JOURNAL OFCLINICALMICROBIOLOGY, Sept. 2008, p. 2906–2911 Vol. 46, No. 9 0095-1137/08/$08.00⫹0 doi:10.1128/JCM.00421-08
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Molecular Characterization of Human-Colonizing
Streptococcus agalactiae
Strains Isolated from Throat, Skin, Anal Margin, and Genital Body Sites
䌤
Nathalie van der Mee-Marquet,
1,2* Laure Fourny,
2Laurence Arnault,
2Anne-Sophie Domelier,
1,2Mazen Salloum,
2Marie-Fre
´de
´rique Lartigue,
1,2and Roland Quentin
1,2Universite´ Franc¸ois-Rabelais, IFR 136, Faculte´ de Me´decine, EA 3854, Bacte´ries et Risque Materno-Fœtal, Tours, France,1and
Laboratoire de Bacte´riologie et Hygie`ne, Hoˆpital trousseau, CHU, Tours, France2
Received 3 March 2008/Returned for modification 15 April 2008/Accepted 7 July 2008
Streptococcus agalactiae carriage was evaluated by sampling four body sites in a group of 249 healthy
individuals including both sexes and a wide range of ages; the aims were to study the population structure of colonizing strains by multilocus sequence typing (MLST) and to evaluate their diversity by serotyping, SmaI macrorestriction analysis, and PCR screening for genetic markers of highly virulent clones for neonates. The prevalences of carriage were 27% in women and 32% in men. The major positive body site was the genital tract (23% in women and 21% in men); skin, throats, and anal margins were also positive in 2%, 4%, and 14%, respectively. These human-colonizing strains belonged mostly to serotypes III (24%), Ia (21%), V (18%), and Ib (17%). Twenty-three sequence types (STs) were identified. The MLST characteristics of the strains isolated from a single anatomic site—genital (vagina [women] or from a sample of the first urination after arising from a night’s sleep [men]), throat, skin, or anal margin—suggest a body site colonization specificity for particular STs: strains of STs 2, 10, 19, and 196 were isolated only from genital sites; strains of STs 1, 8, and 23 were isolated more frequently from throat florae; and strains recovered only from anal margin samples were more closely related to strains isolated from throats than to those from genital sites. Most strains of STs 1, 8, and 23—STs that are increasingly described as being responsible for adult infections—did not carry any markers of strains virulent for neonates, suggesting that the virulence of these strains is probably associated with other genetic determinants. In addition, the genetic diversities of the strains varied between STs: STs 2, 8, 10, 23, and 196 were the most diverse; STs 1 and 19 were more homogeneous; and ST 17 strains formed three distant groups.
Streptococcus agalactiaeis the pathogen most commonly re-sponsible for maternofetal and neonatal infections, and sero-group III sequence type (ST) 17 strains of this species have been identified as more likely to cause these infections (3, 17, 24–26, 28).
S. agalactiae infections in nonpregnant adults have been reported increasingly since the 1970s (5, 7, 10, 14, 20, 21, 23, 26, 27). The major clinical forms consist of skin, soft tissue, and bone infections; bacteremia; urinary tract infections; pneumo-nia; and peritonitis. More rarely,S. agalactiaeis responsible for septic arthritis, meningitis, and endocarditis. Risk factors have been identified (16) and include an age of over 60 years and diagnoses of diabetes, cancer, decubitus acutus, AIDS, long-course corticotherapy, chronic renal disease, cirrhosis, and neurological vessel disorders. However, cases of invasive S. agalactiaeinfections in immunocompetent nonpregnant adults have recently been reported (7, 20, 21, 23). The physiopathol-ogy of these infections is not well understood and may be linked in part toS. agalactiaestrains with particular properties. Reports on the genetic characteristics of the strains associ-ated with infections in adults showed that they are diverse and
belong mostly to serotypes Ia, III, and V (5, 14) and clonal complexes 1, 9, 17, 19, and 23 (17).
The portal of entry of these infections in adults is not often well documented. A better knowledge of the genetic charac-teristics of theS. agalactiaestrains colonizing various body sites would contribute to a better understanding of the origin of infections in adults. Although the molecular features of S. agalactiaestrains isolated during infectious diseases have been described in many cases, little is known about the molecular characteristics of human-colonizing strains. Therefore, we screened forS. agalactiaecarriage in 249 healthy individuals of both sexes; the individuals were living at home and covered a range of ages, from the start of the teenage period until old age. Genital, skin, throat, and anal margin carriage was sam-pled by self-swabbing. The strains isolated were serotyped. The population structure of the strains was investigated by multilo-cus sequence typing (MLST). The genetic diversity of isolates was evaluated by SmaI macrorestriction analysis and PCR screening for five previously described genetic markers ofS. agalactiaestrains able to invade the central nervous systems (CNS) of neonates. Genetic features of colonizing strains iso-lated from the various anatomic sites were studied.
