Imaging Brain Function in Humans at 7 Tesla
Essa Yacoub, Amir Shmuel, Josef Pfeuffer, Pierre-Francois Van De Moortele,
Gregor Adriany, Peter Andersen, J. Thomas Vaughan, Hellmut Merkle, Kamil Ugurbil,
and Xiaoping Hu
*
This article describes experimental studies performed to dem-onstrate the feasibility of BOLD fMRI using echo-planar imaging (EPI) at 7 T and to characterize the BOLD response in humans at this ultrahigh magnetic field. Visual stimulation studies were performed in normal subjects using high-resolution multishot EPI sequences. Changes in R*2arising from visual stimulation were experimentally determined using fMRI measurements ob-tained at multiple echo times. The results obob-tained at 7 T were compared to those at 4 T. Experimental data indicate that fMRI can be reliably performed at 7 T and that at this field strength both the sensitivity and spatial specificity of the BOLD response are increased. This study suggests that ultrahigh field MR systems are advantageous for functional mapping in humans. Magn Reson Med 45:588 –594, 2001.©2001 Wiley-Liss, Inc.
Key words: cerebral function; functional imaging; MRI; fMRI; BOLD; ultrahigh field
Since its initial demonstration in 1992 (1–3), functional magnetic resonance imaging (fMRI) has evolved into a widely used methodology for mapping neuronal function. The basis of most fMRI studies is the blood oxygenation level dependent (BOLD) contrast (4 – 6), which is derived from the fact that the magnetic property of hemoglobin depends on its oxygenation state (7). Because deoxygen-ated hemoglobin is paramagnetic, deoxygendeoxygen-ated blood leads to a local decrease in T*2 and T2. Neural activity causes a local increase in blood flow, which overcompen-sates any increase in oxygenation utilization (8) and leads to a reduction in local deoxyhemoglobin concentration. Consequently, neural activity leads to an elevation inT2 -orT*2-weighted images. Based on this principle, the ma-jority of fMRI experiments are performed by acquiring a series ofT*2-weighted images while a subject performs a certain task or experiences some type of stimulation.
As a susceptibility phenomenon, the BOLD contrast can be imaged with MRI at various magnetic field strengths. The development of fMRI methodology coincided with the introduction of high magnetic fields, and one of the sem-inal studies describing fMRI in the human brain for the first time was performed at 4 T (1). However, due to their wide availability, 1.5 T systems are currently used for the majority of fMRI studies in humans. In animal models, higher magnetic fields (4.7 T and 9.4 T) have been widely used (9 –11). Theoretical considerations (12–14) and
ex-perimental data (10,11,15,16) have consistently revealed that the specificity, sensitivity, and contrast of the BOLD response to neural activity increases with the field strength (also see 17–21). This field strength dependence has led to a recent surge in the number of high field (3 and 4 T) MRI systems available. Recently, two human systems operating at fields higher than 4 T became available. The 8 T system has generated a great deal of interesting data (22), mostly in anatomic imaging. With the 7 T whole-body system in our laboratory, we have begun experimental fMRI studies in humans for the first time at magnetic field strengths that significantly exceed 4 T. The experiments were conducted using a surface coil with a visual stimulation paradigm. Functional mapping of the visual cortex using echo-planar imaging (EPI) at 7 T was demonstrated and activation-related R*2 changes (⌬R*2) were quantified. This article presents the experimental details and results.
MATERIALS AND METHODS
Data Acquisition
Normal subjects were enrolled for this study, which was approved by the institutional review board at the Univer-sity of Minnesota. All subjects provided written consent. Functional imaging studies were performed using a visual stimulation paradigm where the stimulus was generated by red LED goggles flashing at 10 Hz (Grass Instruments, Quincy, MA). A block design was used for all studies.
