Introduction
Colorectal adenocarcinoma is the most com-mon form of colonic cancer affecting approxi-mately 112,000 new patients every year. Colo-rectal cancer accounted for approximately 19% of all cancer-related deaths in the United States in 2007 [1]. Colorectal adenocarcinoma affects patients usually older then 40 years, except in individuals with genetic predisposition to this form of cancer [2]. The patients with high stage tumors are those that frequently develop metas-tases and succumb to the cancer [3].
Sporadic colorectal cancer usually develops following the accumulation of multiple sequen-tial genetic changes within a cell. While somatic mutations of the APC tumor suppressor gene are the first step toward carcinogenesis, the accumulation of other sequential genetic or epigenetic events activate oncogenes (Ras,
c-(DCC, DPC-4, P53, and others) [4-9]. These ge-netic changes are thought to be translated in functional alterations that eventually provide the tumor cell with new malignant attributes such as increase mobility, capability of invading the surrounding stroma, of evading the immune system, and of metastasizing. Recently, the “serrated pathway” was identified as another pathway responsible for colon carcinogenesis. This pathway involves errors in mismatch repair genes and involvement of cyclin B, Braf, TGFBR2, and others [10]
It has been shown that inhibition of apoptosis is critical to colorectal Tumorigenesis [11]. For example, it has been proposed that overexpres-sion of Bcl-XL in cancer may suppress the activ-ity of the proapoptotic molecules Bax and Bak, contributing to cancer progression [12, 13]. It seems that, also in CRC, the dissociation of Bax and Bcl-XL promotes Bax multimerization and
Original Article
Correlation between Mcl-1 and pAKT protein expression in
colorectal cancer
Evita B. Henderson-Jackson6, James Helm4, Masoumeh Ghayouri1,2,3, Ardeshir Hakam1,2, Aejaz Nasir2,
Marino Leon1,2,3, Marilyn Bui1,2,5, Timothy Yeatman1,2,3, Domenico Coppola1,2,3,5
Departments of 1Oncologic Sciences; 2Anatomic Pathology; 3Gastrointestinal Oncology; 4Biostatistic; 5Program of
Experimental Therapeutics; Moffitt Cancer Center and Research Institute; and 6Department of Pathology, at the
Uni-versity of South Florida, School of Medicine; Tampa, Florida, USA.
Received September 25, 2010; accepted October 11, 2010; available online October 12, 2010
Abstract: Mcl-1 inhibits apoptosis in well-differentiated cells by sequestering BAD, BID, and BAX and other apoptotic molecules. pAKT blocks apoptotsis by facilitating the interaction of BAD with BCL-XL. Expression of pAKT and Mcl-1 have been described in colon cancer, however, the relationship between pAKT and Mcl-1 has not. Mcl-1 and pAKT immunohistochemistry was performed using colorectal cancer tissue microarray (TMA). The Holm step-down method was used to adjust for multiple testing. Mcl-1 and pAKT scores, stage, and grade were compared using Spearman’s correlation coefficient. Metastasis and no metastasis groups were compared using the Wilcoxon rank sum test. Mcl-1 and pAKT scores were compared for normal colorectal mucosa (NR), adenoma (AD), and colorectal cancer (CRC) cohorts. The mean (SD) pAKT expression in NR (14) was 2.0 (1.4), in AD (8) was 3.0 (1.7), and in CRC (101) was 5.6 (2.4). These differences were statistically significant. For Mcl-1 the mean (SD) expression was 4.1 (1.7) in NR, 3.2 (1.2) in AD, and 3.3 (2.6) in CRC. Mcl-1 and pAKT scores were directly correlated during various stages of colon car-cinogenesis (p = 0.04). Mcl-1 showed direct correlation with tumor grade (p = 0.001) and tumor stage (p = 0.02) and with presence of metastasis (p = 0.008). We report the correlation of Mcl-1 protein expression with higher grade and stage in colorectal cancer. Mcl-1 correlated also with pAKT expression. We also report the up regulation of pAKT dur-ing the transition from NR to CRC.
tosis [14].