MATERIALS AND METHODS
Population studied.The objective was to enroll 25 to 30 healthy females and 25 to 30 healthy males in each of the five following age ranges: 11 to 24 years, 25
to 40 years, 41 to 54 years, 55 to 70 years, andⱖ70 years. After approval of the
study by the local ethical committee, 249 French individuals of ages from 11 to
* Corresponding author. Mailing address: Laboratoire de Bacte ´ri-ologie et Hygie`ne, Hoˆpital Trousseau, 37044 Tours cedex, France. Phone: (33) 234 389 430. Fax: (33) 247 478 588. E-mail: n.vandermee @chu-tours.fr.
䌤Published ahead of print on 16 July 2008.
2906
on May 16, 2020 by guest
http://jcm.asm.org/
94 years, including 133 women and 116 men, were enrolled. All participating individuals were informed about the study and signed a personal consent form.
S. agalactiaestrain isolation.Self-swabbing was conducted in the morning before washing. Volunteers were instructed to swab or sample the following body sites or product: (i) for both sexes (swabbing), the throat, the interdigital spaces on the feet, and the anal margin; (ii) for females (swabbing), the vulva for teenage (up to 16-year-old) girls and the vagina for adults; and (iii) for men (sampling), the first urination after arising from a night’s sleep (herein referred
to as first urine), which was used to identifyS. agalactiaecolonization of the
meatus urinarius, the preputial gland, and the end of the urethra. Swabs were transmitted to the laboratory within 12 h. Swabs and urine samples (1 ml) were
placed in Todd-Hewitt broth supplemented with gentamicin (8g/ml) and
colis-tin (5g/ml) and incubated for 24 h at 37°C. The selection broths were then
subcultured on streptoB agar medium (Laboratoire AES, Combourg, France) and incubated for 48 h under anaerobic conditions at 37°C. Each morphotype of orange-colored colonies was plated on 5% sheep blood agar (Laboratoire
bio-Me´rieux, Marcy l’Etoile, France) and incubated for 24 h under aerobic
condi-tions with CO2at 37°C. Vitek2 g-positive identification cards (Laboratoire
bio-Me´rieux, Marcy l’Etoile, France) were used for strain identification.
Serotyping.The strains were serotyped using PCR according to a previously published procedure (18).
SmaI DNA macrorestriction and PFGE.DNA was extracted from the isolates, digested with SmaI, and then subjected to pulsed-field gel electrophoresis (PFGE) as previously described (25). The PFGE patterns were analyzed with the Taxotron package (Taxolab; Institut Pasteur, Paris, France).
MLST.The strains were analyzed using MLST according to the procedure described by Jones et al. (17). A tree based on the use of an unweighted-pair group method using average linkages was drawn from allelic profile data by use
of Phylodendron and the entire group BStreptococcus(GBS) MLST database
(http://pubmlst.org/sagalactiae/).
Detection of genetic markers of virulentS. agalactiaestrains.Some genetic markers are significantly more frequent in strains able to invade the CNS of neonates than in other strains, namely, the GBSi1 group II intron downstream
from the C5a-peptidasescpBgene (3), IS1548in thehylBgene (25), and three
prophagic DNA fragments (28). PCR was used to test for the presence of these markers as previously described (3, 25, 28).
Statistical data.Chi-square tests and Fisher’s exact test (two tailed) were used
to test associations, and aPvalue of 0.050 was considered significant.
RESULTS
Population studied.A total of 249 individuals were enrolled in the study, including 133 females and 116 males. Consistent with the study design, we recruited 25 to 30 individuals of each
sex in each age class except one (Table 1). For those ofⱖ70 years of age, fewer men than expected were recruited (n⫽10). The 249 enrolled individuals included 137 individuals unre-lated to other participants and 112 individuals belonging to 47 family groups as follows: 37 partner pairs (29 with no child, 2 with one child, 4 with two children, and 2 with three children), and 3 monoparental groups (1 group with a parent and a child and 2 groups with a parent and two children), and 7 pairs of brothers and/or sisters. Therefore, these family groups in-cluded 37 partner pairs, 37 parent-child pairs, and 19 sibling groups.
