0021-9193/80/01-0111/10$02.00/0
Control of Plasmid
Ri Replication: Functions Involved in
Replication,
Copy
Number
Control,
Incompatibility, and
Switch-off of Replication
S0REN MOLIN*ANDKURT NORDSTROMDepartment ofMolecularBiology,Odense University, DK-5230Odense M,Denmark
Asmall derivative ofplasmidRl wasused to integratively suppress a
chro-mosomal dnaA(Ts) mutation. The strain obtained grewnormallyat
420C.
The integratively suppressed strain was used as recipient for various plasmid Rl derivatives. Plasmid Rl and miniplasmid derivatives of Rl couldbe establishedin the strainthatcarried an integrated Rl replicon, but they were rapidly lost
duringgrowth. However,plasmidsalsocarryingColElreplicationfunctionswere
almostcompletelystablyinherited. Theintegrativelysuppressedstrain therefore allows theestablishmentof bacteria diploid withrespecttoplasmidRland forms a useful and sensitive system for studies of interaction between plasmid Rl replication functions. Several of thechimericplasmids caused inhibition ofgrowth
athightemperatures. All plasmids that inhibited growth carriedone particular
PstIfragmentfromplasmidRl(thePstI Ffragment),and inallcasesthegrowth inhibitioncould be ascribedtorepressionofinitiation of chromosome replication
at
420C,
i.e., they carry a trans-acting switch-offfunction. Furthermore, theanalogous PstI fragments from different copy mutants of plasmid Rl were
analyzedsimilarly, andonemutant wasfoundtolack the switch-offfunction.The different chimericplasmidswerealso tested for theirincompatibilityproperties. All plasmids that carried the switch-off function (and noother plasmids) also carried Rl incompatibilitygene(s). Sincethe PstI F fragment, which ispresent onalltheseplasmids,isverysmall (0.35x 106),it issuggested that the switch-off regulation ofreplication (by an inhibitor), incompatibility, and copy number controlaregoverned bythesamegene.
Plasmidreplicationinbacteria is being stud-iedintensely,and considerable informationhas been obtained about the biochemistry of the replication process. The use of cloning tech-niques involvingrestriction enzymes has made itpossible toobtaina detailedinsight into the geneticorganizationof thereplication regionsof several plasmids (21, 22, 37-39), and, in a few
cases, including the Escherichia coli
chromo-some, replication origins have been sequenced
(6, 24,40).Severalplasmids, likeFandsomeof thelarge,self-transmissible Rplasmids,are pres-ent in E. coli in numbers similar to the
chro-mosomecopynumber.Therefore,it istempting
tospeculatethat thecontrol of
plasmid
replica-tion andpartitioningmay share several features
with that of the chromosome. Despitethe ease
with whichone canmanipulateplasmids (genet-ically and structurally), very little is known
about these control functions. Two problems haveinterestedusforsometime-copynumber control and incompatibility. The former is
di-rectly connected to the problem ofreplication control, and the latter phenomenon was also
previously foundtobe related tothis problem (42). However, it has been argued that plasmid incompatibilityis part of the segregation func-tions andnotan obligatoryfeature of thecopy
numbercontrolsystem(32).
Inseverallaboratoriesthegenetics of the
rep-lication region of the IncFII plasmids(Ri,R6-5, R100)have beeninvestigated(21, 27,37,38). On the basis ofheteroduplexstudies and restriction
enzymefragmentpatterns,theseplasmidsseem
to be nearlyidentical in thisparticular region. However, somedifferences with respect tothe precise location of the replication genes have been observed:the position and number of the replication origin(s) are not fully agreed upon (21, 28, 37), and also the inc gene(s) have been
located differentlyonthe variousplasmids (21, 37,38).
We have attempted to establish a situation
where the expression of the different control
functions could be analyzed. This was
accom-plishedinastraincarryingadnaA(Ts)mutation in the chromosome into which an Rl plasmid
was integrated. Different parts of the plasmid
111
on January 23, 2021 by guest
http://jb.asm.org/
Ri were then cloned on CoLEl plasmids and
transformedinto thisstrain,where possible
in-terference withreplication of the chromosome
attemperaturesabove40°Cwasanalyzed.The
datatobe discussedhere support thehypothesis
thatcopy numbercontrol, incompatibility,and
regulationof initiation ofplasmidDNA
replica-tion are different manifestations of the same
function.This functionactsintrans,i.e.,it isa
cytoplasmiceffector, andtheexpressionofthis
function doesnotrequirethesimultaneous
pres-enceofafunctional plasmid origin for
replica-tion.
MATERIAILS AND METHODS
Bacterial strains,plasmids, andgrowth
con-ditions. All bacterial strains used are E. coli K-12
derivatives. Theyarelisted in Table1 together with theplasmids used. The growth mediumwaseither LB
(3)orA+Bminimal medium(12),suppliedwith0.2% glucose, 1 ug of thiamineper ml, and 1% Casamino
Acids. Fbr growth ofthymine-requiringstrains, thy-mine was added to 5 jg/ml (low-thymine-requiring
strains). The bacteriaweregrownshakingin thermo-statically controlledwaterbaths, and the increase in celldensitywasfollowed inaZeissPMQ3
spectropho-tometer.
Conjugal mating, transformation and P1 transduction.Conjugaltransfer ofplasmidswas
car-riedoutasdescribed previously (30).Interrupted
mat-ing between Hfr strains and F- recipient strainswas
performedaccordingtoMiller(26). Transformation of E. coli with purifiedDNAwasby amodification of
themethoddescribedby Cohenetal. (13). A culture of cells growing in LB medium was harvested by
centrifugationatadensityof 2x108cells perml. The
cellpelletwassuspendedin 1/10 theoriginalculture
volume of 30 mM CaCl2, and 0.2 ml of suspended cells
wasimmediatelymixed with 0.1 ml of DNA dissolved
in TES
[N-tris(hydroxymethyl)methyl-2-aminoeth-anesulfonic acid]-buffer (10 mMTris, 1mMEDTA,
and 20 mMNaCl, pH 8.0) ataconcentration of
ap-proximately10
itg/ml.