A 90-cm bore 7 T magnet (Magnex Scientific, Abingdon, UK) coupled with a Varian INOVA console (Palo Alto, CA) was used. For all studies, a quadrature transmit and re-ceive surface coil, designed for covering the occipital cor-tex (23), was used. Scout images were first obtained with an inversion-recovery (IR) prepared TurboFLASH (24) se-quence (TI: 1.4 s, TR/TE: 8/3 ms, matrix: 128⫻128, slice thickness: 5 mm) for identifying the slice of interest. A sagittal slice, 6 mm from the midline, was selected. An IR-prepared segmented TurboFLASH (TI: 1.4 s, TR/TE: 8/3 ms, two segments, two averages, matrix: 256⫻256, slice thickness: 5 mm) image was obtained over the selected slice as an anatomic reference. Functional imaging studies were performed using a T*2-weighted EPI sequence. The EPI sequence was implemented with a readout gradient consisting of alternating trapezoidal gradient lobes. For Nyquist ghost removal, a reference scan in the fMRI series was obtained and used to correct for discrepancy between the odd and even echoes (25). Note that the algorithm as described by Bruder et al. (25) also removes to a large extent off-resonance induced distortions in the EPI images, providing a reasonable registration between the fMRI data and the anatomic scan.
fMRI experiments were performed using a segmented T*2-weighted EPI sequence (matrix: 256⫻256 matrix, eight
Center for Magnetic Resonance Research and Department of Radiology, University of Minnesota, Minneapolis, Minnesota.
Grant sponsor: National Institutes of Health; Grant numbers: P41RR08079 (a National Centers for Research Resources (NCRR) grant); RO1MH55346; Grant sponsors: W.M. Keck Foundation; National Foundation for Brain Imag-ing (NFFBI).
*Correspondence to: Xiaoping Hu, Ph.D., CMRR, 2021 6th Street SE, Minne-apolis, MN 55455. E-mail: [email protected]
Received 12 June 2000; revised 19 September 2000; accepted 16 October 2000.
segments, 20⫻20 cm2 FOV, 5 mm slice, half-Fourier ac-quisition with eight additional lines). Due to the relatively short T*2 at 7 T, k-space segmentation was needed for high-resolution EPI. The EPI images were acquired with a TR of 0.37 s per segment (corresponding to an acquisition time per image of 3 s) and an average flip angle of 40° (averaged over the Calcarine sulcus). During the acquisi-tion of fMRI data, the subject’s respiraacquisi-tion and heartbeat were monitored for subsequent removal of physiological noise. The study was performed in five normal volunteers. The fMRI protocol consisted of acquiring 81 images con-secutively, during which the stimulus was turned off for nine images and on for nine images in an alternating manner. The same protocol was run for TEs of 10, 15, 22.5, 34, and 51 msec. A study was performed in the same group of subjects at 4 T using the same stimulation protocol. The images at 4 T were acquired on the same sagittal slice using identical parameters except the TEs and a surface coil based on an equivalent design. The TEs used at 4 T were 15, 22.5, 34, 51, and 76 msec. At both fields, field homogeneity was optimized with shimming, achieving a typical slice linewidth of 15 Hz at 4 T and 20 Hz at 7 T.
Data Processing
The acquired k-space data of the T*2-weighted EPI time series were preprocessed using a retrospective technique (26) to remove physiological fluctuations before the appli-cation of a Fourier transform to convert the raw data into images. The first seven images in each time series were discarded to avoid intensity variations during the transi-tion to steady state. The four epochs in the fMRI time series were averaged to generate a single epoch for further analysis.
TheT*2-weighted EPI images obtained at different TEs were first analyzed separately using cross-correlation (27) using a correlation template generated by convolving the boxcar function of the stimulation paradigm with a hemo-dynamic response described by Friston et al. (28). Cross-correlation maps were thresholded with a single threshold (0.7;P⬍0.001) to generate maps at corresponding TEs. In the second part of the data analysis, regions of interest (ROIs) over gray matter and draining veins, detectable at the image resolution employed, were identified. Veins were recognized from the anatomic image and the T*2 -weighted images in which they appeared dark (5,15). The ROIs for the two field strengths were matched by selecting the same sagittal slice for the studies at both fields and using anatomic landmarks for the location of the ROIs (see Fig. 2a). Within each ROI, the average intensity for the activation period, defined as the average of images 4 –9 in the nine-image stimulation period (the first three images skipped to account for hemodynamic response), and that for the resting period, defined to be the average of the three images prior to stimulus onset, were calculated. TheT*2for the resting and active conditions were derived using ex-ponential fitting of the average image intensity vs. TE. The ⌬R*2 of these ROIs was calculated for the two field strengths and all subjects.