Mcl-1 (myeloid cell leukemia-1) is a Bcl-2 family protein that interferes with mitochondrial activa-tion to inhibit apoptosis. Altered expression pat-tern of Mcl-1, as well as of Bax and Bcl-XL, has been described during colorectal cancer pro-gression [11-13, 16]. Backus et al. have de-scribed the interesting co-localization of Bax, Mcl-1 and Bcl-XL reactivity to the apical areas of the normal intestinal mucosa, as opposed to the diffuse cytoplasmic staining in the tumor cells [11].
IGF1-dependent activation of AKT effects prolif-eration, transformation, resistance to apoptosis, and metastatic potential of colon cancer cells [17]. The indication that AKT activation has a pivotal role in colorectal carcinogenesis also derives from the observation that mice lacking the catalytic subunit of PI3 kinase gamma de-velop spontaneous intestinal adenocarcinomas [18]. It is known that loss of PTEN protein acti-vates phosphoinositol (PI)-3 kinase, with gen-eration of PI 3,4,5-triphosphate and recruite-ment and activation of AKT to the plasma mem-brane [19]. It has become evident that AKT acti-vation facilitates cell transformation and tumori-genesis affecting multiple pathways regulating not only apoptosis [20], but also the cell cycle [21], cell motility [22], and angiogenesis [23].
Studies on clinical samples have show that AKT activation is increased in 46% of colorectal car-cinomas, and its association with Ki-67 prolif-eration index and inversely associated with the presence of apoptosis [24]. Others have shown that AKT activation increases also during the transition from benign polyps to carcinoma, and that such activation is inversely correlated to PTEN expression, a tumor suppressor protein known to inhibit the activation of PI3K/AKT pathway in colon cancer [25]. Despite the simi-lar antiapoptotic function of Mcl-1 and pAKT, we found no reports in the current English literature on their correlation in colorectal cancer.
In this study, using stage oriented human colo-rectal cancer tissue microarrays; we evaluated changes in Akt activity and in Mcl-1 expression during the progression from normal colonic mu-cosa to adenoma, to invasive colorectal adeno-carcinoma, using immunohistochemistry. The results were correlated with clinical pathologic findings.
Materials and methods
Selection of cases
Using stage oriented human colorectal cancer tissue microarrays (prepared in the Histology laboratory of the Moffitt Cancer Center Tissue Core Facility), 123 tissue samples (101 CRC, 8 adenomas, and 14 samples of normal colonic mucosa) were analyzed for Mcl-1 expression and pAKT activation by immunohistochemistry. All of the tumors used for the tissue array con-struction were CRC identified from the Moffitt Cancer Center, Anatomic Pathology Depart-ment’s database, CoPath®, and representing surgical resection specimens obtained between 1990 and 2002. All of the specimens were pre-served in 10% buffered formalin, prior to em-bedding in paraffin, for at least 12 hours. The tumors were staged according to the TNM sys-tem, following the recommendations of the American Joint Committee on Cancer, 1988. The stage of the invasive tumors was as follows: 12 patients had stage I, 37 stage II, 43 stage III, and 9 stage IV disease. All tumors occurred in the absence of genetic cancer syndromes such as human non polyposis colon cancer syndrome (HNPCC), familial adenomatous polyposis syn-drome (FAP), etc.; also cancers arising in the background of ulcerative colitis or Crohn’s dis-ease were excluded from the study. The NR samples were taken near the resected colorec-tal margin, away from the tumor site, from pa-tients that underwent colon resection for CRC.
Immunohistochemistry
minutes. After blocking with universal blocking serum (Ventana Medical Systems, Inc., Tucson, Arizona OMNIMAP) for 30 minutes, the samples were incubated with the primary antibodies at 4° C overnight. The samples were then incu-bated with biotin-labeled secondary antibody and streptavidin-horseradish peroxidase for 30 minutes each (Ventana Medical Systems). The slides were developed with 3,3’-diaminobenzidine tetrahydrochloride substrate (Ventana Medical Systems Inc. Tucson, Arizona) and counterstained with hematoxylin (Ventana Medical Systems Inc. Tucson, Arizona). The tissue samples were dehydrated and coversli-ped. Standard cell conditioning (following the Ventana proprietarian recommendations) was used for antigen retrieval. The specificity of the anti-Mcl-1 and anti-pAKT monoclonal antibodies was confirmed by immunostains of colon cancer cell lines constitutively expressing Mcl-1 and pAKT antibodies, and colon cancer cell lines KO for Mcl-1 and pAKT [12, 13]. Negative control was included by using non immune mouse sera and omitting the primary antibodies during the primary antibody incubation step.