The mixturewaskeptonicefor30min, after which itwasplacedat42°C for2min.
Finally,3ml ofLB mediumwasadded, andphenotypic
expression wasallowed during growth for 1 to 2 h.
After this period the cells were spread onselective
plates. Transduction with P1phagewascarriedoutas
described by Miller (26).
PreparationofDNA,restrictionenzyme
anal-ysis, and molecular cloning of DNA fragments. Plasmid DNApreparation, digestion with restriction
enzymes,agarosegelelectrophoresis, andligation of DNAfragmentswereaspreviously described (27).
Total DNA synthesis.Thymine-requiring strains (LC343) were grown exponentially for several
dou-blings in A+B minimal medium supplied with 0.2% glucose, 1% Casamino Acids, and 5 yg of thymineper
ml. Atthe times indicated in thetext[3H]thymine (30 Ci/mmol, 1 ,uCi/ml) wasadded, and 0.2-ml samples weretakento 10%trichloroacetic acid containing 20 jigofthymineperml. Thesampleswerekept overnight
at4°C. Trichloroaceticacid-precipitatedmaterialwas
collectedonglass fiberfilters, washed extensively with
5% trichloroacetic acidcontaining 20 ,ug of thymine
perml,dried,and counted inatoluene-based scintil-lation mixture.Radioactivitywasmeasured inamark
III NuclearChicago scintillationcounter.
Incompatibility. The various chimeric plasmids
were transferred by transformation toE. coli C600 (Rldrd-19). Selectionwasfor theincoming plasmid
only. One colony waspurified to single colonies on
TABLE 1. Bacterial strains andplasmids
Designation Parent Genotype' Source,
refer-ence E.coli strain
1100 endI 15
LC343 leu thy pro lac dnaA46 4
LC343int500 LC343 leuthypro lac dnaA46aphA This paper
EC1005 metrelA nalA 17
W3110poLA W3110 thydrapolA 14
Gil HfrC met ilv 16
RH491 leu trphislysilv rpsL 20
AB2295 AT2255 leu his arg met thy lacY mal 43
xyl gal mtl thi rpsL tonA
Plasmid
Rldrd-19 bla+ cat' aphAM sul+ tra+ 25
Rldrd-19K-1 Rldrd-19 bla+ cat'aadA+sul'tra 2
pKN500 Rldrd-19 aph+ 27 pKN501 Rldrd-19 bla+ 27 pKN800 Rldrd-19 bla+ aph+ 27 pKN1562 Rldrd-19 aph+ 27 pKN520 pKN102 bla+ cop 41, 42 pKN182 pKN104 cop 42
pBR322 ColEl tet+bla+ 7
pKN191 ColE1/Rldrd-19 bla+ 27
aGenesymbolsaccording to Bachmann et al. (1) and Novick et al. (31), cop, copy number.
on January 23, 2021 by guest
http://jb.asm.org/
plates selecting again for the incomingplasmid, and 50 to 100coloniesweretested for the presence of the
resident plasmid (resistance pattern). In all cases
where a chimeric plasmidwas classified ascarrying
inc, more than98% of the tested clones had lost the residentplasmid.
Chemicals, isotopes,and enzymes.Allchemicals
used wereanalysis grade. Agarose, RNase,lysozyme,
and antibiotics werepurchasedfromSigmaChemical Co. (St. Louis, Mo.). Restrictionendonucleases were obtained from C. F. Boehringer & Soehne GmbH,
Mannheim, West Germany,and T4 DNAligasewas
obtained from Miles Laboratories Ltd.(Slough,
Eng-land). Radioisotopes were obtained from The
Radi-ochemical Centre (Amersham, Buckinghamshire,
England).
RESULTS
Integrative suppression ofadnaA muta-tionbyaminiplasmidderived from Rldrd-19. We havetested different smallplasmid de-rivativespreviouslyisolated from Rldrd-19(R1) for theircapacity tosuppress the dnaA46
mu-tation (28). The differentplasmids were trans-forned to strain LC343, and clones growingat 42°C were analyzed. It was found that only if theplasmidcarried EcoRIfragmentF(theKmr fragment,seeBlohm and Goebel [5]) or part of it was the frequency of temperature-resistant clonessignificantlyincreasedcomparedwith the frequency of reversion of the temperature
sen-sitivity ofLC343having no plasmid. For exam-ple, temperature-resistant clones of LC343 were found with afrequency ofapproximately 10-6;
the same wasthe caseifplasmidpKN501 was
present inLC343; but thepresence ofplasmid pKN500 increased this frequency to approxi-mately 10-4. The two plasmids, pKN500 and pKN501, both have EcoRI fragment B ofRl, but the former plasmid has, in addition, the EcoRI Ffragment,and thelatter has the EcoRI Dfragment(ampicillin resistance) (27).
One temperature-resistant clone of LC343 harboring plasmid pKN500 (LC343int5O0) was
analyzedfurther. Severalexperimentswere
car-ried out to showthatplasmid pKN500 is inte-grated into the chromosome of LC343int500. Figure 1 presents the profiles of dye buoyant density gradients(CsCl-ethidium bromide)with total cell extracts of LC343 carrying plasmid pKN500(Fig. 1A) andofLC343int500(Fig.1B).
Intheformercase adistinctpeak corresponding
tocovalentlyclosedcircular moleculeswasseen, whereasthiswas notobservedin thelattercase.
Both strains carried thekanamycinresistance of pKN500. The absence ofautonomous plasmid moleculeswasfurther demonstrated in incom-patibility testsin which akanamycin-sensitive derivative of
plasmid Rl,
Rldrd-19K-1(2),
was transferred by conjugation to either -L 4._ '-5 1 50 CL~~~~~~~~~~~~~~~4 3~~~~~~~~~~~~~~~~3 2 20 1 10 0 20 30 Fraction no.
