MULTIPLE DIFFERENTIATION O F PLASMONS OF DIPLOID SPECIES OF SOLANUM1
PAUL GRUN. MARY AUBERTIN AND ANNE RADLOW
Department of Botany and Plant Pathology, Pennsylvania State Uniuersity, Uniuersity Park, Pennsylvania
Received May 14, 1962
TUDIES of plasmon variations in Epilobium (MICHAELIS 1954; CASPARI 1948), Oenothera ( STINSON 1960; STUBBE 1960), Streptocarpus ( OEHLKERS 1938; OEHLKERS and EBELL 1958), Solanum (KOOPMANS 1951,1952,1954,1955, 1959; BUCK 1960), Clarkia (HIORTH 1948), yeast (EPHRUSSI 1953), and Culex ( LAVEN 1959) have demonstrated that heritable plasmon variations exist and are of evolutionary importance. A further understanding of the role of plasmon variations will require a method for tracing the consecutive steps through which plasmon changes occur. This, in turn, will require that one be able to identify the individual unit characters of which a given plasmon is composed and formu- late what might be termed a plasmantype, or description of a plasmon.
The analysis presented here is the first step of a n attempt to describe such plasmon characters of diploid tuber-bearing species of Solanum. These plants have the advantage that a considerable number of closely related species can be crossed (SWAMINATHAN and HOWARD 1953; HAWKES 1956a; HOWARD 1960; GRUN 1961) to produce fertile hybrids and, therefore, a diversity of genetic and plasmon types can be subject to test.
Terminology
One roadblock to analysis of plasmons is posed by our present lack of a symbol system to describe multiple characteristics of the same Dlasmon. Designations as generally used have identified plasmons by naming their sources or places of origin rather than by describing their characteristics. The individual plasmon characters dealt with here were identified by their interactions with specific genes. It was, therefore, decided to identify each plasmon character by the symbol used to designate the gene with which it interacts. The following example can be used to illustrate the method: A gene has been found which causes deformed flowers when it is present i n the homozygous recessive condition and in a plasmon sensitive to its action. The gene for the deformed flowers was designated by the symbol df, and its allele for normal flowers
of.
The sensitive plasmon was designated dfe, the lower case letter being used since it interacted with the reces- sive allele of the gene, and the superscript s inserted to indicate that the plasmon1Authorized for publication April 9, 1962 as paper No. 2652 i n the Journal Series of the Pennsylvania Agricultural Experiment Station.
1322 P. G R U K , R I . A U B E R T I N A N D A . RADLOW
was sensitive. The resistant plasmon was symbolized, accordingly, as
Df
'.
Each plasmon symbol preceded its respective gene symbol in the formula and was separated from it by a hyphen Thus, df '-df df is the formula of a deformed flower plant while all other plasmon-gene combinations are phenotypically normal. When a dominant gene interacted with a plasmon to produce a n abnormal pheno- t y p , the recessive gene being normal. the sensitive plasmon was designated by the capital letter and the nonsensitive by the lower case letter. Thus. there is adominant gene for indehiscence of anthers (alleles In and in) expressed only in
Ins plasmon. Indehiscence would result from the Zn'-Zn In o r Zns-Zn in combina- tion of plasmms and genes, while the plasmon-gene combination Zn'-in in or in'
plasmon with a n y combination of genes would produce dehiscence.