Immunohistochemical data analysis
The Mcl-1 and pAKT stained tissue cores were examined by two independent observers (EA, DC); and a consensus score was reached for each specimen. The positive reaction for both antibodies was scored into four grades, accord-ing to the intensity of the stainaccord-ing: 0, 1+, 2+, and 3+. The percentages of Mcl-1 or pAKT posi-tive cells were also scored into four categories: 0 (0%), 1 (1-33%), 2 (34-66%), and 3 (67-100%). The product of the intensity by percent-age scores was used as the final score. The final scores were classified as: 0 negative; 1-3, weak; 4-6, moderate; and 7-9, strong. The specimens were also classified by the types of tissue staining positive: normal colonic mucosa, adenoma, and adenocarcinoma.
Statistical analysis
Descriptive statistics for the scores were gener-ated and reported for each tissue group. Mcl-1 and pAKT scores were compared for normal (N), adenoma (AD), and adenocarcinoma (CRC) co-horts. The Holm step-down method was used to adjust for multiple testing. Mcl-1 and pAKT score, stage, and grade were compared using Spearman’s correlation coefficient. Mcl-1 and pAKT expression scores for metastasis and no metastasis groups were compared using the
Wilcoxon rank sum test. The tests were called significant at the significance level of 0.05.
Results
Clinical pathologic findings
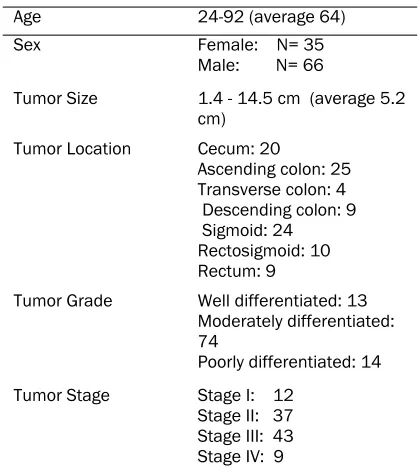
The clinical pathological findings are summa-rized in Table 1. The patients with cancer had an average age of 64 years (range, 24-92). Sixty -six were male and 35 were female. All the tu-mors occurred in the absence of familial adeno-matous polyposis or HNPCC. The tumors ranged in size between 1.4 cm. and 14.5 cm (average 5.2 cm), mostly polypoid and ulcerated. Twenty tumors involved the cecum, 25 the ascending colon, 4 the transverse colon, 9 the descending colon, 24 the sigmoid colon, 10 the rectosig-moid junction, and 9 the rectum. Thirteen tu-mors were well differentiated, 74 were moder-ately differentiated, and 14 were poorly differen-tiated. Of the invasive tumors, 12 were Duke’s stage A, 37 Duke’s stage B, 43 Duke’s stage C, and 9 Duke’s stage D. Only 2 patients, both with rectal cancer, received preoperative radiation, to reduce the size of their tumors.
Immunohistochemical results
[image:3.612.323.532.97.334.2]All of the Mcl-1 positively stained cases had nuclear staining. The pAKT stain was predomi-nantly cytoplasmic, however in some cases nu-clear and cytoplasmic stain was observed. The
Table 1. Clinical pathological findings
Age 24-92 (average 64) Sex Female: N= 35
Male: N= 66
Tumor Size 1.4 - 14.5 cm (average 5.2 cm)
Tumor Location Cecum: 20
Ascending colon: 25 Transverse colon: 4 Descending colon: 9 Sigmoid: 24 Rectosigmoid: 10 Rectum: 9
Tumor Grade Well differentiated: 13 Moderately differentiated: 74
Poorly differentiated: 14 Tumor Stage Stage I: 12
cytoplasmic staining was diffusely granular with variation in intensity seen within the same le-sion of some cases. Cases with variable stain-ing were graded based on the predominant staining intensity and the percentage of positive stain was determined based on the amount of the lesion demonstrating the predominant in-tensity. For Mcl-1 the mean (SD) expression was 4.1 (1.7) in NR, 3.2 (1.2) in AD, and 3.3 (2.6) in CRC (Figure 1A, B, C). For pAKT the mean (SD) pAKT expression in NR (14) was 2.0 (1.4), in AD (8) was 3.0 (1.7), and in CRC (101) was 5.6 (2.4) (Figure 1D, E, F). Despite the fact that a decrease in Mcl-1 staining was observed between NR vs. AD and CRC, the presence of staining within CRC cases correlated with grade and stage.