FIG. 1. Density gradient profiles of DNAfrom(A)
LC343(pKN500)and(B) LC343int500. CellsofLC343
(harboringplasmid pKN500)andLC343int500
grow-ingexponentiallyin LBmediumat30°Cwerelabeled
overthreecell doublingswith(6H]thymine(5
ACi/ml,
30 Ci/mmol). Total cell lysates wereprepared and
analyzedonethidiumbromide-CsCIgradientsas
de-scribed in the text. Eachgradient wasfractionated
into 40fractions,25,ulfromeachfractionwasspotted
ontofilterpaper, and thefilterpaper was washed
threetimes in 5% trichloroacetic acid and driedat
110°C. The individualfractionswerecounted in
tol-uene-containing Omnifluor countingfluid.The
frac-tions from the bottoms ofthe gradients have the lowestfractionnumbers. The broken lines indicate where the ordinateschangescale. Thearrows
indi-catetheposition ofcovalentlyclosed circular DNA.
LC343(pKN500) orLC343int500. Inthe former
casetheclones found lostplasmid pKN500with high frequency after growth without selec-tion for any plasmid. However, in case of LC343int500, onlyloss of Rldrd-19K-1 was ob-served. Thissupports the conclusion thatnofree
plasmid (carrying kanamycin resistance) is
present in the temperature-resistant strain LC343int500.
141,
on January 23, 2021 by guest
http://jb.asm.org/
114 MOLIN AND NORDSTROM
Kanamycin resistance was transduced by phage P1 from strainLC343int500tothe HfrC strain Gll. One transductant clone of Gllwas
analyzed on densitygradientsforthe presence
of free plasmid DNA, as described above, and
nocovalently closed circular molecules could be detected. This strainwasthenmated with the F- strainAB2295, and the time ofentryofKmr
wasdetermined. Thekanamycinresistancewas
transferred at30minaftermixingof donor and recipient cells; before this time no kanamycin-resistant AB2295 cellswereobserved.Since Gll initiates thechromosome transferat 10min on
thegeneticmapof E. coli(1) andtransfers the markers in acounterclockwise orientation, the result of the interrupted mating experiment shows thatpKN500 isintegratedatapositionof approximately80minonthemap.
Finally, we have been ableto showthat the dnaA(Ts) mutation ofLC343 is still present in the temperature-resistant strain LC343int500: P1phagegrown onstrainLC343int500wasused
to transduce strain RH491, selecting for Ilv' recombinants. Approximately 3% of the Ilv' transductants were temperature sensitive, and
one of these showed the same characteristic change in growth rate aftera shift from 30 to
420Casthat shown forLC343 inFig.2.
Physiological characterization of LC343-int500 harboring a fused ColEl-Rl
repli-con.StrainLC343int500growswellatall
tem-peratures (30 to
420C).
Figure 2 shows howstrains LC343 andLC343int500respondedto a
temperature shift from 30 to
420C.
AlthoughLC343 continuedto growfor threetofour cell doublings,thegrowthrate wascontinuously
re-ducedcomparedwith that ofLC343int500. Mi-croscopic observations revealed that the cells of LC343 became very long ("snakes") during growth at 420C, whereas cells ofLC343int500 retainedthe normal cellshapeobservedfor both strainsat300C (datanotshown).
We transferred several
Ri-related
plasmidsto LC343int500. AllRl-derived plasmids that didnotcarryotherreplication functions thanthose
of
Ri
were lost rapidly from the cells whengrownwithout selection for their maintenance.
However, plasmids consisting of multiple repli-cation originswere foundto be more stablein
LC343int500. We analyzed in some detail how different plasmids consisting of ColEl replica-tionfunctionsaswellasRireplication functions affected thegrowth ofLC343int500.
One series ofexperimentswasconcernedwith plasmid pKN191 (27), which consists of the ColEl-like plasmid pSF2124 (36) fused to the EcoRl-Bfragmentof
Ri,
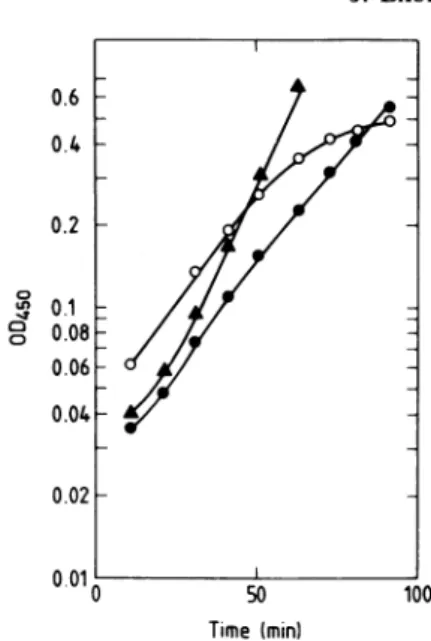
knowntocarry all thereplication functions. 0.6 0.4 0.2 C c] 0 0.1 : 0.08 - 0.06-0.04 0.02 0.01 0 50 100 Time (min)
FIG. 2. Cell growth of strains LC343 (0), LC343int500(A),andLC343int5OO(pKN191) (0) after
atemperatureshiftto42'C. Culturesofthe strains growingexponentiallyin LB mediumsupplied with
10pg of thymineper mlat 30'C werediluted into
prewarmed (42'C)medium,andgrowthwasfollowed
spectrophotometricallyin a ZeissPMQ3
spectropho-tometer at 450 nm.The time0min represents thetime
ofthetemperatureshift.
When cultures of LC343int500 harboring pKNl91weregrownfor10 to 20generationsat
300C, we found that 80 to 90% of the cells retained theplasmid despite the absence of
se-lectionpressure.Thegrowthrate wasthesame as that of LC343 or LC343int500 having no
plasmids,but afteratemperatureshiftto420C,
a difference between cells ofLC343int500 with andwithout pKNl91 becameclear(Fig. 2): the plasmid-containingcellsgrew moreslowlythan LC343int500 but did not come to a complete halt inmassincreaseas wasobservedfor LC343. We further analyzed LC343int500 harboring plasmid pKN191 withrespect to the effect on DNAreplication (chromosome) ofthe
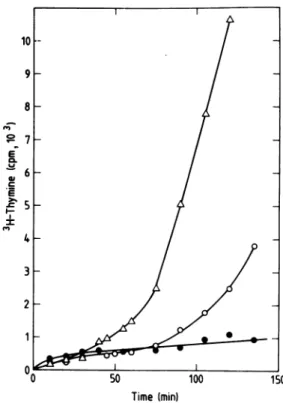
temper-ature shift. The presence of plasmid pKN191
was found to cause a reduction ofthe rate of DNAreplicationinLC343int500 whenthe
tem-perature wasincreasedto420C(Fig.3).(Wewill
refertothiseffectby using the term
switch-off.)
