Copyright 0 1986 by the Genetics Society of America
THE CIS-TRANS TEST SHOWS NO EVIDENCE FOR A
FUNCTIONAL RELATIONSHIP BETWEEN TWO MOUSE
TCOMPLEX LETHAL MUTATIONS: IMPLICATIONS
FOR
THE EVOLUTION OF
THAPLOTYPES
PAUL E. MAINS'
Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1 1 724
Manuscript received February 26, 1986 Revised copy accepted August 18, 1986
ABSTRACT
Mouse t haplotypes often carry embryonic lethal mutations. Sixteen comple- mentation groups are known, but the viability of the heterozygotes between them is often less than 100%. It has been reported that cis heterozygotes of two lethal mutations showed better viability than trans heterozygotes. This could indicate that the mutations were part of the same functional unit, even though they map up to 15 cM apart. However, the tW5 and tu'' haplotypes in our colony did not show a statistically significant decrease in viability when combined in
trans. The cis-trans analysis was repeated using two independent chromosomes, derived by recombination between the tW5 and the tw" haplotypes to provide the two lethal mutations in cis. Two independent chromosomes, representing the reciprocal recombination event, supplied the corresponding wild-type alleles in
cis. These chromosomes were combined in the four pairwise combinations, and male/female reciprocal crosses were done. The cis heterozygotes showed a de- crease, rather than an increase, in viability in seven of the eight cases. These results probably reflect effects of unrelated background genes. The lethal mu- tations, instead of being functionally related, may have occurred in a random, unrelated set of genes and may confer a selective advantage to t haplotypes found in wild populations.
OUSE t haplotypes represent a variant region of the proximal one-third
M
of chromosome 1 7 that shows numerous effects on fertility and devel- opment (BENNETT 1975; KLEIN and HAMMERBERG 1977; SHERMAN and WUDL1977; LYON 1981; FRISCHAUF 1985). t haplotypes contain multiple chromo- somal rearrangements which result in recombination suppression throughout a 10- to 20-cM region when heterozygous with wild type (ARTZT, SHIN and
BENNETT 1982; PLA and CONDAMINE 1984; HERRMANN et al. 1986). Males heterozygous for a t haplotype and a wild-type chromosome show transmission ratio distortion, transmitting the t haplotype to >95% of their offspring. Males who carry two different t haplotypes are sterile. However, such females are fertile, and the level of recombination between the two t haplotypes occurs at
'
Present address: Department of Molecular, Cellular, and Developmental Biology, University of Colorado, Boulder, Colorado 80309.1226 P. E. MAINS
normal levels. t haplotypes are easily identified since they share an allele, t-int, which interacts with the dominant short-tail mutation brachyury ( T ) resulting in tailless T/t heterozygotes (BODE 1984). T mutations are found within the homologous region of the wild-type chromosome 17.
There are a total of 16 embryonic lethal complementation groups known to be associated with different t haplotypes, although not all t haplotypes carry a lethal allele (KLEIN, SIPOS and FIGUEROA 1984). It has been proposed that the region of the chromosome included in t haplotypes contains a set of function- ally related loci which control development (GLUECKSOHN-WAELSCH and ER-
ICKSON 1970; BENNETT 1975). In spite of this association of t haplotypes with lethal alleles, 20-40% of wild mice carry t-bearing chromosomes.
Recently ARTZT, MCCORMICK and BENNETT (1982); ARTZT (1984) and PLA and CONDAMINE (1984) used the normal level of recombination that occurs in females heterozygous for complementing t haplotypes to map many of the lethal mutations. They showed that mutations belonging to different comple- mentation groups mapped to independent locations distributed throughout the 15- to 20-cM region. Formally, mutations belonging to different complernen- tation groups were thought to be alleles of a single complex locus (hence the terms “T locus” and “t alleles”); clearly they represent different genes.
Even though different lethal mutations are at independent loci, it is possible that they are functionally related. T h e viability of mice heterozygous for com- plementing t haplotypes generally ranges from 1 0- 100% (BENNETT 1975). This sometimes incomplete complementation might represent an additive semidom- inance of each lethal, the effects of unrelated background genes, or a direct interaction between the different lethal mutations. T o see if the different lethal loci behaved as part of a functional unit, SHIN et al. (1983b) performed a cis- trans test. Two lethal mutations were placed together by recombination on one chromosome (in cis) rather than being on separate chromosomes (in trans). They found some cases where the complementation between two different lethal mutations was increased from about 75% in trans to 100% if they were in cis and heterozygous with a t-chromosome from which the specific lethals had been removed by recombination. This cis-trans effect was interpreted as indicating that different t lethal mutations were part of a single functional unit, even though they map up to 15 cM apart. T h e authors suggested that the necessity for the functional alleles of widely separated genes to be on the same chromosome for efficient function may be indicative of genomic rearrange- ment during development.
