GEOGRAPHICAL PATTERNS O F
FOUR
POLYMORPHISMS INZOARCES VZVZPARUS AS EVIDENCE OF SELECTION1
FREDDY BUGGE CHRISTIANSEN AND OVE FRYDENBERG
Department of Biological Sciences, Stanford University, Stanford, California 94305 and Department of Genetics, University of Aarhus, DK-8000 Aarhus C, Denmark3
Manuscript received February 12, 1974
ABSTRACT
Two unlinked polymorphisms of a marine teleost, one in hemoglobin and one in esterase, show parallel clines through Danish waters, while two un- linked phosphoglucosemutase polymorphisms maintain constant gene frequen- cies. It is argued that the clinal and the constant polymorphisms cannot be ac- counted for simultaneously by random genetic drift of selectively neutral genes. It is therefore concluded that selection is responsible for at least one of the two classes of geographical patterns, be it the clinal, the constant, or both.
A N Y geographical pattern in a polymorphism can be explained by random genetic drift if one is free to postulate a suitable population structure, i.e., to imagine enough subpopulations, passed or present, of adequate sizes and with adequate migration rates. The hypothesis that a polymorphism is selectively neutral can therefore never be rejected by an argument founded exclusively on an observed geographical pattern. Even regular gene frequency clines, so sug- gestive of selection to some observers, do not contradict a notion of neutrality (WRIGHT 1940, 1951; KIMURA and MARUYAMA 1971). So any evaluation of the selective status of a single polymorphism. based on its geographical variation, re- quires access to solid knowledge of the population structure in a n extent rarely available.
While a single polymorphism always can be explained by a suitable hypo- thetical population structure, this is not necessarily true about two or more polymorphisms, because the breeding structure, contrary to selection, affects all loci and all alleles uniformly. This property was used in 1966 by CAVALLI-SFORZA and, more recently, by LEWONTIN and KRAKAUER (1973) to appraise the selec- tive status of an assortment of polymorphisms in man. The tool employed was a simple index of population structure, the standardized gene frequency variance among subpopulations. This so-called “effective inbreeding coefficient” is expected to be the same for all polymorphisms owing their geographical patterns entirely to the breeding structure. Both investigations concluded that at least some of the surveyed polymorphisms were or had been subject to selection.
This work was supported by two research grants from the Carlsberg Foundation of Copenhagen to 0. FRYDENBERG, a travel grant from the Danish Natural Science Research Council to F. B. CHRISTIANSEN and an N.I.H. grant, GM-10452-11.
Pennanent address of both authors: Department of Genetics, Ny Munkegade, DK-8000 Aarhus C, Denmark.
7GG
F. B. CHRISTIANSEN AND 0. FRYDENBERGThis note is devoted to the interpretation of the geographical patterns of four bi-allelic polymorphisms occurring in the eelpout, Zoarces uiuiparus
L.,
a marine teleost. The eelpout is found along most of the European west coast from the Barents Sea to the British Channel. I t is specially abundant in the inner Danish waters and extends its distribution into the Gulf of Botnia in the Baltic Sea.MATERIALS
The four bi-allelic loci studied a r r a hemoglobin locus, HbI (J. P. HJORTH, in preparation) and thrcc enzyme loci, uiz. two phosphoglucoscmutase loci, Pgml and PgniII (HJORTII 1971) and an esterasc locus, E d I l (SIMONSEN and FRmeNnenc 1972). Pgml, PRmII, and EstIII are iinlinked. as are Z 3 t I I I and Hbl. Nothing is known about the linkage relation hetween the HbI
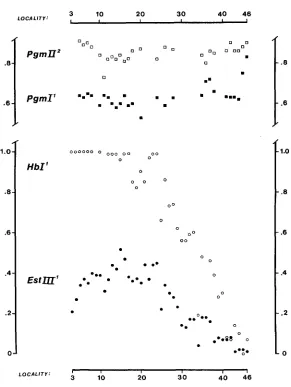
and the Pgm loci. The geographical distribution of the EstIII alleles, which was studied a t the sampling stations shown in Figure 1, has been described by F n ~ o r x w v " PI al. (1973). Geo- Rraptiical data on the other polymorphisms await publication. The gene frequencies, kindly made available for this paper by our departmental collcagues, are summarized in Figure 2. Most samples contained more than one hundred fish, but some were smaller.