Thepresence ofplasmid pSF2124 hadnoeffect
atall onDNAsynthesis or growth when
ana-lyzedinsimilarexperiments.
Figure 3 also shows that the repression of
DNAsynthesis after the temperature shift was
only transient; after 75 to 90 min replication started again. A possible explanation for this
comes from an analysis of the distribution of
on January 23, 2021 by guest
http://jb.asm.org/
REPLICATION FUNCTIONS OF Ri 115
Chimeric plasmids carrying replication functionsofRl. We haverecentlydescribeda
series ofsmallplasmid derivatives from Rl that
all share adistinct regionofthe EcoRI B
frag-ment (27). These plasmids seem to exhibit all
replicationfunctionsofthewild-typeRlparent plasmid, including copynumber control and in-compatibility functions. Toidentify genetically
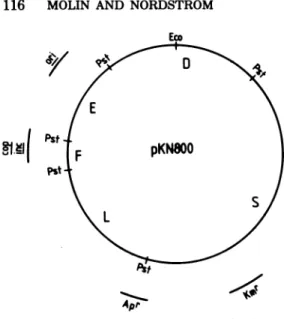
the regiononRl where thefunctioninhibiting DNAsynthesis maps,weclonedthevarious PstI fragmentsfromplasmid pKN800onthecloning vehicle pBR322 (7). As described previously (27), plasmid pKN800carriesasmallpartofthe EcoRIBfragmentfromRldrd-19 togetherwith the kanamycin resistance gene andpartof the TnA transposon. Therestriction enzymemap of
pKN800 is presented in Fig. 5. We obtained a
series of chimeric plasmids with each of the
single fragments as well as different
combina-tions offragments clonedonpBR322 (Table2).
Only one type of chimeric plasmid could be
established in strain W3110poLA: pKN271, which,inadditiontopBR322, carries the PstIE
and F fragments. This result agrees well with
datafrom acopy mutantof Rlpreviously
pub-Time(min)
FIG. 3. Total DNAsynthesis in strains LC343(0),
LC343int500(A),andLC343int500(pKN191) (0)after
a temperatureshiftto42°C. Cultures growing
expo-nentiallyin A +B minimal mediumsuppliedwith
0.2%glucose,1%CasaminoAcids,and5
jg
ofthymineper ml at 30°C were diluted intoprewarmed medium
(42°C) containing [3H]thymine (2
ltCi/ml,
30 Cilmmol)at 0min.Samples of0.1mlwere taken to
ice-cold 10% trichloroacetic acid containing 20pg of
thymineper ml.Thetrichloroaceticacid-precipitated
material wasanalyzed forradioactivityasdescribed in the text. The optical density at 450 nm ofthe dilutedculturesattheinitiationof labelingwas 0.1.
cells with andwithoutplasmidDNA(pKN191). Asstated above, there was a small fraction of thecells ofLC343int500(pKN191)fromcultures growingat
300C
withoutselectionpressurethat had lost theplasmid, and thisfractionwasnor-mally10to20%.(This
instability
isnotrestrictedto this particular strain; in normal E. coli cell
linesplasmidpKN191 is lost withasimilar fre-quency,and thecopynumber of theplasmidhas
beenfoundtobeapproximatelytwocopiesper
genome equivalent, i.e., much lower than
nor-mallyfound forColElplasmids.)Whenaculture of strain
LC343int500(pKN191)
wastransferredto42°C,the fractionofcellsharboringthe plas-mid decreased (Fig. 4). After2hless than 50% of the cells carriedplasmidpKN191,andatthat time the rate ofDNAreplication was close to
halfthat oftheplasmid-freecontrol strain (Fig.
3). .0 0 CL 0 GD a co C ._ %6-0 c:s c 80 60
401
20 10 8 6 4 2 0 1 2 3 Time (hrs)FIG. 4. Loss of plasmid pKN191 from strain
LC343int500 aftera temperature shift to420C. An
exponentially growing culture ofLC343int500
(har-boringplasmidpKN191) in LB medium(30°C) was
transferredto420Cat0min.Samples of the culture
werespreadatdifferent timestogivesingle colonies
onLAplates thatwereincubatedat300C. Fromeach plate 50 colonieswerepatchedtoplatescontaining 50pgof ampicillinper ml.Thefraction of coloniesat each timepoint thatwasresistanttoampicillinwas
taken as a measure of the fraction of cells in the
culturewhich containedplasmid pKN191. 10 E I-c .1_ 0 0 0 0 0 4 C VOL. 141,1980 on January 23, 2021 by guest http://jb.asm.org/ Downloaded from
Apr
S
41 FIG. 5. Restriction enzyme map of plasmid
pKN800. Themap wasobtainedasdescribed previ-ously(27).The letters(E,D,S,L,andF) refertothe different PstI (Pst) fragments. Theposition ofthe replication origin(ori)wastakenfrom Kolleketal. (21). The position ofthe cop orincgene on the F fragment is basedonthe presentcommunication.Eco
isthe siteforEcoRL
lished by Kollek et al. (21). After transfer of
these various chimeric plasmids to strain
LC343int500, the different transformant clones
werescreened for thepresenceof switch-off(of
replication) functions on the plasmids. It was
found thatsimply testing for growthat420Con
platescontaining tetracycline (selecting for the
maintenance oftheplasmids) wassufficientto
identify the plasmids containing the switch-off
functions. The presenceofseveral plasmids
re-sultedinlossofviabilityofLC343int5OOat420C
ontetracyclineplates, and alloftheseplasmids
contained the PstI F fragment. To verify the
presence ofswitch-off functions on these
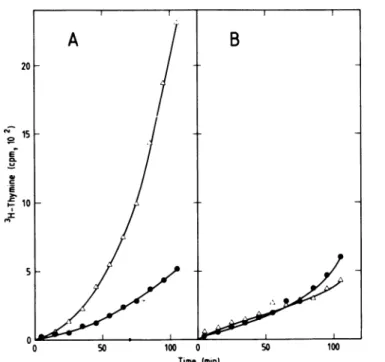
plas-mids, weanalyzed the kineticsofchromosome
replication afteratemperature shift from 30to
420C for each of the strains. Figure6showsone
example of such an experiment. Plasmid
pKN317, which carries the PstI F fragment, had
a strong inhibitory effect on the rate ofDNA
synthesisatthehightemperature.