MATERIALS A N D METHODS
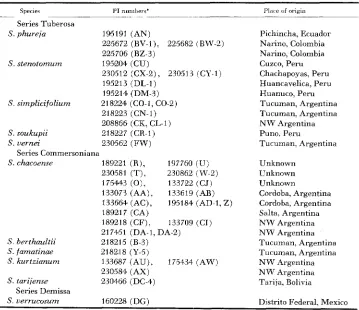
T h e identities and places of origin of the plants used are listed in Table 1 with their U S. Department of Agriculture Plant Introduction numbers. Crosses were carried out in the greenhouse using techniques already described (GRUN 1961;
TABLE 1
Identification and places o f origin of clones used
Plai e of origin
~ _ _
PI numbeis'
.. ~- Species
Series Tuberosa
S. phureja 195191 (AN) Pichincha, Ecuador
225672 (BV-1). 225682 (BW-2) Narino, Colombia
225706 (BZ-3) Narino, Colombia
230512 (CX-e), 230513 (CY-1) Chachapoyas, Peru
195213 (DL-I) Huancavelica, Peru
195214 (DM-3) Huanuco, Peru
S. simplicifolium 218224 (CO-1, CO-2) Tucuman, Argentina
218223 (CN-1) Tucuman, Argentina
208866 (CK, CL-1) N W Argentina
S . stenotomum 1952Q4 (CU) Cuzco, Peru
S. soukupii 218227 (CR-I) Puna. Peru
S. uernei 230562 (FW) Tucuman, Argentina
S. chacoense 1898221 ( R ) , 197760 ( U ) Unknown
230581 (T); 230862 (W-2) Unknomwn 175443 (0), 133722 (CJ) Unkno.ivn
133073 (AA), 133619 (AB) Cordoba, Argentina 133664 (AC), 195184 (AD-1, Z ) Cordoba, Argentina
189217 (CA) Salta, Argentina
189218 (CF), 133709 (CT) N W Argentina
217451 (DA-1, DA-2) N W Argentina
Series Commersoniana
S . berthaultii 218215 (B-3) Tucuman, Argentina
S. famatinae 218218 (Y-5) Tucuman, Argentina
S. kurtzianum 133687 (AU), 175434 (AW) N W Argentina
230584 (AX) N W Argentina
S. tarijense 230466 (DC-4) Tarija, Bolivia
S . uerrucosum 160228 (DG) Distrito Federal, Mexico
P L A S M O N A N D G E N E I N T E R A C T I O N 1323
GRUIV and RADLOW 1961). All of the parents in the crosses listed in the Tables were individuals haying normal fertile phenotypes. Seeds were germinated in the greenhouse. seedlings were established in peat pots and transplanted to the field for observation. Progenies were established in replicated plantings the first year of test, but since the characters being tabulated turned out to have definite expres- sion with no differences between replicates, the plantings of later years were not replica’ ed.
E X P E R I M E N T A L RESULTS
Inheritance of deformed flowers: Flowers of plants showing the character de- formed flowers either lacked anthers or had anther lengths reduced to four mm or less. T h e difference between the normal and deformed flower phenotypes was discontinuous so that plants could be readily classified in one category or the other. Abnormal corollas. slightly shortened styles, and female sterility were sometimes associated with the deformed flowers.
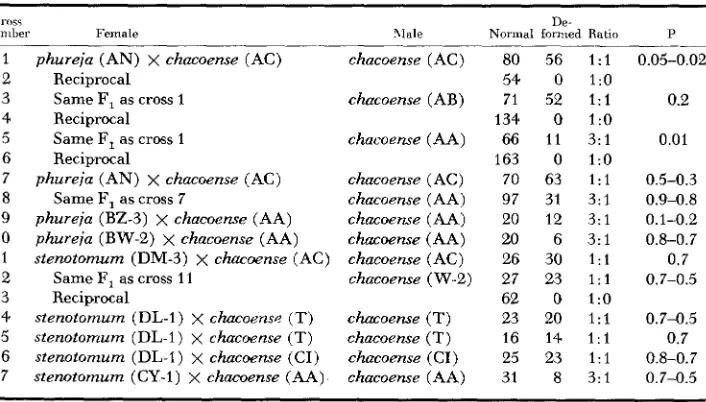
The fact that deformed flowers resulted from a gene-plasmon interaction is shown by a comparison of reciprocal first generation backcrosses 1 with 2 , 3 with 4, 5 with 6, and 12 with 13 of Table 2. Except when a given F, has been used more than once, as i,n crosses 1 and 3, F17s of the same parentage can be assumed to be sibs. Each of the reciprocal sets was produced by using the same F, and S.
chacoense individuals, but reversing the direction in which the cross was made. The deformed flower characteristic consistently appeared when some form of
S. phureja-stenotomum complex was the original plasmon parent, but did not appcar when S. chacoense was the plasmon parent.