Prevalence of carriage.104S. agalactiaestrains were isolated from 73 of the 249 individuals (29%). The strains were isolated from 36 of the 133 females (27%) and 37 of the 116 males (32%). Two morphotypes were observed for an anal margin sample from one male (M113).
(i) Prevalence of carriage according to sex and age. The prevalence of carriage varied according to sex and age (Table 1). For women, the prevalence was between 20% for those agedⱖ70 years and 41% for those of ages between 41 and 54 years. The prevalence for those of ages between 41 and 54 years (41%) was significantly higher than those for other age groups (20 to 27%) (P⫽0.048). For men, the prevalence was more variable (Table 1). Carriage was rare (4%) for those of ages between 11 and 24 years and increased with age to 48% for those of ages between 25 and 40 years and 54% for those of ages between 41 and 54 years. The prevalence was signifi-cantly higher for those of ages between 25 and 40 years and between 41 and 54 years than for those of ages between 11 and 24 years, between 55 and 70 years, and of ⱖ70 years (P ⬍
0.001). The prevalence was significantly higher for women than for men only for those of ages between 11 and 24 years (14 versus 4%) (P⫽0.029).
(ii) Prevalence of carriage according to the site of isolation.
[image:2.585.42.542.81.260.2]The prevalence of carriage varied according to the site of isolation (Table 1). The major positive body site was the gen-ital tract (23% from vagina in women, 21% from first urine in
TABLE 1. Prevalence ofS. agalactiaecarriage by sex, age, and body site
Sex and body site sample type
No. of individuals (% prevalence) withS. agalactiaecarriage in indicated sexaand age groups
11–24 yr (n⫽57;
29 F, 28 M)
25–40 yr (n⫽51;
24 F, 27 M)
41–54 yr (n⫽60;
32 F, 28 M)
55–70 yr (n⫽46;
23 F, 23 M)
ⱖ70 yr (n⫽35;
25 F, 10 M)
All ages (n⫽249;
133 F, 116 M)
Both sexes (n⫽249) 8 (14) 19 (37) 28 (47) 11 (24) 7 (20) 73 (29)
Throat 1 (2) 3 (6) 3 (5) 1 (2) 1 (3) 9 (4)
Genitald 7 (12) 13 (25) 21 (35) 8 (17) 6 (17) 55 (22)
Anal margin 3 (5) 5 (10) 14 (23) 7 (15) 5 (14) 34 (14)
Skinb 1 (2) 1 (2) 1 (2) 2 (6) 5 (2)
Females (n⫽133) 7 (24) 6 (25) 13 (41) 5 (22) 5 (20) 36 (27)
Throat 1 (3) 1 (4) 1 (3) 1 (4) 4 (3)
Vaginal 6 (21) 5 (21) 10 (31) 4 (17) 5 (20) 30 (23)
Anal margin 3 (10) 1 (4) 6 (19) 2 (9) 4 (16) 16 (12)
Skinb 1 (4) 1 (4) 2 (1)
Males (n⫽116) 1 (4) 13 (48) 15 (54) 6 (26) 2 (20) 37 (32)
Throat 2 (7) 2 (7) 1 (4) 5 (4)
First urinec 1 (4) 8 (30) 11 (39) 4 (17) 1 (10) 25 (21)
Anal margin 4 (15) 8 (29) 5 (22) 1 (10) 18 (15)
Skinb 1 (4) 1 (4) 1 (10) 3 (3)
aF, female; M, male.
bSkin carriage was studied by swabbing the interdigital space of the foot.
cFor males, first-urine samples were used to studyS. agalactiaecolonization in the meatus urinarius, the preputial gland, and the end of the urethra.
dS. agalactiaegenital colonization was scored as the sum of vaginal and first-urine sample colonizations.
on May 16, 2020 by guest
http://jcm.asm.org/
men), followed by the anal margin (14%), the throat (4%), and skin (2%).
The prevalence of carriage in various body sites did not differ significantly between men and women of ages ofⱖ25 years (Table 1). By contrast, for those of ages between 11 and 24 years, vaginal carriage was frequent in women (21%), whereas genital carriage was rare (4%) in men (P⬍ 0.001). The frequency of carriage at nongenital sites did not differ significantly according to sex or age.