CIS-TRANS TEST OF THE T COMPLEX
1227
I have repeated the cis-trans test using two pairs of independent recombinant chromosomes in the four possible pairwise combinations. T h e different com- binations are likely to share different chromosomal regions of overlap derived from one or the other parental haplotypes. In most cases, the level of comple- mentation was slightly decreased, rather than increased, in the cis heterozygote. T h e differences in viability are likely to have resulted from differences in genetic background. In any case, the differences in complementation for the cis heterozygote relative to the trans are minor compared to the viabilities of either homozygous mutation, which are
<
1 O-4. Therefore, the cis-trans test provides no evidence for a functional relatedness of different t mutations.Nevertheless, it is surprising that 20-40% of wild mice carry so many dif- ferent deleterious mutations in this small region of the genome. T h e origin of
t lethal alleles is discussed in terms of the evolution and the population dynam- ics of the t haplotypes.
EXPERIMENTAL PROCEDURES
Mice: The t W 5 and tW" chromosomes were originally from J.-L. GUENET (Institute Pasteur, Paris) as an N16 congenic, inbred on the 129/Sv background. These were outcrossed to a Ttfl+tf stock, obtained from S. WAELSCH (Albert Einstein, New York), and have been maintained as outbred stocks. The TtTU2tfpartial t haplotype (STYRNA
and KLEIN 1981) was provided by J. KLEIN (Tubingen, Federal Republic of Germany).
The recombinant chromosomes used in this analysis are described by MAINS (1986). All mice were maintained in our colony at Cold Spring Harbor, New York. In general, one male and two females were placed together in each mating cage.
RESULTS
Isolation of reciprocal crossover products: Recombinant t chromosomes carrying either two or no lethal mutations were first isolated. T h e t haplotypes tu'' and tW5 carry complementing lethal mutations. T h e tw5 lethal mutation maps near, or within, the major histocompatibility complex of the mouse, H- 2 , whereas the lethal mutation carried by the twl' haplotype is closely linked to the coat marker tufted
(tf)
(Figure 1; ARTZT, MCCORMICK and BENNETT1982; ARTZT 1984; PLA and CONDAMINE 1984). T h e partial t haplotype de- rived from the twl' chromosome, TtTu2tf, was used as the source of the twl'
lethal mutation (STYRNA and KLEIN 1981). T h e distal region of this chromo- some carries the twl' lethal allele, the H - 2 of twl', and a tfallele. T h e proximal region is derived from a non-t chromosome and carries a T marker (Figure 1). This partial haplotype does not show transmission ratio distortion.
Recombinants between H-2 and
tf
were isolated from TtTU2tf/tw5 females (Figure 1). Alleles of H - 2 were scored by DNA probes which detect restriction fragment length polymorphisms between the parental haplotypes. Included in the analysis was a probe for the H-2K subregion (WINOTO, STEINMETZ andHOOD 1983), which is likely to mark the right end of the H - 2 complex that is inverted with respect t o wild type in t haplotypes (SHIN et al. 1983a; SHIN, BENNETT and ARTZT 1984). This analysis is described in detail elsewhere
(MAINS 1986).
1228
f W 5
Tf ru2f f
P. E. MAINS
f -inf ~ - 2 W 5 f W 5 + t
I
T H-ZW'2
+
tf tw'2t -int ~ - 2 ~ 5 y 5 tf fWI2
...
-
r
H-ZW"'+
+ +
...