The backcross ratios of normal to deformed flowered plants of Table 2 show a good fit. in all but two crosses (crosses 1 and 5 ) to either a 1: 1 or a 3: 1 ratio. The
TABLE 2
Inheritance of deformed flowers
>\lale Normal fornled Ratio De- P
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
phureja (AN) x chucoense (AC) Reciprocal
Same F, as cross 1 Reciprocal Same F, as cross 1 Reciprocal
Same F, as cross 7
phureja (AN) x chacoense (AC) phureja (BZ-3) x chacoense (AA) phureja (BW-2) x chacoense (AA) stenotomum (DM-3) x chaccrense (AC)
Same F, as cross 11 Reciprocal
sienotomum (DL-1) x chacoerzs~ (T) sienotomum (DL-1) x chacoense (T) stenotomum (DL-1) x chacoense (CI) stenotomum (CY-1) x chacoense (AA)
chacoense (AC) chacoense (AB) chacoense (AA) chacoerrse ( AC) chacoense (AA) chacoense (AA) chucoense (AA) chacoense (AC) chacoense (W-2) chucoense (T) chacoense (T) chacoense (CI )
chacoense (AA)
80 56 1:l
54 0 l : o 71 52 1 : l 134 0 1:0 66 1 1 3 : l 163 0 1:0 70 63 1:l 97 31 3 : l 20 12 3:l
U) 6 3:l
26 30 1:l
27 23 1 : l
62 0 1:0 23 20 1 : l 16 14 1 : l 25 23 1 : l 31 8 3 : l
1324 P. G R U N , M . A U B E R T I N A N D A. R A D L O W
1 : 1 can best be interpreted by assuming that the phureja parent was dfs-Df Df, the chacoense Dfr-df df and the F, df"-Df df. The backcross using the F, as female, would segregate 1 df"-Df df (normal) : 1 df"-df df (deformed). The reciprocal backcrosses would be of the form Dfr-df df x df"-Df df, and since all progeny would be in Of' plasmon, all would be normal. The 3: 1 backcross ratios of Table 2 can best be interpreted as resulting from heterozygosity of the chacoense parent for this gene, the backcross having the form df"-Df df x of'-Df df. Only one S.
chacoense individual, plant AA, was heterozygous for the deformed gene, and it regularly produced the 3: 1 backcross progeny.
Crosses 1 and 5 in Table 2 do not fit the expected ratios, there being in both a deficiency of deformed flower plants. Progeny of both crosses also differed from the other progeny in having plants that did not flower because buds fell off before the flowers opened. It seems likely that these bud-abscission plants were an ex- treme expression of flower deformation. There were 16 such bud-abscission plants in cross 1 and 21 in cross 5 .
Deformed flowers are conditioned, then, by one recessive gene present in certain
S. chacoense plants and effective in the plasmons of certain plants of S. phureja and S. stenotomum. With the knowledge that plants AA, AB, and AC have the df gene, appropriate backcrosses could be made to determine whether other diploid species of Solanum have Df' or df" plasmon. The results of such crosses to test plants of S. berthaultii, famntinae, kurtzianum, simplicifolium, and uerrucosum appear in Table 3. It is apparent that all of the above species have Of' plasmon since deformed flowers did not occur in any of the backcross progenies.
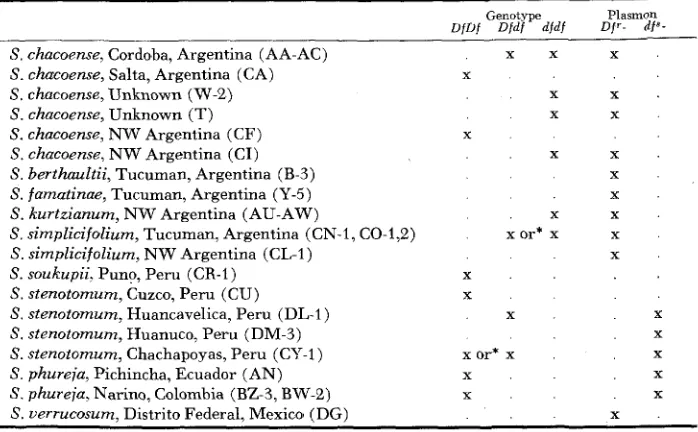
Given the information that certain plants of S. phureja and stenotomum have the df" plasmon, testcrosses could be made to determine the genotypes of unknown plants. Table 4 summarizes the geographic and species distribution of genes and plasmons for deformed flowers. Plants are listed, with the exception of those
TABLE 3
Progenies showing plasmon resistance to deformed flower genes
Cross number Female (17,) Male Normal Deformed
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
berthaultii (B-3) x chacoense (AC) berthaultii (B-3) x chucoense (AC) famatinae (Y-5) x chacoense (AC) famatinae (Y-5) x chacoense (AC) famatinae (Y-5) x chacoense (AC) kurtzianum ( A U ) x chacoense (AC) simplicifolium (CK) x chacoense (AC) simplicifolium (CL-1) x chacoense (AB) simplicifolium (CL-I) x chacoense (AB) simplicifolium (CN-1) X chacoense (AB) simplicifolium (CO-2) X chacoense (AC) simplicifolium (CO-2) x chucoense (AC) uerrucosum (DG) x chacoense (AB) uerrucowm (DG) x chaceense (AB) uerrucosum (DG) x chacoense (AB)
chacoense (AC) chacoense (AC) chacoense (AC) chacoense ( AC) chacoense (AC) chacoense (AC) chucoense (AC) chacoense (AA) chacoense (AC) chucoense (AA) chacoense (AC) chacoense (AC) chacoense ( AC) chucoense (AA) chacoense (AA)
P L A S M O N A N D G E N E I N T E R A C T I O N
TABLE 4
Distribution of deformed flower genes and plasmons
1325
Genotype Plasmon DfUf D f d f djdf of‘- d f a -
S . chacoense, Cordoba, Argentina (AA-AC) x x X S. chacoense, Salta, Argentina (CA)
