10−7 6 0.69±0.08 1.23±0.10 +0.54±0.07 0.001 2.30±0.40 1.60±0.30 −0.71±0.12 0.002
Pea CAH-II
10−9 8 1.39±0.14 1.31±0.16 −0.08±0.07 NS 1.70±0.29 1.93±0.35 +0.23±0.14 NS
10−8 6 0.88±0.12 1.11±0.16 +0.23±0.09 NS 2.11±0.22 1.79±0.13 −0.32±0.10 0.05
10−7 6 0.77±0.08 1.21±0.15 +0.44±0.14 0.05 2.37±0.40 1.80±0.28 −0.57±0.16 0.02
Fat bodies were incubated for 30 min as described in Table 1. Values are means ±S.E.M. of N independent observations.
The significance of a change is indicated by the P value; NS, not significant.
Fig. 3. Time-dependent effect of hypertrehalosaemic peptide on the content of fructose 2,6-bisphosphate in isolated fat body from Blaptica dubia. Paired fat body lobes from the same animal were incubated for 10, 20, 30 or 60 min in 2 ml of medium either in the absence (control) or in the presence of 10 nmol l−1 Bld HrTH (experimental). Values are expressed as nmol of fructose 2,6-bisphosphate per gram of fat body tissue and are means + S.E.M. of
six independent observations. Asterisks indicate a significant difference from the control value; *P<0.05, **P<0.01.
Time (min)
[Fructose 2,6-bisphosphate] (nmol g
−
1)
0 1 2 3 4
**
*
*
**
10 20 30 60 Control
[image:5.609.334.546.406.612.2]Discussion
Our data show that the hypertrehalosaemic neuropeptide Bld HrTH, first isolated from the corpora cardiaca of Blaberus discoidalis and Nauphoeta cinerea (Gäde and Rinehart, 1986; Hayes et al. 1986), is also present in the corpora cardiaca of Blaptica dubia, where it probably functions as a hypertrehalosaemic hormone in this species. The results are in agreement with the suggestion that Bld HrTH is the natural peptide in cockroaches belonging to the families Blaberidae and Blattelidae, while the octapeptides Pea CAH-I and Pea CAH-II are found in the Blattidae (Gäde, 1989, 1996). The findings that, of the three peptides tested, Bld HrTH had the most potent effect on increasing trehalose production and decreasing the content of fructose 2,6-bisphosphate also support this view. Not surprisingly, the greater the dissimilarity between the peptide and the natural hormone Bld HrTH the lower was the potency. Pea CAH-I, which is identical to Bld
HrTH in the first six amino acids, had a higher potency with respect to both parameters than did Pea CAH-II with only four identical positions.
Stimulation of trehalogenesis in the isolated fat body by Bld HrTH is accompanied by a concomitant decrease in the content of fructose 2,6-bisphosphate. This effect is highly significant and suggests that fructose 2,6-bisphosphate is involved in the hypertrehalosaemic action of the hormone. The only established functions of fructose 2,6-bisphosphate are those of an activator of phosphofructokinase and an inhibitor of fructose 1,6-bisphosphatase. In vivo, both enzymes are far removed from equilibrium and are regarded as key enzymes in the regulation of glycolysis and gluconeogenesis. Although the regulatory properties of these enzymes from the fat body have not been characterized in detail, our data show that fructose 2,6-bisphosphate is a potent activator of phosphofructokinase (Fig. 4) as well as being an inhibitor of fructose 1,6-bisphosphatase (Becker et al. 1996; Kasprzyk, 1993).