Statistical analysis
The statistical analysis results are summarized in Table 2. A Wilcoxon rank sum test revealed a statistically significant difference in pAKT stain-ing score between NR and CRC value<0.0001), and between AD and CRC value = 0.006), but not between AD and NR (p-value = 0.61). The same test did not detect
sig-nificant correlation in Mcl-1 staining scores be-tween NR and CRC (p-value = 0.61), AD and CRC (p = 0.76), and between AD and NR (p-value = 0.76). When considering the CRC co-hort, we observed a correlation between Mcl-1 expression and AKT activation (Spearman’s cor-relation coefficient 0.23, p-value = 0.04). Mcl-1 showed direct correlation with tumor grade (Spearman correlation coefficient = 0.36; p = 0.001) and tumor stage (Spearman correlation coefficient = 0.26; p = 0.02). Also, the Wilcoxon rank sum test showed a significant correlation of the Mcl-1 staining scores with presence of metastasis (p = 0.008), however, only 9 cases with metastases were available for evaluation. This finding needs to be re-evaluated using a larger cohort. pAKT staining score did not corre-late with grade, stage, or presence of metasta-ses.
Discussion
Programmed cell death (PCD) is defined as a physiological process that plays a critical role in normal development, cellular differentiation, and tissue homeostasis of multicellular organ-isms [26]. Dysregulation of this physiological
cell death process contributes to the patho-genesis of human diseases including cancer [26,27].
Mcl-1, also known as myeloid cell leukemia-1, is a 37-kd protein with significant homology with Bcl-2. Mcl-1 was isolated from a human myeloid leukemia cell line undergoing differentiation, and in these cells Mcl-1 protein was only in-duced transiently [15]. Mcl-1 interferes with the activation of the mitochondrial pathway of apop-tosis and contributes to the resistance of neo-plastic cells towards apoptosis. This is demon-strated by the fact that Mcl-1 is capable of abro-gating c-Myc induced apoptosis in Chinese ham-ster ovarian cells [28]. It has been postulated that in the absence of Bcl-2, Mcl-1 may interact with Bax to induce its anti-apoptotic effect [29].
Several investigators have studied the expres-sion of Mcl-1 protein in human tissues. In one study, Mcl-1 protein was found in well differenti-ated complex epithelia, such as epidermis, in-testine, colon, prostate, nasopharynx, and upper airway, usually localized to the upper portion of the cells [13]. This localization is in antithesis with that of Bcl-2 protein, usually found in the cytoplasm of cells bordering the basement membrane and in less differentiated cells. This may reflect different functions of Bcl-2 and Mcl-1 in the in vivo regulation of apoptosis. While Bcl-2 has been shown to inhibit PCD to support the long term survival of epithelia related stem cells; ultimately regulating their self renewal [30 -32]. Other studies have reported Mcl-1 protein to be localized in either the mitochondrial mem-brane [33], or in the nucleus [15, 34].
Further-more, Liu H. et al. observed Mcl-1 predominant localization in mitochondria if the cells were tested within 24 hours of seeding, and in the nucleus and/or cytoplasm if tested after long culture time [35].
In our study, using the monoclonal antibody MCL1 (RC13), the Mcl-1 protein stain was con-sistently nuclear, as expected according to Gene Tex proprietarian instructions. We found an overall decrease in Mcl-1 expression during the progression from NR to AD and/or CRC. However, this was due to the presence of a sub-set of NR with high Mcl-1 scores. When consid-ering the CRC cohort alone most of the tumors had moderate or strong Mcl-1 expression and correlated well with pAKT expression. Our data are in contrast with the finding of Krajewska et al. reporting the increase in Mcl-1 during the transition from N to AD, but the downregulation of Mcl-1 protein in the invasive tumors. The au-thors explained this finding as a selection bias as they studied tumors that were mostly poorly differentiated.
In this study we found a statistically significant correlation of Mcl-1 with tumor grade and stage. However, these findings must be confirmed on larger patient cohorts. In fact, the tumors we studied included a preponderant percentage of moderately differentiated CRCs, and only 9 can-cers were metastatic.