S. chacoense, Unknown (W-2) x x .
S. chacoense, Unknown (T) X x
S. chacoense, N W Argentina (CI) . x x .
S . berthaultii, Tucuman, Argentina (B-3) S. famatinae, Tucuman, Argentina (Y-5)
S. kurtzianum, N W Argentina (AU-AW) . x x .
S . simplicifolium, Tucuman, Argentina (CN-1, CO-1,2) x or* x x
S. simplicifolium, NW Argentina (CL-I) S. soukupii, Pun?, Pen1 (CR-1)
S. stenotomum, Cuzco, Peru (CU)
S . stenotomum, Huancavelica, Peru (DL-1) x x
S. stenotomum, Huanuco, Peru (DM-3)
S. stenotomum, Chachapoyas, Peru (CY-1) x or* x x
S. phureja, Fichincha, Ecuador (AN) x . . x
S. phureja, Narino, Colombia (BZ-3, BW-2) x . x
S. uerrucosum. Distrito Federal, Mexico (DG)
X
S. chacoense, N W Argentina ( C F ) X
x
x .
x
x .
x
x
x .
~~~
* Precise genotype still uncertain
from unknown locales, in south-north order. The dfs plasmon has been found. to date, only in the phureja-stenotomum complex whereas D f ’ occurs in a wider range of species. While the df allele occurs frequently among plants having Df’
plasmon. it is, for obvious reasons, less common in plants having the dfs plasmon.
Inheritance of anther indehiscence: Anther dehiscence was tested by determin- ing whether pollen came out when the pore ends of anthers of open flowers were knocked against a clean glass slide. A minimum of five flowers per plant were tested in this manner. The indehiscent plants were often found to have yellow or slightly greenish anthers instead of the normal orange ones, There was also fre- quently a shortening, by approximately one millimeter, of anthers of indehiscent plants. This amount of shortening was very much less than the reduction by three or more millimeters that was typical of plants having deformed flowers.
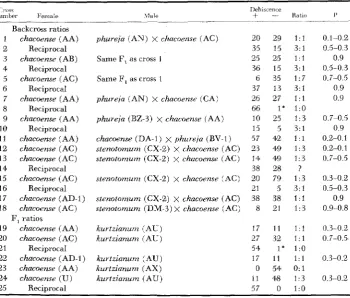
The ratios of dehiscent to indehiscent plants found in reciprocal backcrosses and some F, progenies are listed in Table 5. The word “Reciprocal” by a given cross again indicates that the same individuals were used as in the cross of the line above, but in the reciprocal direction. When a given backcross pro2eny contained plants showing deformed flowers, only the normal nondeformed individuals were included in the dehiscence study.
1326 P . G R U N , M . AUBERTIiV A N D A. RADLOW
TABLE 5
Backcross and F , ratios of dehiscent: indehiscent plants
~ ~~~ ~ ~
(.I-LIEF Dehiscence
+
-
Ratio I'~ __..__--
..
hlale
.- nuniber Female
Backcross ratios
1 chacoense (AA) phureja (AN) x cliacoense (AC) 20 29 1:1 0.1-0.2
2 Reciprocal 35 15 3:l 0.5-0.3
3 chacoense (AB) S a m e F, as cross 1 25 25 1 : l 0.9
6 Reciprocal 37 13 3:l 0.9
7 chacoense (AA) phureja (AN)
x
chacoense (CA) 26 27 1:l 0.9I O Reciprocal 15 5 3 : l 0.9
4 Reciprocal 36 15 3 : l 0.5-0.3
5 chacoense (AC) S a m e F, as cross 1 6 35 1:7 0.7-0.5
8 Reciprocal 66 I * 1:0
9 chacoense (AA) phureja (BZ-3) x chacoense (A A) 10 25 1:3 0.7-0.5 11 chacoense (AA) chacoense (DA-I) x phureja (BV-1) 57 42 1 : l 0.2-0.1 12 chacoense (AC) stenotomum (CX-2) x chacoense (AC) 23 49 1:3 0.2-0.1 13 chacoense (AC) stenotomum (CX-2) x chacoense (AC) 14 49, 1:3 0.7-0.5