Our data show that the observed decrease in the content of fructose 2,6-bisphosphate brings about a decrease in the activity of phosphofructokinase in vitro (see Fig. 4). However, the decrease in fructose 2,6-bisphosphate level would be physiologically significant only if a change in the levels of effectors other than fructose 2,6-bisphosphate did not offset the effect of a decrease in fructose 2,6-bisphosphate level. In practice, corpus cardiacum extract increases the concentration of fructose 6-phosphate and also slightly increases that of the activator AMP, while decreasing the concentration of the inhibitor ATP (Sevala and Steele, 1991). The latter changes would increase rather than decrease the activity of phosphofructokinase. However, we have observed a marked increase in the concentration of the glycolytic (gluconeogenic) intermediate fructose 1,6-bisphosphate in fat body from 8.9µmol l−1to 58.5µmol l−1 after 1 h in the presence of 10−8mol l−1Bld HrTH (A. Becker and G. Wegener, unpublished data). Fructose 1,6-bisphosphate has a negligible effect on the activity of phosphofructokinase in the absence of fructose 2,6-bisphosphate, but strongly reduces the activating effect of the latter (Wegener et al. 1987). Kinetic data and binding studies for phosphofructokinase from locust flight muscle and other animal tissues indicate that fructose 1,6-bisphosphate and fructose 2,6-bisphosphate compete for the same regulatory site on the enzyme, so that fructose 1,6-bisphosphate would lower phosphofructokinase activity by displacing the potent activator fructose 2,6-bisphosphate from its binding site on the enzyme (for references, see Wegener et al. 1987). Thus, a hormone-induced increase in fructose 1,6-bisphosphate level from 10µmol l−1to 60µmol l−1will further decrease the activity of phosphofructokinase under the conditions shown in Fig. 4.
At present, we do not know whether the hypertrehalosaemic hormone generates fructose 1,6-bisphosphate via glycolysis (phosphofructokinase) or gluconeogenesis (reverse aldolase reaction). In either case, an
0 2 4 6 8 10 0
5 10 15 20 25 30
[Fructose 2,6-bisphosphate] (µmol l−1)
Phosphofructokinase activity (m
U m
l
−
1)
Fructose 2,6-bisphosphate concentration in fat body
[image:6.609.59.275.74.286.2]
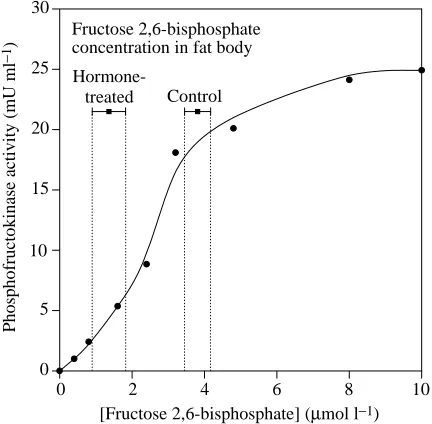
Hormone-treated Control
Fig. 4. Simulation in vitro of the effect of the hormone-induced decrease in fructose 2,6-bisphosphate level on phosphofructokinase activity in fat body. Phosphofructokinase was purified from the fat body of Blaptica dubia, and the effect of fructose 2,6-bisphosphate on the activity of phosphofructokinase was assayed at near-physiological concentrations of substrates and effectors. The bars above the curve indicate the mean concentrations ± S.E.M. of fructose 2,6-bisphosphate in control fat body lobes and in lobes exposed to 10 nmol l−1 Bld HrTH for 60 min. To calculate the concentration of fructose 2,6-bisphosphate in fat body, we assumed the intracellular waterspace to be 58.7 % (as determined for Periplaneta americana according to the method used by Sevala and Steele, 1991) and that fructose 2,6-bisphosphate was completely solubilized in the cytosol. Assay conditions: 20µmol l−1fructose 6-phosphate, 1.5 mmol l−1 ATP, 1 mmol l−1 Pi, 3.5µmol l−1 AMP,
10µmol l−1 fructose 1,6-bisphosphate, pH 7.2. Vopt=40.2 mU ml−1
(for optimal assay conditions, see Results), Vmax=25 mU ml−1; an
increase in fructose 1,6-bisphosphate concentration during trehalogenesis is compatible with a decrease in phosphofructokinase activity.