Carriage in family groups. (i) Partner pairs (37 cases).
Carriage was found for 18 partner pairs (49%) in at least one partner. Carriage was observed for both partners in 11 of the 18 cases (61%), for only the female in 2 cases (11%) and for only the male in 5 cases (28%).
(ii) Relations between parent and child (37 cases).Carriage was identified for four parents and none of their children were carriers. One child (female, 14 years of age) carriedS. agalac-tiae, but her parents wereS. agalactiaenegative.
(iii) Relations between siblings (19 cases). Carriage was identified for four females of 12, 14, 15, and 18 years of age, all having one or two siblings. No carriage was found for the siblings of these carriers.
Carriage and body site of colonization.Forty-seven (21 fe-males, 26 males) of the 73 carriers were positive at a single body site (64%). Carriage was rarely present in more than two body sites (5%).
(i) Female carrier group.Screening of the vagina/vulva, the anal margin, and the throat for 133 women detectedS. agalac-tiae in 35 individuals (26%), accounting for 97% of the 36 carriers. For cases in which there was a single positive site of carriage, it was the vagina/vulva in 15 cases, the anal margin in 2 cases, the throat in 3 cases, and the skin in 1 case. For 14 cases, two body sites were positive (vagina and anal margin). One female was positive at her vagina, throat, and skin.
(ii) Male carrier group.Samples of the first urine, the anal margin, or the throat wereS. agalactiaepositive for 35 of the 116 men (30%), accounting for 95% of the 37 carriers. For cases of a single positive site, carriage was in the first urine in 14 cases, at the anal margin in 7 cases, in the throat in 3 cases, and on the skin in 2 cases. For eight cases, two body samples were positive (first urine and anal margin). For the remaining males, the carriage was positive for three body samples (first urine, anal margin, and throat in two cases and first urine, anal margin, and skin in one case).
Serotypes of human-colonizing strains. Ninety-four of the 104 human-colonizing strains (90%) were typeable. The major serotypes were serotype III (25 strains; 24%), Ia (22 strains; 21%), V (19 strains; 18%), and Ib (18 strains; 17%). Strains belonging to serotypes II (n⫽4; 4%) and IV (n ⫽6; 6%) were rare.
(i) Partner pairs with carriage in both partners (11 cases).
For 10 (91%) of the partner pairs with carriage in both part-ners, the serotypes of strains isolated from the partners were similar. The strains isolated from the partners of one pair were of different serotypes (serotype Ib from male M115 and sero-type V from female F87).
(ii) Individuals with multisite carriage (26 cases). For 25 (96%) of the individuals with multisite carriage, all strains isolated from different body sites were of the same serotype. For the remaining individual, M219, a nontypeable strain, was
isolated from the first urine and a serotype IV strain was iso-lated from the anal margin sample. The two morphotypes isolated from the anal margin swab of M113 both belonged to serotype III.
PFGE characteristics of human-colonizing strains. Fifty-one pulsotypes were obtained with the 104 strains.
(i) Carriage by independent carriers (51 cases). Forty-six pulsotypes were obtained with the 51 strains.
(ii) Partner pairs with carriage in both partners (11 cases).
Strains isolated from both partners in nine partner pairs (82%) were of similar pulsotypes in each partner pair, with each partner pair having a different pulsotype. For one partner pair (M82, F94), the strains from the partners were both of serotype V and of different pulsotypes but were closely related (20% dissimilarity). For the 11th partner pair (M115, F87), the two strains were of different serotypes and in distantly related pulso-types (90% dissimilarity).
(iii) Individuals with multisite carriage (26 cases).For 24 individuals (92%) in the multisite-carriage category, the strains isolated from all positive sites in the body gave similar pulso-types. For individual M219, who had strains of different sero-types, and for individual M165, who had strains of serotype Ib, the strains from the first urine were of a pulsotype distantly related to those of the strains isolated from other positive sites (70% dissimilarity).
The two morphotypes isolated from the anal margin swab from individual M113 showed similar pulsotypes.