T2Measurements of Human Venous Blood
Venous blood was drawn from volunteers from the arm into a standard, evacuated, 10 cc vial that contained an appropriate amount of solid heparin to prevent clotting. The tube was quickly wrapped to insulate it and inserted into a specially constructed RF coil.T2values were mea-sured with a spin echo sequence with different TEs. Each measurement was performed within a minute and the first measurement was performed no later than a minute or two after the blood was drawn. The sample tube was then removed from the coil, shaken, reinserted, and the mea-surement repeated. Three or four such meamea-surements were performed with highly reproducible T2 values in each case. The reported data for each volunteer is the average of these measurements. Blood O2saturation was measured with a blood gas analyzer before and after theT2 measure-ments and found to be the same.
RESULTS
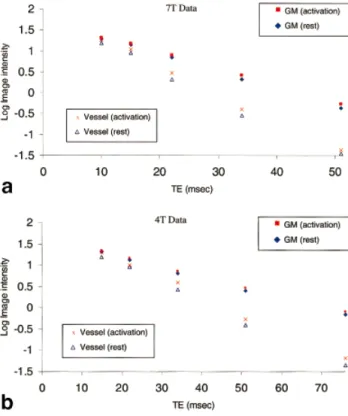
Figure 1a displays activation maps obtained from one sub-ject at all TEs for both 7 and 4 T. Similar maps were generated from data of the other subjects. At the same statistical threshold, the 7 T maps exhibited a considerably larger activated area at all TEs except the last one, partic-ularly in the gray matter regions. A paired t-test for the number of activated pixels (averaged over all TEs) between the 7 T and 4 T data showed that the 7 T maps were significantly (P⬍0.008) larger than those of the 4 T data. To illustrate the quality of the functional data, the time course of the activated regions are shown for TE of 22 msec at 7 T and compared with that of TE of 30 msec at 4 T in Fig. 1b. Activation correlated changes exhibit a higher CNR at 7 T. As expected, the activation maps in Fig. 1a varied with TE and became most prominent at a certain TE, depending on the field strength. This is more clearly demonstrated in Fig. 2b, which plots the signal change⌬S (average signal during activation – average signal during rest) as a function of TE for gray matter and vessel ROIs indicated in Fig. 2a. In the gray matter,⌬S had a maximum at a TE of 25 msec at 7 T, and 35 msec at 4 T, consistent with the theoretical prediction that there is a maximum for activation-induced signal change which occurs when TE⫽T*2(29). At 4 T,⌬S in regions associated with vessels showed an approximate plateau that persisted until a TE of 35 msec. In the ROI surrounding large vessels, a mono-tonic decrease was seen with the TE values of 10 msec or more employed in the 7 T experiments; presumably, this is because the venous blood T*2 is 10 msec or less at 7 T, consistent with the rapid decrease in venous bloodT2and T*2with increasing magnetic field (11). The shortened ve-nous bloodT*2at the higher magnetic field is also evident in theT*2-weighted images (Fig. 2a); the large venous ves-sel contributions (see arrow in Fig. 1a) disappeared in the 7 T activation maps at long TEs, while the same vascular contributions remained prominent with increasing TE at 4 T.
Figure 3 plots the logarithm of the signal with respect to TE during rest and activation, respectively. Although the signals mostly follow an exponential decay, this
exponential behavior is not so well maintained by the signals from large blood vessel regions. This is most likely due to the fact that there is more signal fluctuation in the vascular area from cardiac pulsation. In addition, pixels that contain both tissue and relatively large ve-nous blood volume fraction (e.g., due to the presence of large blood vessels) may also exhibit an oscillatory be-havior (17–20). This oscillatory bebe-havior arises because the deoxyhemoglobin containing blood has a different resonance frequency compared to surrounding tissue (17–20).