Little is known about the mechanism employed by the hypertrehalosaemic hormones to lower the level of fructose 2,6-bisphosphate in the fat body (see Becker et al. 1998). In liver, fructose 2,6-bisphosphate is synthesized from and degraded to fructose 6-phosphate by a specific bifunctional enzyme system 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase (PFK-2/FBPase-2; for a review, see Van Schaftingen, 1990). The relative activities of these two enzymes thus determine the content of fructose 2,6-bisphosphate in a tissue. Because both enzymes catalyse non-equilibrium reactions, their activities must be regulated, i.e. controlled by factors other than the concentrations of their substrates. This idea is supported by studies on the cockroach fat body showing that, during trehalogenesis, the concentration of fructose 6-phosphate (the substrate of PFK-2 and an inhibitor of FBPase-PFK-2) is increased (Steele et al. 1984; Sevala and Steele, 1991) yet fructose 2,6-bisphosphate concentration, as shown in this study, is decreased, i.e. during the transition from the non-stimulated to the stimulated state, the activity of FBPase-2 in the fat body must exceed that of PFK-2. The way in which these enzymes are regulated in the cockroach fat body remains to be investigated. In liver, the activities of PFK-2/FBPase-2 are controlled in a complex manner by counteracting mechanisms involving hormones, interconversions by phosphorylation/dephosphorylation and allosteric effectors (for a review, see Van Schaftingen, 1990).
We conclude from our study that the hypertrehalosaemic neuropeptides decrease the content of fructose 2,6-bisphosphate in the fat body and that this is part of the mechanism for the inhibition of glycolysis at the phosphofructokinase step. This would enable the concentration of glucose phosphates to increase and thus favour the synthesis of trehalose.
This work was supported by grants from the Deutsche Forschungsgemeinschaft, D-53175 Bonn, Germany. We thank Dr R. Kellner (Proteinchemisches Zentrallabor, Institut für Physiologische Chemie, Johannes Gutenberg-Universität Mainz) for his help with peptide sequencing, Mrs Heike Stypa for expert technical assistance and Dipl. Biol. Jana F. Liewald for help with the preparation of the manuscript.
References
BECKER, A., LIEWALD, J. F. AND WEGENER, G. (1998). Signal
transduction in isolated fat body from the cockroach Blaptica dubia exposed to hypertrehalosaemic neuropeptide. J. comp. Physiol. B (in press).
BECKER, A., SCHLÖDER, P., STEELE, J. E. ANDWEGENER, G. (1996).
The regulation of trehalose metabolism in insects. Experientia 52, 433–439.
BRADFORD, M. M. (1976). A rapid and sensitive method for
quantitation of microgram quantities of proteins utilizing the principle of protein–dye binding. Analyt. Biochem. 72, 248–254.
CARROLL, N. V., LONGLEY, R. W. AND ROE, J. H. (1956). The
determination of glycogen in liver and muscle by the use of anthrone reagent. J. biol. Chem. 220, 583–593.
GÄDE, G. (1989). The hypertrehalosaemic peptides of cockroaches: a phylogenetic study. Gen. comp. Endocr. 75, 287–300.
GÄDE, G. (1996). The revolution in insect neuropeptides illustrated by the adipokinetic hormone/red pigment-concentrating hormone family of peptides. Z. Naturforsch. 51c, 607–617.
GÄDE, G. ANDRINEHART, K. L., JR(1986). Amino acid sequence of a
hypertrehalosaemic neuropeptide from the corpus cardiacum of the cockroach, Nauphoeta cinerea. Biochem. biophys. Res. Commun. 141, 774–781.
HAYES, T. K., KEELEY, L. L. AND KNIGHT, D. W. (1986). Insect
hypertrehalosaemic hormone: Isolation and primary structure from
Blaberus discoidalis cockroaches. Biochem. biophys. Res.
Commun. 140, 674–678.
KASPRZYK, A. (1993). Fructose-1,6-bisphosphatase in the fat body of
the cockroach Periplaneta americana larvae. Acta biochim. polon. 40, 130–132.
MCDOUGALL, G. E. ANDSTEELE, J. E. (1988). Free fatty acids as a source of energy for trehalose synthesis in the fat body of the American cockroach (Periplaneta americana). Insect Biochem. 18, 591–597.