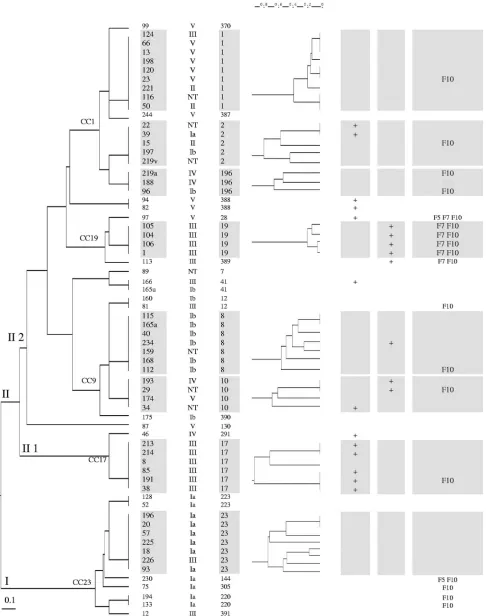
Genetic diversity of the human-colonizing strains.To deter-mine the population structure of the strains and their genetic diversity, we used MLST. We first excluded redundant strains: strains were considered redundant when the isolates from both partners in partner pairs and/or from several body sites of an individual exhibited similar serotypes and pulsotypes. Accord-ing to these criteria, there were 66 independent strains among the 104 isolates. Twenty-three STs were identified by MLST for the 66 strains (Fig. 1), and 45 strains (68%) clustered into eight major STs. Two major phylogenetic groups, groups I and II, were identified. Group I was homogeneous and corre-sponded to clonal complex CC 23. Group II comprised two subdivisions: a homogeneous subgroup (II 1) that corre-sponded to CC 17, and a heterogeneous subgroup (II 2) that included the six remaining major STs, which were STs 1, 2, 196, 19, 8, and 10.
(i) MLST and anatomical origin of strains.For this analysis, we included only the 36 strains isolated from a single anatomic site to ensure that they represented the considered anatomical origin (Table 2). The 12 strains belonging to STs 2, 196, 19, and 10 were all isolated only from genital sites (vagina and first-urine samples). STs 1, 8, and 23 were better represented among strains isolated only from throat (4/6; 67%) than among strains isolated from genital sites (vagina and first-urine sam-ples [2/21; 9%]); similarly, these STs were more common among strains recovered only from the anal margin samples (4/7; 57%) than among strains isolated only from genital sites (vagina and first-urine samples). The seven strains isolated only from the anal margin samples and the six strains isolated only from throat samples were similarly distributed among STs.
(ii) MLST and serotype (Fig. 1).ST 1 was significantly as-sociated with serotype V (P⫽0.001), ST 8 with Ib (P⬍0.001),
2908 VAN DER MEE-MARQUET ET AL. J. CLIN. MICROBIOL.
on May 16, 2020 by guest
http://jcm.asm.org/
FIG. 1. Tree based on MLST and an unweighted-pair group method using average linkages (UGPMA) of the 66 unrelatedS. agalactiaestrains and PFGE diversity according to ST. Footnotes:1, strains were given a code corresponding to the carrier code;2, ST designation;3, PCR result
for GBSi1 group II intron downstream from the C5a-peptidasescpBgene;4, PCR result for IS1548in thehylBgene;5, PCR result for the three
prophage DNA fragments. NT, nontypeable.
2909
on May 16, 2020 by guest
and ST 23 with Ia (P⬍0.001). ST 17 and ST 19 were signifi-cantly associated with serotype III (P⬍0.001).
(iii) MLST and virulence markers of strains able to invade the CNS of neonates (Fig. 1).ST 17 was significantly associated with positivity for the GBSi1 group II intron downstream from thescpBgene (P⬍0.001), and ST 19 with positivity for IS1548
inhylB(P⬍0.001) and for prophage DNA fragments F7 and F10 (Pvalues of⬍0.001 and⬍0.004, respectively). In CC 9, containing ST 8 and ST 10, three of the four strains belonging to ST 10 had a diverse set of virulence markers and, by con-trast, five of the seven strains belonging to ST 8 contained none of the virulence markers studied. Most strains of STs 1, 8, and 23 did not carry any of the genetic markers studied.
(iv) MLST and PFGE (Fig. 1).PFGE diversity within each of the eight major STs was investigated. STs 2, 196, 8, 10, and 23 exhibited substantial diversity of PFGE patterns, with 23 pulsotypes among the 26 strains. STs 1 and 19 exhibited the lowest diversity, with maximal percentages of dissimilarity of 40% for ST 1 and 20% for ST 19. ST 17 exhibited a particular pattern, with six strains clustering in three distantly related pulsotypes (90% dissimilarity).