TheT*2during the rest condition for the ROIs considered were as follows. Averaged over all subjects and all resting period images, the gray matterT*2was 25.1⫾3.5 msec at 7 T and 41.4⫾5.5 msec at 4 T. Note that this measurement should be minimally affected by the shimming because they are voxel-specific and the linewidth of each voxel should be negligible compared to theT*2. These measured values are in good agreement with the optimum TEs cor-responding to the largest⌬S. In the vessel regions, theT*2
was found to be 12.8⫾0.9 msec at 7 T and 23.5⫾2.7 msec at 4 T. However, these values can be regarded as approx-imations because of complications associated with partial volume effects with surrounding tissue. A separate mea-surement of ex vivo venous blood found the T2 to be approximately 7 msec for blood with a normal O2 satura-tion (Y) of 60% (subject 1:T2⫽6.8⫾0.4 msec, Y⫽38%; subject 2:T2⫽7.1⫾0.7 msec, Y⫽39%; subject 3:T2⫽ 13.1⫾0.2 msec, Y⫽59%).
Table 1 lists the⌬R*2for ROIs in gray matter and venous vessel regions for each subject at both field strengths. For gray matter, the average⌬R*2was found to be 1.5⫾0.3 s⫺1 at 7 T and 0.68⫾ 0.09 s⫺1at 4 T. The ratio between the ⌬R*2changes at the two fields is 2.1 ⫾0.2, which repre-sents a supralinear increase with the field strength. The vessel ROI (Table 1) at 4 T shows a larger intersubject variation than the gray matter ROI, presumably due to more contributions from the vascular signal at 4 T. In the vessel ROI, the ⌬R*2 change increased only slightly with the field strength.
FIG. 1. a:Activation maps obtained at different TEs in one subject at 4 T (top) and 7 T (bottom). TEs in msec are marked in the maps.b: Timecourses for TE of 22 msec at 7 T (left) and TE of 34 msec at 4 T (right).
DISCUSSION
Functional mapping in the visual cortex in humans at 7 T is demonstrated in this work. Experimental results indi-cate that fMRI can be robustly performed at 7 T using ultrafast imaging techniques such as EPI. The study based on multiple TEs reveals that at 7 T, in the human visual cortex,T*2is 25 msec and that this is also the optimal TE to be used in BOLD based fMRI studies.
Despite the concerns with the high magnetic field, the only side effect reported by the subjects was a slight diz-ziness if they were pushed into the magnet too fast. This effect is similar to what has been reported at 4 T. Prior to the present fMRI study, we have conducted a behavior study at 7 T using a mental rotation paradigm that was previously employed to evaluate the effects of 4 T (30). The behavior data (to be reported separately elsewhere) showed that there was no difference in subject perfor-mance in and outside the magnet.
The studies presented here demonstrate that blood-related mechanisms that contribute to the BOLD effect (see discussion in 17–21) are virtually inoperative at 7 T for TEs equal to or exceeding the optimum TE of 25 msec, while they are still significant at the optimum TE of 35 msec at 4 T (see Fig. 2, 34 msec and 22 msec images for 4T and 7T, respectively). This has significant implications with respect to the specificity of the func-tional images at 7 T since such blood-related effects are mostly associated with large blood vessels. Previous studies (13,17–21,31,32) based on eliminating blood contributions to BOLD mechanism using Stejskal-Tan-FIG. 2. a:ROIs for data (4 T: top; 7 T: bottom) shown in Figs. 3b, 2b,
and 4. The gray matter ROI is shown in blue and vessel regions are indicated in green.b:Plots of⌬S vs. TE for gray matter ROI (top) and vessel ROI (bottom).