Therepression of DNA synthesisat420Cwas
maintained, causinga5- to 10-fold reduction in
therateofincorporation of [3H]thymine. A
sim-ilarresultwasobtainedwithachimeric plasmid
carrying the PstI F fragment in the opposite
orientation compared with that of plasmid
pKN317.
No other PstI fragment cloned on plasmid
pBR322 was foundtointerfere with growth or
DNAsynthesis in thistypeofexperiment. The
effect ofplasmid pKN271 (PstI-E+F)was
simi-lartothatobserved forplasmid pKN191; after
an initial inhibition DNA synthesis was later
gradually resumed, andagain itcould be shown
that this plasmid was lost from the cells in a
fashion similar towhatwas found for plasmid
pKN191.
Incompatibility properties of the pBR322 chimeric plasmids carrying replication functions of plasmid Rl. The plasmids
carry-ing thedifferentPstIfragments from the repli-cation region of Rlweretested in two typesof incompatibility experiments. First, each chi-meric plasmidwastransformedtoE. coli C600 carrying plasmidRldrd-19, and the incompati-bilitytest wascarriedout asdescribed in Mate-rials and Methods. Itwasfoundthatall plasmids
carrying the PstI F fragment exerted a strong
incompatibility onRldrd-19, whereas all other plasmids were completely compatible with
Rldrd-19(Table 2).
Second, the frequency of transfer
(conjuga-tion) of Rldrd-19tocellscarrying the chimeric plasmids was measured. In these experiments the number of coloniesgrowingonplates selec-tive for both theincomingplasmidand the
res-identplasmidwasdetermined. Thepresenceof pBR322plasmids carrying thePstI Ffragment
TABLE 2. Properties ofchimericpBR322plasmids
PstI
frag-ment
Switch-Plasmid clonedon Parentplasmidb Inc' Switch
plasmid pBR322 pBR322 None pKN800(Cop4) - -pKN271 E+F pKN800(Cop4) + + pKN228 E pKN800(Cop4) - -pKN317 F pKN800(Cop+) + + pKN231 D pKN800(Cop+) - -pKN229 L pKN800(Cop') - -pKN292 S pKN800(Cop') - -pKN431 F pKN520(Cop-) + + pKN388 F pKN182(Cop-) -
-Theletters refertothose used inFig.5.
b Parentplasmidsarethose usedassourcesof
PstI
fragments.Inparentheses is indicated the copy
num-berphenotypeof the parentplasmids:Cop',wild-type
copy number, Cop-, increased (copy mutant) copy number.
'Incompatibilitywastested asdescribed in the text.
Symbols: +,none out of50tested colonies retained
the residentplasmid; -,alloutof 50 testedcolonies retained theresidentplasmid.
dSwitch-offwastested instrainLC343int500(i) as
abilitytogrowonLAplatescontaining tetracycline at
42°Cand (ii) ascapacity to reduce the rate of DNA
synthesis at42°C (see Fig.6). Plasmidsresulting in
temperaturesensitivityforgrowthandDNAsynthesis
areindicatedby+;plasmidsthat donotinterfere with
growthorDNAsynthesisareindicatedby-.
on January 23, 2021 by guest
http://jb.asm.org/
REPLICATION FUNCTIONS OF Ri 117 52 =o ,- 10
z7
0 50 100 0 50 100 Time (min)FIG. 6. Total DNAsynthesis in the strainsLC343int500(pBR322) (A) andLC343int5OO(pKN317) (B) after a temperatureshiftto42°C. Whenculturesof LC343int500 (harboring plasmid pBR322)andLC343int500
(harboringplasmidpKN317) exponentially growingin LB mediumsuppliedwith 0.2%glucoseand 20pgof
thymineper ml reachedanopticaldensityat450nmof 0.1,[3HJthyminewasadded(5,uCi/ml,30Ci/mmol),
andhalfthe culture wastransferredto42°C, whereas the other halfwaskept at 30°C.From both the 30°C cultures(0)andthe42°C (A)culturessamplesweretakentotrichloroaceticacid,andgrowthwasmonitored
spectrophotometrically(optical densityat450nm). Incorporation of [3HJthyminewasanalyzedasdescribed
inthelegendtoFig.3.
reduced the transfer frequencyto
io-
of that ofplasmid-free recipient. No other pBR322 plas-mids hadanyeffectonthetransfer ofRldrd-19.
Thepresenceofplasmid pKN500 (wild-type Rl
replicon,lowcopynumber) in the recipient cells reduced the transferbyafactor ofonly20 to 50.
Analysis ofPstIfragments fromcopy
mu-tantsofRldrd-19. The PstI Ffragmentsfrom
twodifferentcopy mutantplasmidsofRldrd-19
wereclonedonplasmid pBR322. Thetwo copy
mutant plasmids, pKN102 and pKN104, have
beenshowntobeof differenttypesin incompat-ibility experiments (42). The PstI fragments
werecloned from theminiplasmidspKN520 and pKN182, derived from the two copy mutant
plasmids. Plasmid pKN520 (derived from
pKN102) was constructed in vitro and consists
ofthetwoEcoRIfragmentsB and D (41).