-
FIGURE 1 .-Recombinant chromosomes used for the cis-trans analysis. The parental chromo- somes are shown at the top. Females heterozygous for these two chromosomes were mated to
+tf/
+tf
males, and the recombinants t"""', tCsh"", Tt"h"O' and TtcSuo7 were selected as described in MAINS (1986). The H - 2 type (denoted H-2"' and H-2'"") and lethal factors (denoted as t"' andt"'") are indicated. The relative orders between the tw5 lethal mutation and H-2 and between the
t"" lethal mutation and t f a r e unknown. The portion of each recombinant chromosome derived from each parental haplotype is as shown, with ... representing 2n undefined region, and
-
representing material derived from non-t chromosomes. The centromere is to the left.have the centromere proximal region and H-2 of the tW5 haplotype parent, and the distal tfmarker of TtTU2$ As expected, progeny testing showed that these chromosomes carry both lethal mutations. Both show >97% transmission through males as expected for complete t haplotypes (data not shown). T h e recombinant chromosomes TtcShzo5 and TtCshzo7 are the reciprocal product in that they retain the proximal region and H-2 characteristic of TtTu2tf, and the distal +fallele of tw5. These chromosomes were found by progeny testing to carry neither lethal mutation. TtcshZo5 and TtcshZo7 show 45.4% ( N =
152)
and49.4% ( N =
237)
transmission through males, respectively, neither of which is significantly different from 50%. The designation tcsh"' will be used to non- specifically denote either tcshlo' or tcShlo2 , and TtcshZo0 will denote TtcshZ0' orExperimental design: Two recessive mutations are considered to be part of the same functional unit if an animal carrying both mutations shows a wild- type phenotype when the mutations are in cis and heterozygous with a wild- type chromosome, but displays the mutant phenotype when the mutations are in trans (LEWIS 1951; BENZER 1959). Thus, the cis-trans test compares the phenotype of the trans heterozygote to the phenotypes of the mutant and wild- type controls. T h e mutant control is the phenotype ( i . e . , viability in this case) of each homozygous mutant alone. In our case, the tw5 and twI2 mutations appear to be absolute lethals (see DISCUSSION). T h e wild-type control combines one chromosome carrying both mutations in cis with a second chromosome that carries the cis combination of the corresponding wild-type alleles at each locus. This will control for any semidominance displayed by combining the two recessive mutations in one individual.
CIS-TRANS TEST OF THE T COMPLEX 1229
With the t complex, two wild-type controls are necessary. One is to use a chromosome 17 from non-t-bearing mice to provide the wild-type alleles in cis. A second “wild-type” control is to provide wild-type function derived from t chromosomes. This is necessary because t chromosomes are known to carry deleterious mutations other than their specific lethal alleles (ARTZT, BABIARZ and BENNETT 1979; BABIARZ, GARRISI and BENNETT 1982; SHIN et al. 198313; KLEIN, SIPOS and FICUEROA 1984). TtCsh2” and Ttcsh2” each contain the wild- type allele for the tW5 lethal mutation derived from the tw12 parental chromo- some, and vice versa. These chromosomes should be the reciprocal crossover products of the chromosomes which carry the two lethals in cis. If not, the quasireciprocal products will share an undefined common region of chromo- some derived from one or the other parental chromosome and would thus differ from the trans heterozygote. Any decrease in viability could be attrib- uted to semideleterious alleles specific to one of the parental haplotypes which map to this overlap.
T o guard against the problem of the undefined overlap between pairs of crossovers, the two pairs of chromosomes described above were used in the four pairwise combinations. These would be likely to differ in their regions of overlap. In addition, male/female reciprocal crosses were performed to control for any maternal or paternal effects of these chromosomes.
SHIN et al. (1983b) found a statistically significant decrease in the level of complementation to 0.72 for trans heterozygotes of the tW5 and tWI2 mutations.
For the mice used here, the viability of the trans heterozygotes of tw12/tw5 was given by the following crosses: T/tW5 (male) X TtTu2tfl+ (female) resulted in 58 normal tail and 39 tailless progeny. T h e reciprocal cross of TtTu2tJ/+ (male) X +/tW5 (female) yielded 40 normal tail, 22 short tail and 28 tailless offspring. T h e overall complementation was 39
+
28/58+
22 = 0.84. This is not significantly different from 1.0 or the 0.72 value reported by SHIN et al. (1983b)(P
= 0.3 and 0.4, respectively, forx2
with d.f. = 1).I wished to see if the cis heterozygote differed from the trans in its comple- mentation properties. It is first necessary to determine if there is any decrease in viability simply resulting from the simultaneous heterozygosity of the two lethal mutations. This was done by placing the tcSh1” chromosomes opposite a non-t chromosome
17.
This is shown in Table 1 crosses of the form:TtCsh2”/+ (male) X T/tcSh’” (female).
Normal tail offspring (tcsh”’/+) are of the desired genotype. There is no male transmission ratio distortion in this cross. Therefore, if the animals in question show no decrease in viability, normal tail animals (tCsh”’/+) will be as frequent as their non-t siblings (T/+), which are distinguished by their short tails. This is true in all cases (Table 1, lines 1, 3, 5 and 7). Therefore, the combined effect of the two lethal mutations on tc5h101 and tcshlo2 is fully recessive to the wild-type chromosomes derived from our outbred stock.