DISCUSSION
TheS. agalactiaecolonization rates reported in diverse pop-ulations vary from 2 to 30% (1, 2, 4, 6, 8, 9, 11, 12, 15, 19, 22). Using a procedure recommended for the detection ofS. aga-lactiaefrom lower vaginal and rectal samples (4, 6) and self-swabbing, we obtained a carriage rate (29%) consistent with, although slightly higher than, values reported by others; this validates the procedure used and demonstrates the efficiency of self-swabbing forS. agalactiaedetection.
Data concerning the course ofS. agalactiaecolonization are divergent. In view of the high rates of anorectal colonization, it has been suggested that the primary site of infection is the lower gastrointestinal tract and consequently that genital car-riage merely reflects contamination from the anorectum (1, 4, 9). However,S. agalactiaecolonization of genital sites may be equally high, and a high prevalence of carriage among sexually active populations has been described (12, 19); these observa-tions led to the idea that vaginal colonization and sexual con-tact are major factors for the transmission ofS. agalactiae(19). The apparent discrepancies between the various findings may
be consequences of the specificities of the populations studied. In particular, girl college students (2, 4, 12, 19), pregnant women (1), and elderly women in menopause may present different characteristics depending on their gastrointestinal and genitourinary tract conditions and their sexual activity. Our study, assessing S. agalactiae colonization in a larger healthy population, which was representative of both sexes and a wide range of ages, brings additional data. First, in the young-est group (⬍18 years of age), colonization was rare in males although notable in females, with vaginal colonization twice as frequent as anal colonization, and anal colonization was in all cases associated with vaginal colonization. Second, various ob-servations are informative for the three median-age classes (25 to 70 years of age): (i) the vaginal colonization rate was higher than the anorectal rate; (ii) genital colonization was highest (54%) in 41- to 54-year-old men; and (iii) the prevalence of carriage among partner pairs was high, and strains isolated from most partners were closely related. Third, colonization was less prevalent among men in the oldest age class (⬎70 years of age) than among the younger-age classes. Fourth, in accordance with previously published data (12, 19), anal car-riage was not observed in family members who have no sexual contact (parent/child, child/child), despite them presumably sharing food and food-related utensils. These observations are consistent with sexual contact playing an important role in the spread ofS. agalactiae.Nevertheless, one of the limitations of our study is that we did not evaluate either the sexual activity of the carriers or rectal carriage. Therefore, other explanations of the variations in carriage rate between sexes and according to age are possible, such as different ecological conditions in genital tracts for bacterial colonization depending, for exam-ple, on the hormonal status of the individuals.
S. agalactiaeis responsible for pneumonia and adult invasive infections (5, 7, 10, 14, 20, 21, 23, 26, 27) including bacteremia, endocarditis (sometimes associated with vertebral infections), endophtalmitis, and meningitis in patients for whom the gen-ital tract is unlikely to be the portal of entry (7, 13, 20). By comparing MLST characteristics of strains from a single ana-tomic site, we found that strains isolated from the throat dif-fered from genital strains and belonged mostly to STs 1, 8, and 23, three STs previously described as being involved in infec-tions of nonpregnant adults (17). These observainfec-tions suggest that the throat could be a portal of entry for adult invasive infections; nevertheless, given the small number of isolates we studied, this notion requires confirmation by further studies including larger numbers of throat carriage strains.
We report the molecular characteristics and genetic diversity of human-colonizing strains. Most were equally distributed among the four major serotypes involved in invasive cases of disease (Ia, Ib, III, and V) (5, 17, 20, 26). The major STs that we identified for colonizing strains have also been identified as major STs for strains isolated during infectious diseases (17). Indeed, 68% of colonizing strains belonged to STs 1, 2, 8, 17, 19, 23, and 196, STs that have been associated with neonatal and/or adult infections (17). Using MLST data and genetic characteristics as assessed by PFGE and PCR for several mark-ers of horizontal transfer (introns, insertion elements, and prophagic elements), we found substantial differences in the diversities of strains according to their STs as follows and as shown in Fig. 1: (i) in STs 1 and 19, the strains showed a low
TABLE 2. Site of carriage and phylogenetic position defined by MLST (STs) of the colonizingS. agalactiaestrains isolated from a
single anatomic site: genital tract (vagina or first urine), throat, skin, or anal margin
Body site of
isolationa
No. of strains of indicated ST
1 2 8 10 17 19 23 196 Others Total
Throat 2 1 1 2 6
Anal margin 2 1 1 3 7
Skin 1 1 2
Genital tractb 1 3 3 1 4 1 2 6 21
Total 6 3 2 3 2 4 3 2 11 36
aCarriage site is reported in this table only when the carriage was found only
at a single body site.
bVagina for women; first urine for men.