We also report the increased expression of pAKT during the progression from N to AD to CA. This is an expected finding following our previ-ous observation that in colon cancer the expres-Table 2. Immunostains results and statistics
pAKT Mean(sd) Median Minimum Maximum
Normal (N) 2 .0 (1.4) 2.5 0 4
Adenoma (AD) 3.0 (1.7) 3 0 6
Carcinoma (CA) 5.6 (2.4) 6 0 9
*Wilcoxon rank sum test: CA vs. N, p-value <0.0001; AD vs. CA, p-value = 0.006; AD vs. N, p-value = 0.61
Mcl-1 Mean(sd) Median Minimum Maximum
Normal (N) 4.1 (1.7) 3 2 6
Adenoma (AD) 3.2 (1.2) 3 2 6
Carcinoma (CA) 3.3 (2.6) 3 0 9
* Wilcoxon rank sum test: CA vs. N, p-value = 0.61; AD vs. CA, p-value = 0.76; AD vs. N, p-value = 0.76 Other Statistics:
Presence of
[image:5.612.81.535.98.285.2](IGF1-R) increases with the progression of the disease and that the tumorigenic and anti-apoptotic functions of this receptor are modu-lated via activation of AKT [17, 36].
Itoh et al. have described pAKT in the nucleus and cytoplasm of epithelial cells of normal colo-rectal mucosa. pAKT was localized especially on the surface as opposed to the deep layer of the mucosa [24]. In the same study, 46% of the colorectal carcinoma samples, showed high level of pAKT. The authors reported a close as-sociation between pAKT and Ki-67 proliferative activity, and between pAKT and number of apoptotic bodies [24]. Others have shown pAKT activity to be significantly higher in colorectal cancers than polyps [25]. In our study we ob-served a similar finding with up-regulation of pAKT during the transition from normal colorec-tal mucosa to carcinoma, supporting a role for pAKT in colon carcinogenesis.
When considering the CRC cohort alone, we observed a direct correlation between Mcl-1 and pAKT, reflecting the common anti-apoptotic function of these proteins. These proteins likely favor the progression of colorectal carcinomas by promoting cell growth and by rescuing tumor cells from apoptosis. We also observed the pro-gressive AKT activation during the transition from NR to AD to CRC. These changes were sta-tistically significant. However, pAKT did not cor-relate with tumor grade and/or stage in this size limited study.
Conclusion
This study describes, for the first time, the direct correlation between Mcl-1 protein expression and AKT activation in colorectal cancer. In this study we found that Mcl-1 expression is signifi-cantly correlated with tumor grade and stage, and that AKT activation level increases during the progression from NR to AD to CRC.
Acknowledgments
We thank the Histology Section of the Tissue Core at the Moffitt Cancer Center and Research Institute for the support in performing the im-muno-histochemical stains. We also thank An-drea Dattilo for her help with preparation of the manuscript. The authors have no conflict of in-terests.
Please address correspondence to: Domenico Coppo-la, MD, Anatomic Pathology Division, Moffitt Cancer
nolia Drive, Tampa, Florida 33612-9497, USA. Tel: (813)745-3275; Fax: (813)745-1708; E-mail: Dome-nico.Coppola@moffitt.org
References
[1] Jemal A, Siegel R, Ward E, Murray T, Xu J, and Thun MJ. Cancer Statistics, 2007. CA Cancer J Clin 2007; 57:43-66.
[2] Nishisho I, Nakamura Y, Miyoshi Y, Miki Y, Ando H, Horii A, Koyama K, Utsunomiya J, Baba S, Hedge P. Mutations of chromosome 5q21 genes in FAP and colorectal cancer patients. Science 1991;253(5020):665-9.
[3] Astler VB, Coller FA. The prognostic significance of direct extension of carcinoma of the colon and rectum. Ann Surg1954; 139:846-851. [4] Winawer SJ, Fletcher RH, Miller L, Godlee F,
Stolar MH, Mulrow CD, Woolf SH, Glick SN, Ganiats TG, Bond JH, Rosen L, Zapka JG, Olsen SJ, Giardiello FM, Sisk JE, Van Antwerp R, Brown-Davis C, Marciniak DA, Mayer RJ. Colo-rectal cancer screening: clinical guidelines and rationale. Gastroenterology 1997;112:594-642.