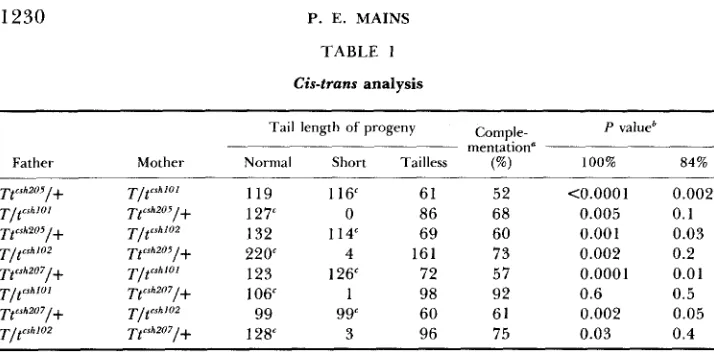
1230 P. E. MAINS TABLE 1
Cis-trans analysis
Tail length of progeny Comple- P value' mentation"
Father Mother Normal Short Tailless (%) 100% 84%
119 116' 61 52
1 27c 0 86 68
132 114" 69 60
220' 4 161 73
123 126' 72 57
106' 1 98 92
99 99" 60 61
128" 3 96 75
<0.0001
0.005 0.001 0.002 0.0001 0.6 0.002 0.03
0.002 0.1 0.03 0.2 0.01 0.5 0.05 0.4
a Tailless/control using TtcShZu5 or Tt""207 as a "wild-type" chromosome.
*
Statistical significance calculated by x 2 (d.f. = 1). Control value used to calculate complementation.provide t-derived materials for all loci, but lack either lethal mutation. In crosses described above, the ratio of tailless (Ttc5h200/tcsh100) to short tail (T/+) offspring is the measure of the viability of the cis heterozygote, with wild type derived from t chromosomes relative to their non-t sibs. In contrast to the cis heterozygotes with a non-t chromosome to provide wild-type function, a statis- tically significant decrease in the level of complementation was seen (Table 1, lines 1, 3, 5 and
7).
T h e viability of Ttcsh200/tc5h100 was also significantly different from the value seen for trans heterozygotes. If the t lethal mutations were acting as part of the same functional unit, the cis heterozygote should show the opposite result, an improved level of complementation, relative to the trans heterozygote.T o determine if there are any parental effects, male/female reciprocal crosses for each combination were performed:
T/tcshl" (male) X TtGsh2"/+ (female)
In this cross, the males transmit the t chromosome to >97% of their offspring. T h e ratio of tailless (Ttcsh200/tc5h100) to normal tail (+/t"hlOO) offspring is a mea- sure of the viability of the cis heterozygote, with wild type derived from t
chromosomes, relative to the wild-type viability. (In the previous crosses, the wild-type viability was assessed by the frequency of non-t sibs (T/+). Because of segregation distortion in the above cross, this class cannot be used. However, since t""hlo"/+ was shown above to have the equivalent viability, this quantity can be used.)
CIS-TRANS TEST OF THE T COMPLEX
DISCUSSION
1231
There is no cis-truns effect for tw5 and P I 2 : It has been proposed that the
t complex includes a set of functionally related genes which control embryo- genesis (GLUEKSOHN-WAELSCH and ERICKSON 1970; BENNETT 1975). SHIN et al. (1983b) concluded from the cis-trans test that several t-lethal mutations behaved as part of a functional unit. T h e cis-trans analysis reported here for the t-lethal mutations from the tw5 and twl' haplotypes found no evidence for such interactions.
Two recessive mutations are considered to be part of the same functional unit if an individual heterozygous for both mutations shows a wild-type phe- notype when the mutations are in cis, but displays the mutant phenotype when the mutations are in trans (LEWIS 1951; BENZER 1959). If the mutations are inactivating the same function, the trans heterozygote will behave as a homo- zygous mutation. T h e wild-type control places both mutant alleles in cis on one chromosome while the homologue contains the wild-type alleles of these loci in cis. Any semidominance resulting from heterozygosity of both mutations at once will be detected by this combination. If the homozygous mutations show high penetrance, the question of whether the trans heterozygote resem- bles the wild-type cis control or the homozygous mutants will be unambiguous. T h e phenotype of the trans heterozygote of tW5 and twl' closely resembles that of the wild-type control, rather than either homozygous mutation. T h e incomplete level of complementation between tW5 and twl' (0.84) reported here was not statistically significant. However, SHIN et al. (1983b) did report a statistically significant decrease in the level of complementation to 0.72 for these lethal mutations, and complementation of 0.84 reported here is not significantly different from this value. However, even this decrease is minor compared to the viability of either homozygous mutation. T h e viabilities of
tw5/tw5 or tw12/tw12 are probably much less than homozygotes have never been reported (BENNETT, DUNN and ARTZT 1976). Since the viability of tw5/ twI2 heterozygotes is at least 0.72, this represents an improvement of at least four orders of magnitude.
By the criteria used for the cis-trans test, the tW5 and twl' lethal mutations clearly belong to different functional units.