Plas-midpKN182 is aspontaneously generated
min-iplasmid from plasmid pKN104 carrying only
thereplication region (including the PstI
frag-mentsF, E,andD)from the parentplasmidbut
noantibiotic resistance gene. TheclonedPstI F
fragments were identified by their molecular weightsand thepresence ofaSalI sitecloseto
onePstIsite (38).
The resulting chimeric plasmids, pKN388 (parentplasmidpKN104) and pKN431 (parent plasmid pKN102), were testedfor switch-off of replicationfunctions and incompatibility
prop-erties asdescribed above. The PstI Ffragment
from thie copy mutantpKN102 inhibited DNA
replication in strain LC343int500 at 42°C in a
mannerverymuchsimilartothatobserved for
plasmidpKN317(wild type), and also the incom-patibility reaction was indistinguishable from that ofplasmidpKN317.In contrast,the PstIF
fragment from thecopy mutantpKN104 hadno
measurable effectonRl replication and
main-tenance in these tests. These results are
pre-sented in Table2.
Thus, inonetype ofcopy mutant (pKN102)
the mutation is probably located outside the
PstI F fragment, whereas in case of plasmid pKN104 the cop mutation is within this
frag-mentandisclearlycausing lossof theswitch-off function.
DISCUSSION
Integrative suppression ofadnaA
muta-tion. Several self-transmissibleplasmids,likeF, VOL. 141,1980
on January 23, 2021 by guest
http://jb.asm.org/
118 NORDSTROM
may be inserted intothechromosomeof E.coli,
giving risetoHfr strains (8).Oneway to obtain
such strains is by integrative suppression of
dnaA(Ts) mutations (28,29).Whentheplasmid
isintegratedas partof the chromosome in these strains, it is found thatreplication of the
chro-mosome atthe restrictive temperatureis
initi-ated from the integrated plasmid origin (4, 9, 11). Thus, at a temperature where initiation from the chromosomal origin is blocked, the whole chromosomemaybe consideredas agiant replicating plasmid.
We have chosenminiplasmidsforintegrative suppressionfirst ofalltoreduce the number of potential functionalreplication origins,since R plasmids (R100) have been claimed to contain several origins (33).Furthermore,itwasrecently shown(10) thatplasmidR100, whenintegrated intothe chromosome ofadnaA(Ts) strain,
ex-cisedplasmid molecules identicaltother deter-minant of theplasmid,andthepresenceof such covalentlyclosed circular moleculeswould have disturbedsomeofour measurements.The strain used in allexperimentswasshowntocarrythe inserted Rlplasmid in theregionofthenormal chromosomaloriginandtocarrythe dnaA(Ts) mutation.
Controlofreplicationof Rl. Whenplasmid
Rl(pKN500)isintegrated into the chromosome ofLC343, theresultingstrain(LC343int500)
pos-sesses two origins that are functional at 300C
andone (Rl) that is functionalat400C. We do
notknow whichorigin isbeing usedatthelow
temperature, but since theamount of DNA in
LC343int500 is notsignificantly different from thatinLC343(datanotshown),wemay assume thatthetwo originsare notboth activeduring the cell cycle. Transfer of various Rl plasmid derivatives to LC343int500 does not seem to
interfere with replication of the chromosomeat
300C. This means that the process of DNA
elongation isnotaffectedby the controlsystem
of Rl. However, after a temperature shift to
420C,
whenRl takesoverthe control ofinitia-tionofreplication of thechromosome, the
pres-enceof several Rlplasmidderivatives was found
to interfere with chromosome replication. We
have used the term
switch-off,
introduced by
Pritchard, for this type of inhibition of DNA synthesis (34, 35). Our experiments do not
di-rectly revealthe nature of the switch-off
func-tion, but the fact that it acts in transsuggests
that either atranscriptionor atranslation
prod-uct is responsible for the switch-off of
replica-tion.
The presence of the trans-acting switch-off
function was first of all demonstrated from a
composite plasmid consisting of plasmid
pSF2124 and the EcoRI Bfragment ofplasmid Rl.However, since such fused replicons turned
out tobe unstably maintained eveninnormal
E. coli strains (apparently due to a much
re-ducedcopynumber compared with that of
plas-mid pSF2124), the analysis of the switch-off
function iscomplicated by the factthat a
grow-ing population of cells is amixture of plasmid-free andplasmid-carrying cells. The reasonfor
the apparently defective replication of these
plasmidsis atpresent not known.
Thefractionation of the replicationregion of
plasmid Rl into several small PstI fragments thatwereclonedindividuallyonthevector plas-mid pBR322 revealed thataDNAfragment of
the size0.35 x 106daltons (PstI-F) carries the
switch-off function, and no other part of the replication region could be shown to interfere with Rlreplication.We thereforeconcludethat a geneinvolved in the control of replication of plasmid Rl is located on the PstI F fragment and that the function expressed from thisgene actsintrans. This conclusionwassupported by thefinding that the PstIFfragmentfroma copy
mutantplasmid (pKN104) expressedno
switch-offfunction, i.e., thecopmutation ofthis plasmid liesin the switch-offgene.Incompatibility func-tion(s)wasalso showntobelocatedonthePstI F fragment, and again no inc activity was
ex-pressedfrom the PstIFfragment derivedfrom
thecopy mutantplasmid pKN104. This directly
shows thatat leastpart of the incompatibility phenotype isconcerned withreplicationcontrol.
Theassumption thatonlyonefunction is
re-sponsible for switch-off, incompatibility, and
copynumbercontrol doesnotnecessarilymean
that no othergenes orDNAsequences are in-volved in these controlfunctions. In contrast, we
havealso shown thatonetype of copymutant
(representedby plasmid pKN102)harbors a
mu-tation outside the switch-offfunction, as
dem-onstrated for plasmid pKN431. Furthermore, this particular mutantsegregates neither from cells containing Ri (42) norfromcellscarrying
the switch-off function on multicopy plasmids (e.g., pKN317 [datanotshown]).Theseresults
are in accordance with thephenotype ofa
mu-tation in the target for the replication control
function. Only genetic recombination
experi-ments will provide information concerning the
locationof such a region.