BALTIC SEA
3 10 2 0 30 40 46
1 I I I I I
767 LOCALITY: PgmL12 PgmI' 1.0 .8 .6 .4 .2 0 HbI' EstILT'
L O C A L I T Y :
0
0 0
0
m .
0
0 0 0
0 0 0 0 0 0 0 0 0 0 0 e * -.e
e -.e- 0
e -
-
e 0
e 0
.
I I 1
3 10 2 0 30 40 46
.8
1
.6 1 .o.8
.6
.4
.2
0
FIGURE 2.-The gene frequencies at the various sampling stations which appear arranged in geographical order.
R E S U L T S A N D D I S C U S S I O N
768 F. B. CHRISTIANSEN AND 0. FRYDENBERG
low of some 8 percent in Holland (FRYDENBERG et al. 1973). The PgmW allele exhibits slight frequency variations in the northern region, being perhaps lower in the Kattegat than in the Lim Fjord, but the overall pattern of both Pgm
loci is one of constant gene frequencies. Two small samples from the inner Baltic, #45 and 46, indicate that the frequencies attained by all four polymorphisms at
the mouth of the Eastern Baltic, #41-44, prevail throughout the inner Baltic Sea. Our observations may be thought about in terms of a population subdivided into three parts: A northern Kattegat subpopulation characterized by high HbZ'
and EstIIZl frequencies, a southeastern Baltic subpopulation with very low gene frequencies i n the HbZ and EstIIZ polymorphisms, and a connecting Belt popula- tion featuring two parallel frequency clines. The problem posed by these data may therefore be condensed to the question of explaining the divergence between the Kattegat and the Baltic subpopulations, shown in Table 1.
TABLE 1
Gene frequencies in the northern and the southeastern subpopulation
Hi111 EssiIIl ' Pgml' P g m l P The Kattegat subpopulation 1
.oo
.40 .60 3 5The Baltic subpopulation .IO .05 .60 3 5
If
N
is the effective population size and m the migration rate per generation, two subpopulations are expected to diverge due to random genetic drift if N m e 1 and to remain alike if N m 1(WRIGHT
1940;KIMURA
and MARUYAMA 1971). This model would indeed explain the frequency pattern at each locus separately. Sufficiently large population sizes or extensive migration or a sufficiently short separation time would make the consistency 04 the Pgm loci probable. Smallpopulation sizes, very restricted migration o r long separation time would account not only for the divergence but also f o r parellel diffusion clines in the other two polymorphisms. Slight perturbations in the relation between the two clines or in the constancy of the Pgm polymorphisms, may be credited to minor local differ- entations due to a low migration.
Now, if N m has been large ever since these waters were invaded after the last Glacial Age some 10,000 years ago, genetic drift cannot be held responsible for the divergence of the clinical polymorphisms. The question is therefore whether a product, N m , small enough to allow divergence at all, could have prevailed in
a time span that would still allow many polymorphisms to maintain constant gene frequencies. If the gene frequency differences between the two subpopula- tions in Table 1 are compared to the distributions expected among isolated pop- ulations
(WRIGHT
1940; KIMURA 1955; see also Figures 8.4.2 and 9.2.1 in CROW and KIMURA 1970), it appears that the separation time must have been shorter than N/10 to make the constancy of the Pgm loci probable, whereas the diverg- ence in the two other loci requires a separation time of at least N generations. Obviously, these requirements cannot be met at the same time. Thus the drift models, which appear applicable to our observations, make either the constant or769 evoked to account for the geographical pattern of at least one of the two groups
of polymorphisms.