T h e minor decrease in the viability of the trans heterozygote is unlikely to reflect a rigorous requirement that the corresponding wild-type alleles be car- ried in cis to function efficiently. Instead, the effect may be due to additional modifier genes carried in the genetic background of the strains or within the
t haplotypes themselves.
T h e genetic background has been shown to affect the level of complemen- tation of t haplotypes. KLYDE (1970) outcrossed lines of t o and t"" which showed 28% complementation to genetically heterogeneous stocks. Sublines which showed complementation as low as 16% or as high as 72% were ob- tained. BENNETT and DUNN (1964) reported that the level of complementation of t o with different isolates of the twl' lethal mutation varied from 1-90%.
1232 P. E. MAINS
haplotypes are effectively isolated from the gene pool of mice d u e to the suppression of recombination with the wild-type form of chromosome 17,
which may allow them to accumulate additional deleterious mutations (PAIGEN
1980; TRES and ERICKSON 1982). In fact, t chromosomes have been found which carry multiple lethal or deleterious mutations (ARTZT, BABIARZ and
BENNETT 1979; BABIARZ, GARRISI and BENNETT 1982; SHIN et al. 1983b;
KLEIN, SIPOS and FIGUEROA 1984).
To test for these possibilities, SHIN et aE. (1983b) used two types of wild- type controls for the cis-trans test. One was to use a non-t chromosome to provide wild-type function in cis. This resulted in 100% viability when heter- ozygous with t-lethal mutations in cis. An additional control was to use a t chromosome lacking the specific lethal mutations to provide the wild-type func- tion for those loci, yet retaining t chromatin in the rest of the region. One such t chromosome lacking the specific lethals (derived from recombination between the t w z 2 and tw3' complementation groups) resulted in 100% viability when heterozygous with each of two doubly mutant chromosomes (carrying the lethal mutations for twl' and tw3' or for tW5 and t W z ' ) . This was interpreted by the authors as an argument against the possibility that other deleterious alleles, shared by the t haplotypes, are responsible for the decreased viability of the trans heterozygote.
T h e 1 chromosomes that supply the wild-type alleles of both lethal mutations must be the reciprocal crossover product of the chromosome carrying the two lethal alleles in cis. Otherwise they may share considerable overlap, derived from one o r the other parental chromosome. Because of the limited genetic resolution with the mouse, any pair of reciprocal recombinants will certainly share a large overlap, which could include the distance between the tW5 and the t w z 2 lethal alleles,
7
cM, an appreciable portion for the 15- to 20-cM tcomplex (ARTZT, MCCORMICK and BENNETT 1982; ARTZT 1984; CONDAMINE, GUENET and JACOB 1983). Overlapping recombinant chromosomes would con- trol for deleterious genes shared by both parental chromosomes, but not for deleterious genes present in only one of the parents.
To guard against problems of undefined overlap, two pairs of independent crossover products derived from tw5 and t W z 2 were used to provide the cis combinations of the lethal and wild-type alleles (Figure 1). T h e four pairwise combinations, which should vary the regions of undefined overlap, as well as male/female reciprocal crosses, were performed. Only one of the eight com- binations for these cis heterozygotes showed an increase, rather than a de- crease, in viability compared to the trans heterozygote (Table l). Therefore, even the minor cis-trans effect reported by SHIN et al. (1983b) was usually not observed.
CIS-TRANS TEST OF THE T COMPLEX 1233
type chromosomes could contain a fully viable allele which is completely dom- inant to the twl’ semilethal, Therefore, full viability would be seen for combi- nations which share a common region derived from tW5. Decreased viability would be seen for overlapping regions homozygous for tw” or heterozygous for tW5 and t w I 2 . There would be full viability when a non-t wild-type chro- mosome is used, but not for the trans heterozygote. Obviously, there are many variations on this theme, depending on how the recombinant chromosomes overlap. SHIN et al. (1983b) also reported two pairs of recombinant chromo- some which were known to have a considerable overlap of the t w J 2 haplotype (although outside of the region discussed here), and both showed decreased complementation. As the authors pointed out, this showed that the t w J 2 hap- lotype carries a deleterious allele for a gene(s) in addition to the known lethal mutation.
As discussed above, the differences in viabilities could also result from dif- ferences in genetic background outside the t complex. In the only case where
100% viability of the cis heterozygote was observed, the reciprocal male/female cross showed the opposite result. This may have resulted from fortuitous dif- ferences in modifier genes outside the t complex in individual mice (or stocks). T h e level of genetic variability could not be easily assessed from the data presented here. A large number of different individuals were used, so the number of offspring for individual pairs was usually small. In addition, one male was caged with two females. However, the most extreme variability that was seen was for the cross of tcshJoJ (male) X TtcshZo5 (female). T h e overall complementation was 0.68, but this included values from 0.2 (N = 30), 0.65 (N = 33) to 0.94 (N = 128) for different sets of mice (one male with two females).