FIG. 3. a:Plot of signal intensity vs. TE for gray matter ROI and vessel ROI at 7 T.b:Plot of signal intensity vs. TE for gray matter ROI and vessel ROI at 4 T.
ner gradients (33) have demonstrated that most if not all of the signal changes induced by neuronal activation originate from intravascular or blood-related effects at 1.5 T. The same conclusion was shown to be applicable, albeit to a lesser extent, even to 4 T at TE values of 20 –30 msec that are often used at this field strength (17–20,34). At these field strengths, the fractional BOLD signal changes induced by activation substantially ex-ceed the ⬃2% microvascular blood volume fraction in the brain (35). Therefore, these blood-related effects are thought to arise from blood within the large vessels (17) (i.e., with diameters comparable or larger than voxel dimensions) or voxels that are occupied by both tissue and venous blood with comparable partial volumes (i.e., voxels partially occupied by relatively large blood ves-sels). The partial volume effect in the latter case can lead to large signal modulations with activation due to the susceptibility-induced difference in the resonance fre-quencies of the two compartments (17–20). We have previously argued that these mechanisms should rap-idly become inoperative at magnetic fields above 4 T (17–20) due to the rapidly decreasingT2of blood with increasing magnetic field (11) and we have experimen-tally demonstrated the absence of these mechanisms at 9.4 T (11). Similar results are also expected at 7 T in awake human subjects. While the field strength is some-what lower relative to the animal studies, fractional oxyhemoglobin content in venous blood is also signifi-cantly higher in awake humans relative to anesthetized animals (60% vs.⬃80% in the animal model in the 9.4 T study (11)). Consistent with these expectations, bloodT2 at 7 T was found to be approximately 7 msec for 60% oxygenation level. This value is in remarkable agree-ment with the value of 6.7 msec predicted from the model based on 1.5 T data on human blood T2(36) if a quadratic dependence on magnetic field is assumed.
In the absence of the above-described BOLD mecha-nism related to blood effects, macroscopic venous blood vessels can only contribute toT*2-weighted images at 7 T through the extravascular BOLD mechanism arising from static averaging of field gradients around these vessels (e.g., see discussions in 17–20,16-19). Such ef-fects, however, are expected to be small at sites distant from the activated tissue because of dilution from ves-sels draining inactive areas. Therefore, they are not ex-pected to be a major source of degradation of specificity of the fMRI maps at 7 T.
If only the extravascular BOLD effect associated with blood vessels larger than capillaries and postcapillary venules were to dominate the fMRI signals at 4 and 7 T, then at best only a linear increase in activation related ⌬R*2is expected. If we consider the blood-related contri-butions toT*2-weighted fMRI, then the linear dependence becomes an upper limit in the 4 T to 7 T comparison. Therefore, the supralinear increase observed in ⌬R*2 in going from 4 to 7 T demonstrates the presence of a signif-icant microvascular contribution which is expected to contribute supralinearly. Note that, although the anatomic landmarks used in ROI placement were clearly identifiable on both the 4T and 7T images, and the slices appear to be very similar in location, there may be some error in regis-tering the ROIs across the different scans in an attempt to assess the field strength dependence.
Increased microvascular contribution and diminishing large vessel contribution to fMRI leads to increased spec-ificity at the higher magnetic field of 7 T. It is possible that basal signal fluctuations increase at 7 T. This is currently being investigated separately. However, even if enhanced, such fluctuations will not necessarily degrade fMRI stud-ies at 7 T because they can be eliminated by a variety of methods (26,37,38). In fact, in this study, where the fluc-tuations associated with respiration and cardiac pulsation were removed, the temporal stability of the fMRI time series was found to be comparable at the two field strengths and a clear gain in contrast-to-noise ratio for fMRI was demonstrated in going from 4 to 7 T. This point is also illustrated by the timecourses shown in Fig. 1b. In addition, MR SNR also increases with field strength, as systematically demonstrated in a recent volume coil study (39), providing further gains for fMRI studies. Therefore, we conclude that major advantages are realized in fMRI studies of brain function at 7 T.