We have recently proposed a model for the
control of Rl replication in which it is argued
that two principally different control circuits
participate: oneisacis-actingswitch-off
mech-anism that prevents foraconsiderable time
rein-itiation of replication on a plasmid molecule
which hasjustfinisheditsreplication cycle.The
on January 23, 2021 by guest
http://jb.asm.org/
other isatrans-acting control determining the
overall replication rate (atthe initiation level)
by means of a cytoplasmic inhibitor molecule
(18). Theformermechanism preventsmultifork
initiationonthesamemolecule, the latteris the
mechanismresponsiblefor copynumber control, and, as discussed in the accompanying paper
(19), the properties of the replication control
systemshareseveral features with models
pro-posinga negative control (23, 24, 35). We have
in the present communication described some
features of the trans-acting control.
Concluding remarks.Recently, Timmiset
al. (38) andTaylor and Cohen (37) showed that
onePstI fragment from thereplication region of plasmid R6-5 (an IncFII plasmid, like R1) carries
a gene the function of which is similar to the switch-off (inc, cop) function described in the
presentcommunication.Although the PstI
frag-ment from plasmid R6-5 (and R100) is larger than the PstIFfragment of Rl, thepartof the
R6-5fragment which islocated adjacenttothe fragment equivalenttothe PstI Efragment
ap-pears identical to the PstI F fragment of Rl.
This conclusion is based onthe physical maps
presented by Timmis et al. (38), Kollek et al. (21), andTaylor and Cohen (37). However,
in-vestigations of smallplasmids derived from the
Rlcopy mutantpKN102 carriedoutby Kollek
etal. (21) indicate the position ofthe incgene onthe PstI Dfragment, whereasno detectable incompatibilitywas expressed from the PstI F
fragment.Itisnotclearatpresentwhatcanbe
thereasonfor thisdiscrepancy. We have shown
thatincompatibility, copynumbercontrol, and replication switch-off functions are located on
thesamesmallpiece of DNA,and,inagreement
with the results of Timmisetal. (38),noother DNA fragment from this region contains any
observable function related to incompatibility andreplication control. We therefore conclude that one gene product is responsible for copy
numbercontrol,incompatibility,and switch-off ofreplication.
ACKNOWLEDGMENTS
Our workwassupported bythe Danish Medical Research Council(projects8000and10149).
Theexcellenttechnical assistance of EvaHeynOlsen and LisLyngsieishighly appreciated.
LITERATURE CITED
1.Bachmann,B.J.,K.B.Low,and A.L.Taylor.1976. Recalibrated linkage map of Escherichia coli K-12. Bacteriol. Rev. 40:116-167.
2. Beard,J.P.,andJ.C.Connoily.1975.Detectionofa
protein, similartothesex pilussubunit, in the outer membrane ofEscherichia colicellscarrying a dere-pressed F-like R factor.J.Bacteriol.122:59-65. 3. Bertani, G.1951.Studiesonlysogenesis.I.The mode of
phage liberation bylysogenic Escherichia coli. J. Bac-teriol. 62:293-300.
4. Bird, R. E., M.Chandler, and L. Caro. 1976. Suppres-sion of an Escherichia coli dnaA mutation by the integrated Rfactor R.100.1: change of chromosome replication origin in synchronized cultures. J. Bacteriol. 126:1215-1223.
5. Blohm, D., and W. Goebel. 1978. Restriction map of the antibiotic resistance plasmid Rldrd-19 and its deriva-tives pKN102(Rldrd-19B2) and Rldrd-16 for the en-zymes BamHl, HindIII, EcoRI and Sal. Mol. Gen. Genet. 167:119-127.
6. Bolivar, F., M. C.Betlach, H. L. Heyneker, J. Shine, R.L. Rodriguez, and H. W. Boyer. 1977. Origin of replication of pBR345 plasmid DNA. Proc. Natl. Acad. Sci. U.S.A. 74:5265-5269.
7.Bolivar, F., R. L. Rodriguez, P. J. Greene, M. C. Betlach, H. L. Heyneker, H. W. Boyer, J. Crosa, and S. Falkow. 1977. Construction and characteriza-tion of newcloning vehicles. II.Amultipurpose cloning system.Gene 2:95-113.
8. Broda, P. 1967. The formationofHfr strains in Esche-richiacoli K-12. Genet. Res. 9:37-47.
9. Chandler, M.,L.Silver,andL.Caro.1977.Suppression ofanEscherichia coli dnaA mutation by the integrated Rfactor R100.1: origin of chromosome replication dur-ing exponential growth. J. Bacteriol. 131:421-430. 10.Chandler, M., L.Silver, J.Frey,and L. Caro. 1977.
Suppressionof an Escherichia coli dnaA mutation by the integrated R factor R100.1: generation of small plasmids after integration. J. Bacteriol. 130:303-311. 11.Chandler, M., L Silver, Y. Roth, and L. Caro. 1976.
Chromosome replication inanHfr strain of Escherichia coli. J. Mol. Biol.104:517-523.
12.Clark,J.D., and 0.Maal0e.1967.DNAreplication and thedivisioncycle in Escherichia coli. J. Mol. Biol. 23: 99-112.
13.Cohen, S. N., A. C. Y. Chang, and L. Hsu. 1972. Nonchromosomal antibiotic resistance in bacteria: ge-netic transformation of Escherichia coli by R-factor DNA.Proc. Natl. Acad.Sci.U.S.A.69:2110-2114. 14.DeLucia, P., and J. Cairns.1969.Isolation of an E. coli
strainwithamutationaffecting DNA polymerase. Na-ture(London) 224:1164-1166.
15. Diirwald, H., and H. Hoffmann-Berling. 1968. Endo-nucleaseI-deficientand ribonucleaseI-deficient Esch-erichia colimutants.J.Mol. Biol. 34:331-346. 16. Eriksson-Grennberg,K. G.,H.G. Boman, J. A. T.
Jansson,and S. Thoren.1965.Resistance of Esche-richia coli to penicillins. I. Geneticstudy of some am-picillin-resistantmutants.J. Bacteriol. 90:54-62. 17. Grinsted, J., J. R. Saunders, L. C. Ingram, R. B.