14 Reciprocal 38 28 ?
15 chacoense (AC) stenotomum (CX-2) x chacoense (AC) 20 79 1:3 0.3-0.2
16 Reciprocal 21 5 3:1 0.5-0.3
17 chacoense (AD-I) stenotomum (CX-2) x chacoense (AC) 38 38 1:1 0.9 18 chacoense (AC) stenotomum (DM-3) x chacoense (AC) 8 21 1:3 0.9-0.8
19 chacoense (AA) kurtzianum (AU) 17 11 1:1 0.3-0.2
20 chacoense (AC) kurtzianum (AU) 27 32 1 : l 0.7-0.5,
F, ratios
21 Reciprocal 54 I * l : o
23 chacoense (AA) kurtzianum (AX) 0 54 0:l
25 Reciprocal 57 0 l : o
22 chacoense (AD-1 ) kurtzianurn (A U ) 17 11 1:l 0.3-0.2
24 chacoense (U) kurtzianum (AU) 11 48 1:3 0.3-0.2
* T h e few exceptional indehiscent plants are felt to hare resdtrd froin iegicgatimi of gem1 s i olher than the pl.iimr~n-
\ e n s : i w f a < t 11s being wriildered here.
and 6, show a n occasional sterile plant, though the ratio observed essentially ex- cludes steriles. The cause of such sterile exceptions is unknown, but it is probably outside of the province of the factors being considered here. One cross. cross 14, has not bceu interpretable. The ratios in S chacosnse plasmon are generally, however, those to be expected of dominant genes for indehiscence. while those in S. phureja and S. stenotomum plasmons point to recessive indehiscence genes. 1. S. chacoense plasmons: T h e 1 : 1 ratios can be interpreted by assuming that
S. chacomsc has the plasmon that is sensitive to dominant genes for indehiscence,
I n y , and homozygous recessive genes, in in, to produce normal flowers; while
S. phureja has in'-Zn In, the recessive resistant plasmon and homozygous domi- nant genes. I t is necessary to assume that the gene responsible for indehiscence is dominant because in the backcross chacoense x (phureja: x chacoense) a recessive
phureja gene would not be expressed, while a recessive chacoense gene would have been expressed in the chacoense parent itself. The F, phureja x chacoense
P L A S M O N A N D GENE I N T E R A C T I O N 1327
plasmon would segregate 1 Ins-Zn
in
(indehiscent) : 1 Zns-inin
(normal). The1:3 ratio can be interpreted on the same basis with ihe added ;Issumption that
the S . phureja or S. stenotomum parent has two dominant genes, either of which
can lead to indehiscence in Ins plasmon. Thus S. chacoense should be symbolized
Ins& in in' in', the S. phureja inr-Zn In In' In', and the phureja x chacoense F,
in'-Zn in In' in'. Finally, the 1 : 7 ratio (cross 5 ) points to a comparable mecha-
nism, but requiring the operation of one of three genes, In, Znl, or I n 2 in Ins plas- mon to produce indehiscence. Since the same F, was used for cross
5
as for crosses 1 and 3, both of which produced 1 : 1 ratios, it seems likely that the plasmon and genes of S. chucoense plant AC are sensitive i o two more dominant genes for indehiscence than are S. chacoense plants AA and AB. This could either indicate that AC plasmon is different from AA and AB plasmons, or that AA and ABhave dominant protector or restorer genes lacked by AC.
Crosses 19-25 of Table 5 include reciprocal F,'s of S. kurtzianum and S. cha- coense. The contrasts of crosses 20 with 21, and 24 with 25 indicate that indehis- cence of these Fl's is conditioned by plasmon factors. The 1 : 1 ratios of crosses
19,20, and 22 suggest that S. kurtzianum plant AU is heterozygous for one domi- nant gene that produces indehiscence in the S. chacoense plasmons tested. T h e
100 percent indehiscence of cross 23 suggests that plant AX of S. kurtzianum is homozygous for dominant indehiscence gene (s) and the 1 : 3 ratio (cross 24) that plant AU is heterozygous for two genes that cause indehiscence in the plasmon of
S . chacoense plant U. It has still to be determined whether the dominant factors
of S . kurtzianum are the same as those of S. phureja and stenofomnm.