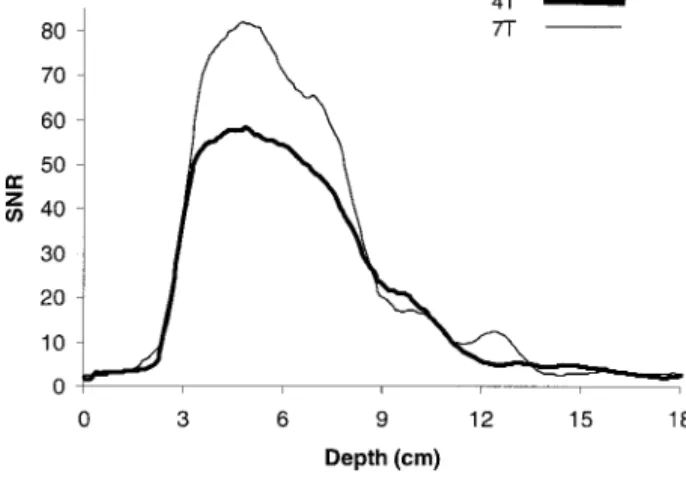
A potential problem at 7 T is theB1inhomogeneity and its potential degradation on SNR. While a full investiga-tion of this issue is outside the scope of this article, we examined the SNR in the acquired images to get an idea on this issue. The SNR profiles in the raw images along a line perpendicular to the surface coil were obtained by smooth-ing the image profile and dividsmooth-ing by the noise estimated from a region outside the brain. The profiles for the two field strengths are plotted in Fig. 4 for one of the subjects. Evidently, the SNR at 7 T is higher for most of the ROI but falls off more quickly, probably due to increased dielectric Table 1
R*2Change Arising From Visual Stimulation
Subject
Gray matter⌬R*2(s⫺1) Vessel region⌬R*2(s⫺1)
4 T⌬R*2 (s⫺1) 7 T⌬R*2 (s⫺1) Ratio 4 T⌬R*2 (s⫺1) 7 T⌬R*2 (s⫺1) Ratio kb 0.69 1.5 2.20 1.5 1.9 1.27 sc 0.68 1.4 1.99 1.1 1.3 1.18 pa 0.58 1.0 1.80 1.6 1.7 1.06 rg 0.62 1.4 2.30 1.2 1.3 1.22 dc 0.82 1.9 2.34 0.9 1.1 1.16 Mean⫾STD 0.68⫾0.09 1.46⫾0.32 2.13⫾0.23 1.26⫾0.29 1.46⫾0.33 1.18⫾0.09 The negative sign in front of the⌬R*2is ignored for clarity.
resonance. Thus, the effect of dielectric resonance in the present study is not substantial.
ACKNOWLEDGMENTS
The authors thank Mr. Sang-Pil Lee for assistance with the T2measurement of the venous blood.
REFERENCES
1. Ogawa S, Tank DW, Menon R, Ellermann JM, Kim S-G, Merkle H, Ugurbil K. Intrinsic signal changes accompanying sensory stimulation: functional brain mapping with magnetic resonance imaging. Proc Natl Acad Sci USA 1992;89:5951–5955.
2. Kwong KK, Belliveau JW, Chesler DA, Goldberg IE, Weisskoff RM, Poncelet BP, Kennedy DN, Hoppel BE, Cohen MS, Turner R, Cheng H-M, Brady TJ, Rosen BR. Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc Natl Acad Sci USA 1992;89:5675–5679.
3. Bandettini PA, Wong EC, Hinks RS, Tikofsky RS, Hyde JS. Time course EPI of human brain function during task activation. Magn Reson Med 1992;25:390 –398.
4. Ogawa S, Lee T-M, Kay AR, Tank DW. Brain magnetic resonance imaging with contrast dependent on blood oxygenation. Proc Natl Acad Sci USA 1990;87:9868 –9872.
5. Ogawa S, Lee T-M. Magnetic resonance imaging of blood vessels at high fields: in vivo and in vitro measurements and image simulation. Magn Reson Med 1990;16:9 –18.
6. Ogawa S, Lee T-M, Nayak AS, Glynn P. Oxygenation-sensitive contrast in magnetic resonance image of rodent brain at high magnetic fields. Magn Reson Med 1990;14:68 –78.