Sykes,and M. H. Richmond.1972.Properties ofan Rfactor whichoriginated in Pseudomonasaeruginosa 1822.J. Bacteriol. 110:529-537.
18. Gustafsson, P.,H.Dreisig,S.Molin,K.Nordstrom, and B. E. Uhlin.1979.DNAreplicationcontrol: studies ofplasmidRl.ColdSpringHarborSymp. Quant.Biol. 43:419-425.
19.Gustafsson, P., and K. Nordstrom. 1980.Control of plasmidRlreplication: kinetics ofreplicationinshifts between differentcopynumberlevels. J. Bacteriol. 141: 106-110.
20. Hoess,R.H.,andR. K.Herman.1973.Segregationof induced frameshift mutations and thesequence of gene replicationinEscherichia coliK-12.Genetics 74:227-242.
21. Kollek, R., W.Oertel,and W. Goebel.1978.Isolation and characterizationof theminimalfragmentrequired for autonomousreplicationof acopymutant(pKN102) of theantibioticresistance factorRI. Mol. Gen. Genet. 162:52-58.
on January 23, 2021 by guest
http://jb.asm.org/
22.Manis, J. J., and B. C.Kline. 1977. Restriction endo-nucleasemapping and mutagenesisof theFsexfactor replication region. Mol. Gen.Genet. 152:175-182. 23. Matsubara, K.,and Y. Takeda. 1975. Role of thetof
gene in the production and perpetuationof the Xdv plasmid. Mol.Gen. Genet. 142:225-230.
24. Meijer, M., E. Beck, F. G. Hansen,H.E. N.Bergmans,
W.Messer, K.vonMeyenburg, andH.Schaller.
1979.Nucleotidesequenceoftheoriginofreplicationof the Escherichia coli K-12 chromosome. Proc. Natl. Acad. Sci. U.S.A. 76:580-584.
25. Meynell, E., and N. Datta. 1967. Mutantdrug-resistant factors ofhigh transmissibility.Nature(London)214: 885-887.
26. Miller, J.H. 1972. Experimentsin moleculargenetics.
ColdSpring HarborLaboratory,ColdSpringHarbor, N.Y.
27.Molin, S., P. Stougaard, B.E.Uhlin,P.Gustafson, and K.Nordstrzim. 1979. Clusteringofgenesinvolved inreplication, incompatibility,and stablemaintenance of the resistanceplasmid Rldrd-19.J. Bacteriol. 138: 70-79.
28. Nishimura, A., Y. Nishimura,andLCaro.1973. Iso-lation of Hfr strains from R+and ColV2'strains of Escherichia coliand the derivation ofanR'lac factor by transduction. J.Bacteriol.116:1107-1112. 29. Nishimura, Y.,L.Caro,C. M.Berg, and Y.Hirota.
1971.ChromosomereplicationinEscherichiacoli. IV. Control of chromosomereplicationandcelldivisionby
anintegrated episome.J. Mol. Biol.55:441-456. 30. Nordstrom, K.1971. Increased resistance to several
an-tibioticsbyonemutation inanR-factor, Rla.J.Gen. Microbiol. 66:205-214.
31. Novick, R. P., I. C. Clowes, S.N.Cohen,R.Curtiss
III,N.Datta,and S. Falkow. 1976.Unifonn
nomen-clature for bacterial plasmids: a proposal. Bacteriol.
Rev. 40:168-189.
32. Novick, R. P.,and F. C.Hoppensteadt.1978. On plas-midincompatibility. Plasmid 1:421-434.
33. Perlman, D.,andR.H.Rownd. 1976.Twooriginsof replication in composite R plasmid DNA. Nature
(Lon-don) 259:281-284.
34. Pritchard, R. H. 1978. Control of DNA replication in bacteria. p. 1-26. In I. Molineux and M. Kohiyama (ed.), DNAsynthesis, present and future. NATO Ad-vanced Study Institute SeriesA: Life Sciences. Plenum Publishing Corp., New York.
35. Pritchard, R.H., P. T. Barth, and J.Collins. 1969. Control ofDNA synthesis in bacteria. Symp. Soc. Gen. Microbiol. 19:263-297.
36. So, M., R. Gill, and S. Falkow. 1975. The generation of aColEl-ApR cloning vehicle which allows detection of inserted DNA. Mol. Gen. Genet. 142:239-249. 37.Taylor, D.P.,andS. N. Cohen. 1979.Structuraland
functional analysis of cloned DNA segments containing thereplication and incompatibility regions of a mini-plasmid derived from a copynumber mutant of NR1. J. Bacteriol. 137:92-104.
38. Timmis,K. N., I.Andres, and P. M. Slocombe. 1978. Plasmidincompatibility: cloning analysis of an incFII determinant of R6-5. Nature (London) 273:27-32. 39. Timmis, K. N., F. Cabello, and S. N. Cohen. 1975.
Cloning, isolation and characterization of replicating regions of complex plasmid genomes. Proc. Natl. Acad. Sci.U.S.A. 72:2242-2246.
40.Tomizawa, J., H. Ohmori, and R. E. Bird. 1977. Origin ofreplication of colicin El plasmid DNA. Proc. Natl. Acad.Sci. U.S.A.74:1865-1869.
41. Uhlin, B. E., P. Gustafsson, S. Molin, K. Nordstrom, and D. Blohm. 1979. Replication and copy number control of theplasmid Rl inEscherichiacoli,p.33-45. InS.Mitsuhashi (ed.), Microbial drug resistance, vol. 2. Japan Scientific Societies Press, Tokyo.
42.Uhlin, B. E., and K.Nordstrom. 1975. Plasmid incom-patibilityand control ofreplication:copymutants of the R-factor Rl inEscherichia coli K-12. J. Bacteriol. 124:641-49.
43. Wehr, C. T., L Waskell, and D. A. Glaser. 1975. Characteristics ofcold-sensitive mutants ofEscherichia coliK-12defectiveindeoxyribonucleicacidreplication. J.Bacteriol. 121:99-107.
on January 23, 2021 by guest
http://jb.asm.org/