2910 VAN DER MEE-MARQUET ET AL. J. CLIN. MICROBIOL.
on May 16, 2020 by guest
http://jcm.asm.org/
level of genetic diversity, which may indicate the recent emer-gence of clones or highly selective environmental conditions; (ii) in STs 2, 8, 10, 23, and 196, the strains showed a high genetic diversity, and these strains therefore may have been subject to various ecological conditions and consequently dis-play genome diversification; and (iii) in ST 17, most of the colonizing strains were positive for the GBSi1 group II intron downstream from the C5a-peptidase genescpBbut could be classified into one of three distant PFGE groups that may result from different genetic events or ecological pressure. By contrast to STs 2, 10, 17, 19, and 196, most of the strains of STs 1, 8, and 23 did not carry any markers of strains responsible for neonatal meningitis (Fig. 1). This suggests that strains of these STs may have mechanisms different from those involved in the invasion of the CNS of neonates.
In conclusion, our study suggests a possible body site colo-nization specificity of the various phylogenetic groups (STs) that compose theS. agalactiaespecies and shows differences of genetic diversification between STs. Further studies should be conducted to elucidate how the body site environments may have selected bacteria having a particular fitness.
ACKNOWLEDGMENT
This work was supported by the Centre Hospitalier Universitaire, Tours, France.
REFERENCES
1.Badri, M. S., S. Zawaneh, A. C. Cruz, G. Mantilla, H. Baer, W. N. Spellacy, and E. M. Ayoub.1977. Rectal colonization with group B Streptococcus:
relation to vaginal colonization of pregnant women. J. Infect. Dis.135:308–
312.
2.Baker, C. J., D. K. Goroff, S. Alpert, V. A. Crockaett, S. H. Zinner, J. R. Evrard, B. Rosner, and W. M. McCormack.1977. Vaginal colonization with
group B Streptococcus: a study in college women. J. Infect. Dis.135:392–397.
3.Bidet, P., N. Brahimi, C. Chalas, Y. Aujard, and E. Bingen.2003. Molecular characterization of serotype III group B-Streptococcus isolates causing
neo-natal meningitis. J. Infect. Dis.188:1132–1137.
4.Bliss, S. J., S. D. Manning, P. Tallman, C. J. Baker, M. D. Pearlman, C. F. Marrs, and B. Foxman.2002. Group BStreptococcuscolonization in male and nonpregnant female university students: a cross-sectional prevalence
study. Clin. Infect. Dis.34:184–190.
5.Blumberg, H. M., D. S. Stephens, M. Modansky, M. Erwin, J. Elliot, R. R. Facklam, A. Schuchat, W. Baughman, and M. M. Farley.1996. Invasive group B streptococcal disease: the emergence of serotype V. J. Infect. Dis.
173:365–373.
6.Brimil, N., E. Barthell, U. Heindrichs, M. Kuhn, R. Lu¨tticken, and B. Spellerberg.2005. Epidemiology ofStreptococcus agalactiaecolonization in
Germany. Int. J. Med. Microbiol.296:39–44.
7.Cordoba-Lopez, A., M. I. Bueno Alvarez-Arenas, J. Monterrubio-Villa, and G. Corcho-Sanchez.2002.Streptococcus agalactiaepleural empyema in a
healthy adult. Enferm. Infecc. Microbiol. Clin.20:478–479.
8.Cumming, C. G., and P. W. Ross.1981. Carriage of group B streptococci in
the upper respiratory tract. J. Clin. Pathol.34:813.
9.Easmon, C. S. F., A. Tanna, P. Munday, and S. Dawson.1981. Group B
streptococci, gastrointestinal organisms? J. Clin. Pathol.34:921–923.
10.Farley, M. M., R. C. Harvey, T. Stull, J. D. Smith, A. Schuchat, J. D. Wenger, and D. S. Stephens.1993. A population-based assessment of invasive disease
due to group BStreptococcusin nonpregnant adults. N. Engl. J. Med.328:
1807–1811.
11.Ferrieri, P., and L. L. Blair.1977. Pharyngeal carriage of group B
strepto-cocci: detection by three methods. J. Clin. Microbiol.6:136–139.