TABLE 6
Progenies showing resistance to genes that condition indehiscence
Cross
numher Female
~ ~~
Dehiscence
Male
+ - -
1 chacoense (DA-3) phureja (BZ-3) 6 0
2 chacoense (DA-3) X phureja (.BZ-3) chacoense (CA) 16 0
3 chacoense (DA-3) phureja (BV-1) 7 0
4 chacoense (DA-3) x phureia (BV-1) chacoense (CA) 44 0
5 chacoense (DA-I ) phureia (BV-I) 7 0
6 chacoense (DA-I)
x
phureja (BV-1) chacoense (AA) 71 07 chacoense (AA) chacoense (DA-1) x phureja (BV-1) 57 42
8 chacoense (W-2) stenotomum (DM-3) x chacoense (AC) 60 2*
9 chacoense (AC) Same F, as cross 8 7 8
11 chacoense (R) kurtzianum ( A X ) 15 1'
12 stenotomum (CU) stenotomum (DM-3) x chacoense (AC) 48 3*
13 stenotomum (CU) stenotomum (DM-3) x chacoense (AC) 51 0
14 chacoeme (AC) Same F, as cross 13 8 21
10 chacoense (R) kurtzianum (AU) 53 0
15 simplicifolium (CK) kurtzianum (AU) 9 0
16 s i m p l i c i f o h m (CK) kurtzianum (AX) 5 0
1 7 tarijense (DC-4) kurtzianum (AU) 57 0
18 Recipmal 2 0 0
1328 P. G R U N , M. A U B E R T I N A N D A. RADLOW
The results of crosses listed in Table 5 established that the following plants have dominant gene(s) for indehiscence in sensitive In' plasmon: S. phureja
plants AN. RZ-3, and BV-1, S. stenotomum plants CX-2 and DM-3, and S. kurt- zianum plants AU and AX. Some of these plants were used to determine whether other biotypes available had In' or in' plasmons (Table 6 ) . S. chacoense appears not to be a uniform species, for while the chwoense plants AA-AD-1 have plasmon I n S (Table 5 ) , chacoense plants DA-1, DA-3, W-2, and R appear to have
in' plasmon. Cross 7 (Table 6) is included f o r comparison with cross 6 since it points up this difference between the Zn' plasmon of S. chacoense plant AA and the in' plasmon of DA-1. Crosses 9 and 14 are also listed for comparison with those that directly precede them. The in' plasmon was also found (Tables 5 and 6 ) in tested biotypes of S. phureja, stenotomum, kurtizianum, simplicifolium.
and tarijense.
2. S. phureja and S. stenotomum plasmons: The 3: 1 ratios of crosses 2,4, 6, 10, and 16 (Table 5 ) are recessive ratios showing that there are recessive genes of
S. chacoense that lead to indehiscence in S. phureja or stenotomum plasmons. Two alternative interpretations could at the start explain these 3: 1 ratios. One assumes that the chacoense parent is heterozygous for a gene which. in the homo- zygous recessive condition in phureja or stenotomum plasmon, could lead to indehiscence. This is designated the pl (pollenless) factor to distinguish it from the dominant indehiscent factor. S. chacoense would be PIr-PI p l (normal), S.
phureja. with sensitive plasmon, would be pls-Pl PI (normal), and the F, phureja x chacoense could be pls-Pl pl or pl'-PI PI. If the heterozygous F, is backcrossed using chacosnse as the male, one fourth of the progeny will be pl'-pl pl, and SO pollenless. The alternative interpretation would assume a two-gene mechanism:
S. phureja would be pls-Pl PI Pl1 PI1, S. chacoense PP-pl p l p l z pll, the F, pls-Pl pl PI1 pl'. and, thus, one fourth of the offspring following backcrossing by S. chacoense male would be pl"-pl p l p l L p l z (pollenless). To distinguish between these alternatives, second backcross progenies were grown using the pollen- producing sibs of pollenless plants of the first generation backcross as female parents. According to the one gene hypothesis, such dehiscent sibs could be either
pP-Pl PI or pl'-Pl pl. The former class would not segregate in the second back- cross. and the latter would. According to the two gene hypotheis, the pollen- producing sibs could be pl'-Pl pl Pl2 p l ' , pl'-Pl p l pi2 p l l , or pl'-pl pl PI1 p l l , all of which should segregate (3: 1, 1 : 1, and 1 : 1 respectively) in the succeeding back- cross generation. Of five second backcross progenies raised, two were essentially nonsegregating with ratios of 33:O and 31: 1, a result which favors the former of the two hypotheses. The 31 : 1 ratio is no fit to the 3: 1 or 1 . 1 and the one pollenless plant probably resulted from segregation of factors other than those being con- sidered here. To date the PI' plasmon has been located in S. chacoense (Table 5 )
while p l A has been found in S. stenotomum, phureja. and uerrucosum.