7. Pauling L, Coryell CD. The magnetic properties and structure of hemo-globin, oxyhemoglobin and carbonmonoxyhemoglobin. Proc Natl Acad Sci USA 1936;22:210 –216.
8. Fox PT, Raichle ME. Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc Natl Acad Sci USA 1986;83:1140 –1144. 9. Silva A, Lee SP, Yang C, Iadecola C, Kim S. Simultaneous BOLD and
perfusion functional MRI during forepaw stimulation in rats. J Cereb Blood Flow Metab 1999;19:871– 879.
10. Duong TQ, Silva AC, Lee SP, Kim SG. Functional MRI of calcium-dependent synaptic activity: cross correlation with CBF and BOLD measurements. Magn Reson Med 2000;43:383–392.
11. Lee SP, Silva AC, Ugurbil K, Kim SG. Diffusion-weighted spin-echo fMRI at 9.4 T: microvascular/tissue contribution to BOLD signal changes. Magn Reson Med 1999;42:919 –928.
12. Ogawa S, Menon RS, Tank DW, Kim S-G, Merkle H, Ellermann JM, Ugurbil K. Functional brain mapping by blood oxygenation level-de-pendent contrast magnetic resonance imaging. Biophys J 1993;64:800 – 812.
13. Boxerman JL, Bandettini PA, Kwong KK, Baker JR, Davis TJ, Rosen BR, Weisskoff RM. The intravascular contribution to fMRI signal changes: Monte Carlo modeling and diffusion-weighted studies in vivo. Magn Reson Med 1995;34:4 –10.
14. Kennan RP, Zhong J, Gore JC. Intravascular susceptibility contrast mechanisms in tissues. Magn Reson Med 1994;31:9 –21.
15. Menon RS, Ogawa S, Tank DW, Ugurbil K. 4 Tesla gradient recalled echo characteristics of photic stimulation-induced signal changes in the human primary visual cortex. Magn Reson Med 1993;30:380 – 386.
16. Gati JS, Menon RS, Ugurbil K, Rutt BK. Experimental determination of the BOLD field strength dependence in vessels and tissue. Magn Reson Med 1997;38:296 –302.
17. Ugurbil K, Ogawa S, Kim S-G, Hu X, Chen W, Zhu XH. Imaging brain activity using nuclear spins. In: Maraviglia B, editor. Magnetic reso-nance and brain function: approaches from physics. Amsterdam: Ital-ian Physical Society Press; 1999. p 261–310.
18. Ugurbil K, Hu X, Chen W, Zhu XH, Kim SG, Georgopoulos A. Func-tional mapping in the human brain using high magnetic fields. Philos Trans Royal Soc Lond B 1999;354:1195–1213.
19. Ugurbil K, Chen W, Hu X, Kim S-G, Ogawa S, Zhu XH. Functional MRI at high fields: practice and utility. In: Young I, editor. Magnetic reso-nance imaging and spectroscopy in medicine and biology. Chichester, UK: John Wiley & Sons; 2000. p 603– 623.
20. Ugurbil K, Adriany G, Andersen P, Chen W, Gruetter R, Hu X, Merkle H, Kim DS, Kim SG, Strupp JP, Zhu XH, Ogawa S. Magnetic resonance studies of brain function and neurochemistry. Annu Rev Biomed Eng 2000;2:633– 660.
21. Ogawa S, Menon RS, Kim SG, Ugurbil K. On the characteristics of functional magnetic resonance imaging of the brain. Annu Rev Biophys Biomol Struct 1998;27:447– 474.
22. Robitaille PM, Abduljalil AM, Kangarlu A, Zhang X, Yu Y, Burgess R, Bair S, Noa P, Yang L, Zhu H, Palmer B, Jiang Z, Chakeres DM, Spigos D. Human magnetic resonance imaging at 8 T. NMR Biomed 1999;12: 315–319.