[5] Nakamura Y, Nishisho I, Kinzler KW, Vogelstein B, Miyoshi Y, Miki Y, Ando H, Horii A, Nagase H. Mutations of the adenomatous polyposis coli gene in familial polyposis coli patients and spo-radic colorectal tumors. Princess Takamatsu Symp 1991; 22:285-92.
[6] Cho KR, Oliner JD, Simons JW, Hedrick L, Fearon ER, Preisinger AC, Hedge P, Silverman GA, Vogelstein B. The DCC gene: structural analysis and mutations in colorectal carcino-mas. Genomics 1994; 19:525-31.
[7] Thiagalingam S, Lengauer C, Leach FS, Schutte M, Hahn SA, Overhauser J, Willson JK, Marko-witz S, Hamilton SR, Kern SE, Kinzler KW, Vo-gelstein B. Evaluation of candidate tumor sup-pressor genes on chromosome 18 in colorectal cancers. Nat Genet 1996;13:343-6.
[8] Polyak K, Xia Y, Zweier JL, Kinzler KW, Vogel-stein B. A model for p53-induced apoptosis. Nature 1997; 389(6648):300-5.
[9] Irby RB, Mao W, Coppola D, Kang J, Loubeau JM, Trudeau W, Karl R, Fujita DJ, Jove R, Yeat-man TJ. Activating SRC mutation in a subset of advanced human colon cancers. Nat Genet 1999;21(2):187-90.
[10] Kelsen DP, Daly JM, Kern SE, Levin B, Tepper JE, Van Cutsem E. Principles and Practice Gas-trointestinal Oncology 2nd ed. Philadelphia: Lippincott, Williams and Wilkins, 2008.
[11] Backus HHJ, Van Groeningen CJ, Vos W, Dukers DF, Bloemena E, Wouters D, Pinedo HM, Peters GJ. Differential expression of cell cycle and apoptosis related proteins in colorectal mu-cosa, primary colon tumours, and liver metasta-ses. J Clin Pathol 2002; 55;206-211.
adeno-carcinomas. Cancer Res 1996; 56: 2422-2427.
[13] Krajewski S, Bodrug S, Krajewska M, Shabaik A, Gascoyne R, Berean K, Reed JC. Immunohis-tochemical analysis of Mcl-1 protein in human tissues. Am J Pathol 1995;146:1309-1319. [14] Ming L, Wang P, Bank A, Yu J, Zhang L. Puma
dissociates Bax and Bcl-XL to induce apoptosis in colon cancer cells. JBC 2006; 281(23): 16034-16042.
[15] Kozopas KM, Yang T, Buchan HL, Zhou P, Craig RW. MCL1, a gene expressed in program mye-loid cell differentiation, has sequence similarity to BCL2. Pro. Natl. Acad. Sci. USA 1993; 90:3516-3520.
[16] Schulze-Bergkamen H, Ehrenberg R, Hickmann L, Vick B, Urbanik T, Schimanski CC, Berger MR, Schad A, Weber A, Heeger S, Galle PR, Moehler M. Bcl-XL and Myeloid cell leukaemia-1 contrib-ute to apoptosis resistance of colorectal cancer cells. World J Gastroenterol 2008;14: 3829-3840.
[17] Sekharam M, Zhao H, Sun M, Fang Q, Zhang Q, Yuan Z, Dan HC, Boulware D, Cheng JQ, Coppola D. Insulin-like growth factor 1 receptor enhances invasion and induces resistance to apoptosis of colon cancer cells through the Akt/Bcl-x(L) pathway. Cancer Res 2003 ;63:7708-16.
[18] Sasaki T, Irie-Sasaki J, Horie Y, Bachmaier K, Fata JE, Li M, Suzuki A, Bouchard D, Ho A, Red-ston M, Gallinger S, Khokha R, Mak TW, Haw-kins PT, Stephens L, Scherer SW, Tsao M, Pen-ninger JM. Colorectal carcinoma in mice lack-ing catalytic subunit of PI(3)K gamma. Nature 2000; 406 (6798):897-902.