Distribution of lobed male sterility: A single gene exists i n certain clones of
S. chacoense, kurtzianum, phurejo, simplicifolium, stenotomum, and torulpanum
P L A S M O N A N D G E N E I N T E R A C T I O N 1329
spores are regularly produced by cytoplasmic male sterile plants of this type and they have a diagnostic four-lobed appearance. Frequently, such spores do not take up propio-orcein stain. Using the symbol system adopted here, S. uerrucosum can be designated Lba-Zb Zb, having plasmon sensitive to the dominant Lb (lobed) allele, and being itself homozygous recessive. S. chacoense is Zbr-Lb Lb, and male sterile F,’s of uerrucosum X chacoense Lbs-Lb lb.
Tests were run by attempting to make reciprocal F, crosses between S. uer- rucosum from Tlaxcala, Mexico and other biotypes used in these tests. As has previously been discussed ( GRUN and RADLOW 1961 ) these FI7s can be produced readily when S . uerrucosum is the female parent, but usually the reciprocals can not be produced. The lobed sterility appeared in all interspecific Fl’s tested hav- ing uerrucosum as female, and the three reciprocal combinations produced were normal and fertile (Table 7 ) .
DISCUSSION
The deformed flower and dominant indehiscence characters described here are closely related to those described by KOOPMANS (1951, 1952, 1954, 1955, and
1959), and the data presented constitute a confirmation of her finding that they
are conditioned by gene-plasmon interactions. As described here the characters have a simple one to three gene inheritance, while KOOPMANS’ data were inter- preted on a polygenic basis. Differences i n the parental material used or environ- mental modifications of expression may account for the contrasts in data. One manifestation of a difference in expression is the fact that the normal and de- formed flower phenotypes were distinct in the garden at University Park, Pa., while
KOOPMANS’
plants showed continuous variation and so were difficult to classify.TABLE 7
Sterility of crosses using S. verrucosum, Tlaxcala, Mexico as female parent
Male Parent Sterile Fertile
S. chncoense, Unknown (W, U)
S. chacoense, Cordoba, Argentina (Z-AD)
S. chacoense, Unknown (0, T )
S . chacoense, Salta, Argentina (CA)
S. chacoense ssp. subtilius, Unknown (CI, CJ)
S. chacoense ssp. subtilius, Unknown (DA)
S. stenotomum, Huancavelica, Peru (DL)
S. stenotomum, Chachapoyas, Peru (CX)
S . k u r t z i a “ , Unknown (AV) S. famatinae, Tucuman, Argentina (Y)
S. simplicifolium, Tucuman, Argentina (CO)
S. uernei, Tucuman, Argentina (FW) Reciprocal
Reciprocal
Reciprocal
3 0
0 2
2 0
0 34
10 0
2 0
9 0
0 14
4 0
20 0
10 0
3 0
6 0
1 1 0
1330 P. G R U N , M. A U B E R T I N A N D A. R A D L O W
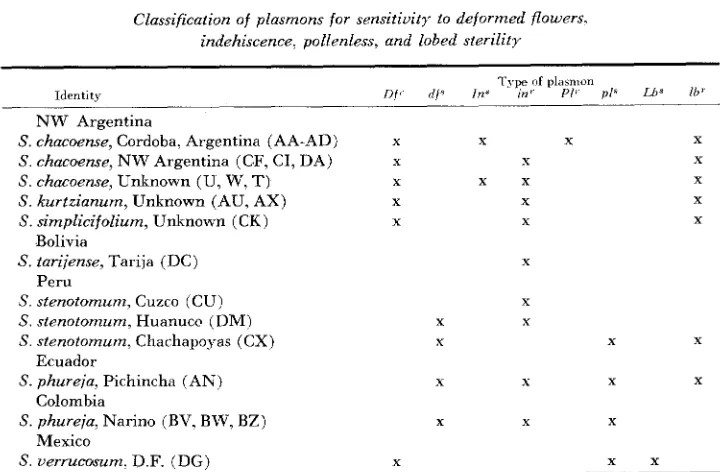
TABLE 8
Classification of plasmons for sensitivity io deformed flowers, indehiscence, pollenless, and lobed sterility
1 ype of pldrnion
Identrtj Jlf' d f s IN" ~ n ' Pi' p l R Lb' lb' NW Argentina
S. chacoense, Cordoba, Argentina (AA-AD) X X X X
S chacoense, NW Argentina (CF, CI, DA) Y Y X
S. chacoense, Unknown (U, W, T) X x x X
S . kurtzianum, Unknown (AU, AX) X X X
S. simplicifolium, Unknown (CK) X X X
S. tarijense, Tarija (DC) X
S stenotomum, Cuzco (CU) X
S. stenotomum, Huanucc? (DM) X X
S stenotomurn, Charhapopas (CX) X X
S. phureja, Pichincha (AN) X X X
S. phureia, Narino (BV, BW, BZ) X X X
S . v e r r u c a " . D F. (DG', X x x
Bolivia Peru
Ecuador Colombia Mexico
X
X
Table 8 summarizes the plasmon characters of the biotypes analyzed with the plant sources arranged in south-north order. Four different plasmon types,
Of'-
In'-Plr-Lbr- Df'-in'-?-Lb', df'-in'-pls-lbr, and Dfr-?-pl'-Lb' are represented in the Table. The distinct pattern of geographic distribution of each of the individual plasmon characters indicates that each character is independent and separate from the others.