23. Adriany G, Yacoub E, Tkac I, Andersen P, Merkle H, Vaughan JT, Ugurbil K. 7T vs. 4T: preliminary B1, SNR, SAR comparison in the human head. In: Proc 8th Annual Meeting ISMRM, Denver, CO, 2000.
24. Haase A. Snapshot FLASH MRI: application to T1, T2, and chemical shift imaging. Magn Reson Med 1990;13:77– 89.
25. Bruder H, Fisher H, Reinfelder HE, Schmitt F. Image reconstruction for echo planar imaging with nonequidistant k-space sampling. Magn Re-son Med 1992;23:311–323.
26. Hu X, Le TH, Parrish T, Erhard P. Retrospective estimation and correc-tion of physiological fluctuacorrec-tion in funccorrec-tional MRI. Magn Reson Med 1995;34:201–212.
27. Bandettini PA, Jesmanowicz A, Wong EC, Hyde JS. Processing strate-gies for time-course data sets in functional MRI of the human brain. Magn Reson Med 1993;30:161–173.
28. Friston KJ, Jezzard P, Turner R. Analysis of functional MRI time-series. Hum Brain Map 1994;1:153–171.
29. Ugurbil K, Garwood M, Hendrich K, Hinke R, Hu X, Menon RS, Merkle H, Ogawa S, Salmi R. Imaging at high magnetic fields: initial experi-ences at 4 Tesla. Magn Reson Q 1993;9:259 –277.
30. Tagaris GA, Kim S-G, Strupp JP, Ugurbil K, Georgopoulos AP. Mental rotation studied by functional magnetic resonance imaging at high field (4 Tesla): performance and cortical activation. J Cogn Neurosci 1997; 9:419 – 432.
31. Lai S, Hopkins AL, Haacke EM, Li D, Wasserman BA, Buckley P, Friedman L, Meltzer H, Hedera P, Friedland R. Identification of vascu-lar structures as a major source of signal contrast in high resolution 2D and 3D functional activation imaging of the motor cortex at 1.5 T: preliminary results. Magn Reson Med 1993;30:387–392.
32. Song AW, Wong EC, Tan SG, Hyde JS. Diffusion weighted fMRI at 1.5 T. Magn Reson Med 1996;35:155–158.
33. Stejskal EO, Tanner JE. Spin diffusion measurements: spin-echoes in the presence of a time dependent field gradient. J Chem Phys 1965;42: 288 –292.
FIG. 4. A plot of SNR vs. depth for representative EPI images in one subject at 7 T (red) and 4 T (black). The SNR was calculated by smoothing an intensity profile and dividing it with the average noise standard deviation from the image background.
34. Menon RS, Hu X, Adriany G, Andersen P, Ogawa S, Ugurbil K. Com-parison of spin-echo EPI, asymmetric spin-echo EPI and conventional EPI applied to functional neuroimaging: the effect of flow crushing gradients on the BOLD signal. In: Proc 2nd Conference SMR, San Francisco, 1994.
35. Pawlik G, Rackl A, Bing RJ. Quantitative capillary topography and blood flow in the cerebral cortex of cats: an in vivo microscopic study. Brain Res 1981;208:35–58.
36. Wright GA, Hu BS, Macovski A. Estimating oxygen saturation of blood in vivo with MR imaging at 1.5T. J Magn Reson Imag 1991;1:275–283.
37. Mitra PP, Ogawa S, Hu X, Ugurbil K. The nature of spatio-temporal changes in cerebral hemodynamics as manifested in functional mag-netic resonance imaging. Magn Reson Med 1997;37:511–518. 38. Biswal B, DeYoe EA, Jesmanowicz A, Hyde JS. Removal of
physiolog-ical fluctuations from functional MRI signals. In: Proc 2nd Conference SMR, San Franciso, 1994.
39. Vaughan JT, Garwood M, Collins CM, DelaBarre L, Adriany G, Andersen P, Merkle H, Smith MB, Ugurbil K. 7T vs. 4T: preliminary B1, SNR, SAR comparison in the human head. In: Proc 8th Annual Meeting ISMRM, Denver, CO, 2000.