[19] Cantley LC, Neel BG. New insights into tumor suppression: PTEN suppresses tumor formation by restraining the phophoinositide 3-kinase/ AKT pathway. Proc Natl Acad Sci USA 1999; 96 (8):4240-4245.
[20] Khwaja A. Akt is more than just a Bad kinase. Nature 1999; 401(6748):33-34.
[21] El-Deiry WS. Akt takes centre stage in cell-cycle deregulation. Nat Cell Biol 2001; 3(3):E71-73. [22] Sheng H, Shao J, Washington MK, DuBois RN.
Prostaglandin E2 increases growth and motility of colorectal carcinoma cells. J Biol Chem 2001; 276(21): 18075-18081.
[23] Kureishi Y, Luo Z, Shiojima I, Bialik A, Fulton D, Lefer DJ, Sessa WC, Walsh K. The HMG-CoA reductase inhibitor simvastatin activates the protein kinase Akt and promotes angiogenesis in normocholesterolemic animals. Nat Med 2000; 6(9):1004-1010.
[24] Itoh N, Semba S, Ito M, Takeda H, Kawata S, Yamakawa M. Phosphorylation of Akt/PKB is required for suppression of cancer cell apop-tosis and tumor progression in human colorec-tal carcinoma. Cancer 2002; 94(12):127-134. [25] Colakoglu T, Yildirim S, Kayaselcuk F, Nursal
TZ, Ezer A, Noyan T, Karakayali H, Haberal M. Clinicopathological significance of PTEN loss and the phosphoinositide 3-kinase/Akt
path-way in sporadic colorectal neoplasms: is PTEN loss predictor of local recurrence? Am J Surg 2008; 195:719-725.
[26] Cotran RS, Kumar V, Collins T. Cellular injury and cell death. In: Robbins Pathologic basis of disease.7th ed. Philadelphia: Elsevier Saun-ders,2004.
[27] Maddika S, Ande SR, Panigrahi S, Weglarczyk K, Zuse A, Eshraghi M, Manda KD, Wiechec E, Los M. Cell survival, cell death and cell cycle pathways are interconnected: implications for cancer therapy. Drug Resistance Updates 2007;10:13-29.
[28] Reynolds JE, Yang T, Qian L, Jenkinson JD, Zhou P, Eastman A, Craig RW. Mcl-1, a member of the Bcl-2 family, delays apoptosis induced by c-Myc overexpression in Chinese hamster ovary cells. Cancer Res 1994;54:6348 6352. [29] Krajewski S, Krajewska M, Shabaik A,
Miya-shita T, Wang HG, Reed JC. Immunohistochemi-cal determination of in vivo distribution of Bax, a dominant inhibitor of Bcl-2. Am J Pathol 1994;145: 1323-1333.
[30] Adams JM, Kelly PN, Dakic A, Carotta S, Nutt SL, Strasser A. Role of "cancer stem cells" and cell survival in tumor development and mainte-nance.Cold Spring Harb Symp Quant Biol 2008;73:451-9.
[31] LeBrun DP, Warnke RA, Cleary ML. Expression of bcl-2 in fetal tissues suggests a role in morphogenesis. Am J Pathol 1993; 142:743-753.
[32] Kamada S, Shimono A, Shinto Y, Tsujimura T, Takahashi T, Noda T, Kitamura Y, Kondoh H, Tsujimoto Y. Bcl-2 deficiency in mice leads to pleiotropic abnormalities: accelerated lymphoid cell death in thymus and spleen, polycystic kidney, distorted intestine and hair hypopig-mentation. Cancer Res 1994; 55:354-359. [33] Yang T, Kozopas KM, Craig RW. The
intracellu-lar distribution and pattern of expression of Mcl -1 overlap with, but are not identical to those of Bcl-2. J Cell Biol 1995; 128:1173-1184. [34] Zhang D, Li F, Weidner D, Mnjoyan ZH, Fujise K.
Physical and functional interaction between myeloid cell leukemia 1 protein (MCL1) and fortilin: the potential role of MCL1 as a fortilin chaperone. J Biol Chem 2002; 7:37430-37438.
[35] Liu H, Peng HW, Cheng, YS, Yuan HS, Yang-Yen HF. Stabilization and Enhancement of the An-tiapoptotic Activity of Mcl-1 by TCTP. Mol Cell Biol 2005; 25(8):3117-3126. 33