Therefore, there are two explanations for the observed minor variation in complementation, both of which may be operating. Additional deleterious al- leles may be carried within t haplotypes, or the effect of genetic background outside of the t complex may influence the level of complementation. Either possibility seems much simpler than postulating that the tW5 and twJ2 lethal mutations are part of a single functional unit requiring cis interactions.
In all four cases shown in Table 1, the complementation was lower in the crosses where the female contributed the chromosome with two lethal muta- tions. This could be fortuitous, or it could show a slight semidominance of the lethal mutations. For example, a mother heterozygous for the two lethals may be slightly less healthy and so may not carry some of the slightly less healthy, doubly heterozygous embryos to term.
1234 P. E. MAINS
1980; TRES and ERICKSON 1982). An alternative, but not mutually exclusive, explanation is that there is actually natural selection in favor of these lethal mutations in wild mouse populations. This was suggested by the mathematical modeling of t chromosomes in wild populations (LEWONTIN and DUNN 1960;
LEWONTIN 1962, 1968).
T h e authors found that a t chromosome that resulted in homozygous male sterility, but which did not decrease viability, would be unstable if mice live in small demes. A chromosome that causes a high level of transmission ratio distortion and homozygous male sterility will have an equilibrium frequency of 0.7 in a large, randomly mating population. However, in small populations, genetic drift becomes important. Often, the t chromosome will drift to 1.0 among the males; therefore, all of the males in the deme will be sterile, and this deme will become extinct. Half of the demes became extinct in less than ten generations in the computer simulation. However, a recessive lethal mu- tation will greatly stabilize the chromosome by decreasing its equilibrium fre- quency and thereby decreasing the chances of it drifting to 1.0. A lethal allele decreased the equilibrium frequency to 0.45. In LEWONTIN’S model, the time for half of the demes to become extinct was increased from 10 to 80 genera- tions. Therefore, there may be a selective advantage conferred to t chromo- somes which acquire lethal mutations.
T h e effect of genetic drift in small demes is sensitive to the precise popu- lation structure, e.g., deme size and the amount of migration between demes
(LEWONTIN and DUNN 1960; LEWONTIN 1962; LEVIN, PETRAS and RASMUSSEN
1969; LACY 1978). However, it could explain why t haplotypes which contain a lethal or semilethal mutation are much more common than those which are viable.
These ideas suggest how the seemingly unrelated phenotypes of segregation distortion, male sterility and lethality may be interrelated. Arguments can also be made to include the other t-related phenotypes. First, segregation distortion must be associated with a deleterious phenotype (PAIGEN 1980). If not, the chromosome will rapidly become fixed in the population. These chromosomes would not be detected as causing transmission ratio distortion since the pop- ulation is no longer polymorphic for a chromosome sensitive to distortion. In the case of the t chromosomes, either the lethality or the male sterility could serve to prevent fixation of the t haplotype. All t chromosomes seem to have the same factors governing sterility, but differ in the lethal alleles that they carry. Therefore, it seems likely that the original t chromosome showed reces- sive male sterility, and then different lethal mutations were selected after the chromosomes spread. It is possible that the same factors which cause segrega- tion distortion when heterozygous cause sterility when homozygous (LYON
1984).
CIS-TRANS TEST OF T HE T COMPLEX 1235 that would rapidly become fixed in the population. There are also combina- tions of the segregation distortion factors which show 50% transmission when either heterozygous with wild type or when heterozygous with a complete t
chromosome; they are “immune” to the effects of segregation distortion. This recombinant chromosome would keep the frequency of the chromosome show- ing segregation distortion and male sterility very low (HARTL 1977). T h e seg- regation distortion system of Drosophila also shows recessive male sterility associated with crossover suppression (HARTL 1977).
In a sense, the tail interaction factor (t-int) is also an integral part of the t complex. This gene, which appears to be at a separate locus closely linked to T (BODE 1984), results in tailless T / t animals. t-int greatly facilitated the dis- covery and analysis of t haplotypes because it allowed the easy identification of t-bearing animals. It is possible that wild mice harbor other regions of the genome with properties like t haplotypes, but they have not been detected because they do not have such an easily scorable phenotype.
All t chromosomes are likely to have arisen from a common ancestor (NI-
ZETIC, FIGUEROA and KLEIN 1984). This is shown by the characteristics they all share which are not found in non-t mice. Different t haplotypes seem to share the same chromosomal rearrangement, as well as alleles governing seg- regation distortion and male sterility. t chromosomes also show a restricted set of closely related H-2 types (STRUM, FICUEROA and KLEIN 1982; NIZETIC,
FICUEROA and KLEIN 1985), polymorphic testis proteins (SILVER et al. 1983) and DNA restriction fragment length polymorphisms (FOX et al. 1985; HERR-
MANN et al. 1986). T h e only clear feature that differentiates among t haplo- types is the specific lethal alleles they carry.