12.Foxman, B., B. Gillespie, S. D. Manning, L. J. Howard, P. Tallman, L. Zhang, and C. F. Marrs.2006. Incidence and duration of group B Strepto-coccusby serotype among male and female college students living in a single
dormitory. Am. J. Epidemiol.163:544–551.
13.Fromm, S. E., and J. S. Toohey.1996. Septic arthritis of the hip in an adult
following repeated femoral venipuncture. Orthopedics19:1047–1048.
14.Harrison, L. H., J. A. Elliot, D. M. Dwyer, J. P. Libonati, P. Ferrieri, L. Billmann, A. Schuchat, et al.1998. Serotype distribution of invasive group B streptococcal isolates in Maryland: implications for vaccine formulation.
J. Infect. Dis.177:998–1002.
15.Islam, A., and E. Thomas.1980. Faecal carriage of group B streptococci.
J. Clin. Pathol.33:1006–1008.
16.Jackson, L. A., R. Hilsdon, M. M. Farley, L. H. Harrison, A. L. Reingold, B. D. Plikaytis, J. D. Wenger, and A. Schuchat.1995. Risk factors for group
B streptococcal disease in adults. Ann. Intern. Med.123:415–420.
17.Jones, N., J. F. Bohnsack, S. Takahashi, K. A. Oliver, M. S. Chan, F. Kunst, P. Glaser, C. Rusniok, D. W. Crook, R. M. Harding, N. Bisharat, and B. G. Spratt.2003. Multilocus sequence typing system for group BStreptococcus.
J. Clin. Microbiol.41:2530–2536.
18.Kong, F., S. Gowan, D. Martin, G. James, and G. L. Gilbert.2002. Serotype identification of group B streptococci by PCR and sequencing. J. Clin.
Microbiol.40:216–226.
19.Manning, S. D., P. Tallman, C. J. Baker, B. W. Gillespie, C. F. Marrs, and B. Foxman.2002. Determinants of co-colonization with group B
Streptococ-cusamong heterosexual college partner pairs. Epidemiology13:533–539.
20.Martins, E. R., C. Florindo, F. Martins, I. Aldir, M. J. Borrego, L. Brum, M. Ramirez, and J. Melo-Cristino.2007.Streptococcus agalactiaeserotype Ib as an agent of meningitis in two adult nonpregnant women. J. Clin. Microbiol.
45:3850–3852.
21.Matsuo, K., K. Nakatuka, Y. Yano, W. Fujishima, and K. Kashima.1998. Group B streptococcal metastatic endophtalmitis in an elderly man without
predisposing illness. Jpn. J. Ophtalmol.42:304–307.
22.Maurer, M., M. C. Thirumoorthi, and A. S. Dajani.1979. Group B
strep-tococcal colonization in prepubertal children. Pediatrics64:65–67.
23.Mouly, S., F. Berenbaum, and G. Kaplan. 1999. Group B streptococcal vertebral osteomyelitis with bacteraemia in an adult with no debilitating
condition. Scand. J. Infect. Dis.31:316–317.
24.Musser, J. M., S. J. Mattingly, R. Quentin, A. Goudeau, and R. K. Selander.
1989. Identification of a high-virulence clone of type IIIStreptococcus
aga-lactiae(group BStreptococcus) causing invasive neonatal disease. Proc. Natl.
Acad. Sci. USA86:4731–4735.
25.Rolland, K., C. Marois, V. Siquier, B. Cattier, and R. Quentin.1999. Genetic
features ofStreptococcus agalactiaestrains causing severe neonatal
infec-tions, as revealed by pulsed-field gel electrophoresis andhylBgene analysis.
J. Clin. Microbiol.37:1892–1898.
26.Schuchat, A.1998. Epidemiology of group B streptococcal disease in the
United States: shifting paradigms. Clin. Microbiol. Rev.11:497–513.
27.Tyrrell, G. J., L. D. Senzilet, J. S. Spika, D. A. Kertesz, M. Alagaratnam, and J. A. Lovgren.2000. Invasive disease due to group B streptococcal infection in adults: results from a Canadian, population-based, active laboratory
sur-veillance study-1996. J. Infect. Dis.182:168–173.
28.van der Mee-Marquet, N., A. S. Domelier, L. Mereghetti, P. Lanotte, A. Rosenau, W. van Leeuwen, and R. Quentin.2006. Prophagic DNA fragments inStreptococcus agalactiaestrains and association with neonatal meningitis.
J. Clin. Microbiol.44:1049–1058.