PILKIS, S. J. (1990). (ed.) Fructose 2,6-bisphosphate. Boca Raton, FL: CRC Press.
ROE, J. H. (1955). The determination of sugar in blood and spinal fluid with anthrone reagent. J. biol. Chem. 212, 335–343.
SCARBOROUGH, R. M., JAMIESON, G. C., KALISH, F., KRAMER, S. J., MCENROE, G. A., MILLER, C. A. AND SCHOOLEY, D. A. (1984).
Isolation and primary structure of two peptides with cardioacceleratory and hyperglycaemic activity from the corpora cardiaca of Periplaneta americana. Proc. natn. Acad. Sci. U.S.A. 81, 5575–5579.
SEVALA, V. L. AND STEELE, J. E. (1991). Regulation of glycolytic intermediates in cockroach fat body by the corpus cardiacum. J. comp. Physiol. B 161, 349–355.
SOKAL, R. R. ANDROHLF, F. J. (1973). Introduction to Biostatistics.
San Francisco, CA: W. H. Freeman.
STEELE, J. E. (1961). Occurrence of a hyperglycemic factor in the
corpus cardiacum of an insect. Nature 192, 680–681.
STEELE, J. E. (1963). The site of action of insect hyperglycemic
hormone. Gen. comp. Endocr. 3, 46–52.
STEELE, J. E., COULTHART, K. C. ANDMCCLURE, J. B. (1984). Control
of hexose phosphate and citrate in fat body of the cockroach (Periplaneta americana) by the corpus cardiacum. Comp. Biochem. Physiol. 79B, 559–563.
STEELE, J. E., MCDOUGALL, G. E. AND SHADWICK, R. (1988).
Trehalose efflux from cockroach fat body in vitro: paradoxical effects of the corpus cardiacum and methylxanthines. Insect Biochem. 18, 585–590.
VAN SCHAFTINGEN, E. (1984). D-Fructose 2,6-bisphosphate. In
Methods of Enzymatic Analysis, vol. VI (ed. H. U. Bergmeyer), pp. 335–341. Weinheim: Verlag Chemie.
VANSCHAFTINGEN, E. (1990). Role of fructose-2,6-bisphosphate in the regulation of hepatic carbohydrate metabolism. In Fructose-2,6-bisphosphate (ed. S. J. Pilkis), pp. 65–85. Boca Raton, FL: CRC Press.
Fructose-2,6-bisphosphatase from rat liver. Eur. J. Biochem. 124, 143–149.
VANSCHAFTINGEN, E. ANDHERS, H. G. (1981). Inhibition of fructose-1,6-bisphosphatase by fructose 2,6-bisphosphate. Proc. natn. Acad. Sci. U.S.A. 78, 2861–2863.
VANSCHAFTINGEN, E., HUE, L. ANDHERS, H. G. (1980). Fructose
2,6-bisphosphate, the probable structure of the glucose- and glucagon-sensitive stimulator of phosphofructokinase. Biochem. J. 192, 897–901.
VANSCHAFTINGEN, E., LEDERER, B., BARTRONS, R. ANDHERS, H. G.
(1982b). A kinetic study of pyrophosphate: fructose-6-phosphate phosphotransferase from potato tubers. Eur. J. Biochem. 129, 191–195.
WEGENER, G., BEINHAUER, I., KLEE, A. AND NEWSHOLME, E. A. (1987). Properties of locust muscle 6-phosphofructokinase and their importance in the regulation of glycolytic flux during prolonged flight. J. comp. Physiol. B 157, 315–326.
WEGENER, G., KRAUSE, U. AND THUY, M. (1990). Fructose 2,6-bisphosphate and glycolytic flux in skeletal muscle of swimming frog. FEBS Lett. 267, 257–260.
WIENS, A. W. ANDGILBERT, L. I. (1965). Regulation of cockroach
fat-body metabolism by the corpus cardiacum in vitro. Science 150, 614–616.