Therefore, present-day t haplotypes may have arisen from an ancestral chro- mosome that displayed transmission ratio distortion held in check by male sterility. T h e system was preserved as a unit due to recombination suppression. T h e ancestral t chromosomes spread, and later, individual chromosomes were stabilized against the effects of genetic drift by acquiring different recessive lethal mutations.
This model does not necessitate a special role for the t complex during embryonic development, although such a role is not precluded. T h e data pre- sented in this paper do not indicate that the tW5 and twl’ lethal mutations are part of the same functional unit. T h e fact that so many genes required for embryogenesis map to the t complex does not necessarily imply a special de- velopmental role for this region. In Drosophila, it is estimated that 85% of all genes can mutate to lethal alleles (RAFF and KAUFMAN 1983). T h e t complex may be unique in that it represents a region of the mouse genome that is being “saturated” with lethal mutations due to its dynamics in natural popu- lations. Therefore, instead of playing a special role in development, the t
region may simply carry a random, unbiased set of lethal mutations.
1236 P. E. MAINS
Boulder). 1 also wish to thank DEBORAH LUKRALLE, CATHERINE JACKSON and CONSTANCE PARMA for maintenance of the mouse facility, and JUDITH WILSON for preparation of the manuscript. P.E.M. was supported by a Damon Runyon-Walter Winchell Cancer Fund postdoctoral fellowship (DRG 638).
LITERATURE CITED
ARTZT, K., 1984 Gene mapping within the T / t complex of the mouse. 111. t-lethal genes are arranged in three clusters o n chromosome 17. Cell 3 9 565-572.
ARTZT, K., B. BARBIARZ and D. BENNETT, 1979 A haplotype (tw7’) overlapping two complemen- tation groups. Genet. Res. 33: 279-285.
ARTZT, K., P. MCCORMICK and D. BENNETT, 1982 Gene mapping within the T/t complex of the mouse. 1. t-lethal genes are nonallelic. Cell 28: 463-470.
ARTZT, K., H.-S. SHIN and D. BENNETT, 1982 Gene mapping within the T/t complex of the mouse. 11. Anomalous position of the H-2 complex in t haplotypes. Cell 2 8 471-476.
Genetic analysis of the tW7j’ haplotype of the mouse using deletion mutations: evidence for a parasitic lethal mutation. Genet. Res. 39: 1 1 1- 120.
BABIARZ, B., G. J. GARRISI and D. BENNETT, 1982
BENNETT, D., 1975
BENNETT, D. and L. C. DUNN, 1964
BENNETT, D., L. C. DUNN and K. ARTZT, 1976
T h e T-locus of the mouse. Cell 6: 441-454.
Repeated occurrences in the mouse of lethal alleles of the
Genetic change in mutations at the T/t-locus in
O n the topology of the genetic fine structure. Proc. Natl. Acad. Sci. USA 45:
Ethylnitrosourea mutagenesis and the isolation of mutant alleles for specific genes located in the T region of mouse chromosome 17. Genetics 1 0 8 457-470.
Recombination between two mouse t haplo- types (t““’tfand ti”-’): Segregation of lethal factors relative to centromere and tufted
(tf)
locus. Genet. Res. 42: 335-344.Fox, H. S., G. R. MARTIN, M. F. LYON, B. HERMANN, A.-M. FRISCHAUF, H. LEHRACH and L. M.
FRISCHAUF, A.-M., 1985
GLUECKSOHN-WAELSCH, S. and R. P. ERICKSON, 1970 T h e T-locus of the mouse: implications for
HARTL, D. L., 1977 Mechanism of a case of genetic coadaptation in populations of Drosophila
HERRMANN, B., M. BUCAN, P. E. MAINS, A.-M. FRISCHAUF, L. M. SILVER and H. LEHRACH, Genetic analysis of the proximal portion of the mouse t complex: evidence for a second
T h e control of differentiation by the T complex. lmmunol.
Polymorphism o f t complex genes in European wild
Selection in mice for change in viability of a semilethal genotype at the T
Dynamics of t-alleles in Mus musculus population: review and speculation. same complementation groups. Genetics 49: 949-958.
the mouse. Genetics 83: 361-372.
BENZER, S., 1959
BODE, V. C., 1984 1607-1620.
CONDAMINE, H., J.-L. GUENET and F. JACOB, 1983
SILVER, 1985 Molecular probes define regions of the mouse t complex. Cell 40: 63-69.