Since the generation time in eelpouts is 2 to 3 years, a population size of the order of 50,000 is needed to conserve supposedly neutral Pgm frequencies during ten millennia. We have released 10,000 marked fish on station #18, and the re- captures, which vanished to zero in a few months, were never more than 10 per- cent. Present populations are thus very large, and they may also in the past have been sufficient so to secure the P g m constancies. On the other hand, the diverg- ence in the clinical polymorphisms may, if these are assumed neutral, always be claimed to be due to occasional bottlenecks. From 6,000 to 4,000 B.C. the Baltic Sea was cut off from the Kattegat, forming the Ancylus Lake in which some or- dinarily marine organisms survived (MERTZ 1924;
NORDMANN
1936). The eel- pout is very tolerant of fresh water and, although there is no geological evidence of this, it might have formed a completely isolated Ancylus population.If
SO, a founder effect may be educed to justify random divergence. This is all pure spec- ulation which serves only to demonstrate that our ignorance about the real breed- ing structure throughout the last 10,000 years keeps u s from deciding which of the polymorphisms, the constant or the clinical ones, are unlikely results of ran- dom genetic drift. Evidence of a quite different nature, namely from direct measurements of fitness values, strongly suggests that natural selection operates on the EstZZZ polymorphism in present days ( CHRISTIANSEN,FRYDENBERG
and SIMONSEN 1973; CHRISTIANSEN et aZ. 1974).Finally we want to stress that the arguments presented here pertain specific- ally to the four polymorphisms discussed. W e believe it reasonable to conclude from the geographical patterns that at least two of these polymorphisms are or have been subject to selection. This notion does not imply a general rejection of the theory, sustained by KIMURA and OHTA (1971), that neutral mutations are sufficient to account for many polymorphisms in natural populations. However, unless our sample of polymorphisms is highly biased, it indicates that there is a sizeable fraction of all polymorphisms that are not selectively neutral.
LITERATURE CITED
CAVALLI-SFORZA, L. L., 1966
CHRISTIANSEN, F. B., 0. FRYDENBERG and V. SIMONSEN, 1973
Population structure and human evolution. Proc. Roy. Soc. London, Ser. B 164: 362-379.
Genetics of Zoarces populations. IV. Selection component analysis of an esterase polymorphism using population samples including mother-offspring combinations. Hereditas 73 : 291-304.
Genetics of
Zoarces populations. V I . Further evidence, based on age group samples, of a heterozygote deficit in the EstZII polymorphism. Hereditas (In press.)
An Introduction to Population Genetics Theory. Harper and
Row, New York.
Genetics of Zoarces
populations. 111. Geographic variations in the esterase polymorphism, EstIIZ. Hereditas 73:
233-238.
Genetics of Zoarces populations. I. Three loci determining the phospho- glucosemutase isoenzymes i n brain tissue. Hereditas 69 : 233-242.
CHRISTIANSEN, F. B., 0. FRYDENBERG, A. 0. GYLDENHOLM and V. SIMONSEN, 1974
CROW, J. F. and M. KIMURA, 1970
FRYDENBERG, O., A. 0. GYLDENHOLM, J. P. HJORTH and V. SIMONSEN, 1973
770
KIMURA, M., 1955 Solution of a process of random genetic drift with a continuous model. Proc. Natl. Acad. Sci. U.S. 41: 144-150.
KIMURA, M. and T. MARUYAMA, 1971 Pattern of neutral polymorphism in a geographically structured population. Genet. Res. 18: 125-131.
KIMUFLA, M. and T. OHTA, 1971 Theoretical Aspects of Populaiion Genetics. Princeton Univer- sity Press, Princeton.
LEWONTIN, R. C. and J. KRAKAUW, 1973 Distribution of gene frequency as a test of the selec- tive neutrality of polymorphisms. Genetics 74: 175-1 95.
MERTZ, E. L., 1924 Oversigt over de sen- og postglaciale Niveauforandringer i Danmark. Resume en francais: Les changements de niveau postglaciaires du Danemark. Danmarks Geologiske Underssgelser, I1 Raekke, Nr. 41.
NORDMANN, V., 1936 Menneskets Indvandring til Norden. English summary: The immigration of Man to Northern Europe. Ibid. I11 Raekke, Nr. 27.
SIMONSEN, V. and 0. FRYDENBERG, 1972 Genetics of Zoarces populations. 11. Three loci deter- mining esterase isozymes in eye and brain tissue. Hereditas 70: 235-242.
WRIGHT, S., 1940 Breeding structure of populations in relation to speciation. Am. Naturalist
74: 232-248.
-,
1951 The genetical structure of populations. Ann. Eugenics 15:323-354.
Coresponding editor: T. PROUT