T h e T / t complex of the mouse. Trends Genet. 1: 100-103.
mechanisms of development. Cur. Top. Dev. Biol. 5: 281-316.
melanogaster. Proc. Natl. Acad. Sci. USA 74: 324-328.
1986
inversion within t haplotypes. Cell 44: 469-476.
Rev. 33: 70-104.
KLEIN, J. and C. HAMMERBERG, 1977
KLEIN, J., P. SIPOS and F. FIGUEROA, 1984
KLYDE, G. J., 1970
LACY, R. C., 1978
mice. Genet. Res. 44: 39-46.
locus. J. Hered. 61: 39-43.
CIS-TRANS TEST OF THE T COMPLEX 1237
LEVIN, B. R., M. L. PETRAS and D. I. RASMUSSEN, 1969 The effects of migration on the main- tenance of a lethal polymorphism in the house mouse. Am. Nat. 103: 647-661.
LEWIS, E. B., 1951 Pseudoallelism and gene evolution. Cold Spring Harbor Symp. Quant. Biol.
16: 159-174.
LEWONTIN, R. C., 1962 Interdeme selection controlling a polymorphism in the house mouse. Am. Nat. 9 6 65-78.
LEWONTIN, R. C., 1968 The effects of differential viability on the population dynamics of t alleles in the house mouse. Evolution 22: 262-273.
LEWONTIN, R. C. and L. C. DUNN, 1960 The evolutionary dynamics of a polymorphism in the house mouse. Genetics 45: 705-722.
LYON, M. F., 1981 The t-complex and the genetic control of development. Symp. Zool. Soc.
LYON, M. F., 1984 Transmission ratio distortion in mouse t-haplotypes is due to multiple distorter genes acting on a responder locus. Cell 37: 621-628.
MAINS, P. E., 1986 A region flanking H-2K is duplicated to a distant site in most mouse t haplotypes. Immunogenetics 25: 357-363.
NIZETIC, D., F. FIGUEROA and J. KLEIN, 1982 Evolutionary relationships between the t and H-2
haplotypes in the house mouse. Immunogenetics 19: 31 1-320.
PAIGEN, K., 1980 Temporal genes and other developmental regulators in mammals. pp. 419- 470. In: The Molecular Genetics of Development, Edited by T. LEIGHTON and W. F. LOOMIS. Academic Press, New York.
PLA, M. and H. CONDAMINE, 1984 Recombination between two mouse t haplotypes (t""tf) and
tL4-'): mapping of the H-2 complex relative to centromere and tufed (tj) locus. Immunogenetics Lond. 47: 455-477.
2 0 177-185.
RAFF, R. A. and T. C. KAUFMAN, 1983 Embryos, Genes and Evolution. Macmillan, New York.
SHERMAN, M. I. and L. R. WUDL, 1977 T-complex mutations and their effects. pp. 136-234. In: Concepts in Mammalian Embryogenesis, Edited by M. I. SHERMAN. MIT Press, Cambridge, Massachusetts.
SHIN, H.-S., D. BENNETT and K. ARTZT, 1984 Gene mapping within the T / t complex of the
Inversion in the H - 2
mouse. IV. The inverted MHC is intermingled and several t-lethal genes. Cell 39: 573-578.
SHIN, H.-S., L. FLAHERTY, K. ARTZT, D. BENNETT and J. RAVETCH, 1983a
complex of t-haplotypes in mice. Nature 306: 380-383.
SHIN, H.-S., P. MCCORMICK, K. ARTZT and D. BENNETT, 1983b Cis-trans test shows a functional relationship between non-allelic lethal mutations in the T/t-complex. Cell 33: 925-929.
proteins specified by genes within the mouse t complex. Cell 35: 35-45.
wild mice. I. The H-2 haplotypes of 20 t-bearing strains. Genet. Res. 40: 73-88.
transmission ratios. Genet. Res. 3 8 315-325.
discloses no inversions. Nature 2 9 9 752-754.
SILVER, L. M., J. UMAN, J. DANSKA and J. I. GARRELS, 1983
STRUM, S., F. FICUEROA and J. KLEIN, 1982
STYRNA, J. and J. KLEIN, 1981
TRES, L. L. and R. P. ERICKSON, 1982
WINOTO, A., M. STEINMETZ and L. HOOD, 1983
A diversified set of testicular cell
The relationship between t and H-2 complexes in
Evidence for two regions in the mouse t complex controlling
Electron microscopy of t-allele synaptonemal complexes
Genetic mapping in the major histocompatibility complex by restriction enzyme site polymorphisms: most mouse class I genes map to the T l a complex. Proc. Natl. Acad. Sci. USA 8 0 3425-3429.