Two facts arising directly or indirectly from the data in Table 8 seem impor- tant in assaying the role of plasmon variation in the evolution of these plants: (1 ) It seems a surprising fact that four different plasmon characters directly related to male sterility have arisen in this plant group, while plasmon factors influencing other phenotypic characters have not been found. The lack of other phenotypic expressions has not been a result of a failure to search for them. Detailed observations of qualitative and quantitative features of flower, leaf. stem, and tuber characters of reciprocal F,'s and backcrosses were made. It is also evident that a large proportion of the cytoplasmic factors previously reported to have been found in other plant genera have been factors leading to male sterility. (2) The resistant form of three of the four plasmon characters,
D f ' , in?, and lb?' are distributed through a relatively wider range of species and a
wider geographic range than are their sensitive counterparts. The fourth char- acter, pollenless, has not yet been sufficiently mapped that any conclusions con- cerning its distribution can be drawn. The inference can be made that Of', in', and
P L A S M O N A N D GENE INTERACTION 1331 These two facts lead to the hypothesis that the important function of plasmon change under natural conditions may have been in the formation of isolating barriers to gene flow between populations. The change from the resistant to the sensitive plasmon would have resulted in a barrier to flow of genes from the parent population which contained the gene to which the new plasmon was sensitive into the population containing the newly arisen sensitive plasmon. The barrier presented by the recessive sensitive plasmons (dfs and pl“) taken alone would not have been a strong one since the sterility would have been expressed in only half of the pertinent backcross progeny. A dominant change such as that to Ins or Lb’ would obviously have been of much more intense and immediate effect. Thus, the S . chacoense population at Cordoba, Argentina, which has the Ins plasmon, is isolated from introgression of genes from Zn-bearing gametes. This difference between the chacoense at Cordoba and other chacoense populations could mark a first step in division of S. chacoense into two species.
If plasmon differentiation is a step in formation of isolating barriers, the impor- tant function of the plasmon characters may be expressed only under the unusual conditions when particular crosses have occurred. The inference can be drawn from this that the plasmon character is a highly stable one capable of maintaining itself over extensive areas and long periods of time even when it is not often ex- pressed. One may wonder whether the plasmon characters discussed here are self-reproducing permanent entities usually referred to as “cytoplasmic”
(CASPARI 1948) o r are a carry over from gene products of the original female parent, referred to as “maternal.” Recurrent backcrosses are usually used to draw this distinction, the character being considered cytoplasmic if it maintains itself after many backcross generations away from the genotype with which it orig- inally was iound. The limitation of this method is that leakage of cytoplasm from the pollen parent, as has been demonstrated, for example, in Epilobium
(MICHAELIS 1954) and Oenothera ( STINSON 1960), could cause a cytoplasmic variant to change during repeated backcrossing and act as though it were a mater- nal variant. It seems best to recognize that both possibilities exist in Solanum, though the df8 plasmon has maintained itself for three generations away from the original maternal genotype.
Finally. it should be mentioned that plasmon analysis may supply a tool of practical importance. Several different hypotheses have been advanced for the origin of the cultivated potato (SWAMINATHAN and HOWARD 1953; HOWARD 1960;
HAWKES 1956b; CORRELL 1952). By using appropriate crosses, especially using the diploid forms of S. tuberosum that have recently been obtained (HOUGAS, PELOQUIN and Ross 1958; PELOQUIN and HOUGAS 1960) it should be possible to classify the plasmon of this species and thereby gain information of a new sort concerning its evolutionary origin.
SUMMARY
1332 P. G R U N , M. A U B E R T I N A N D A. RADLOW
different unit characters of the same plasmon was developed: Plasmon df" pro- duces deformed flowers in the presence of genotype df df; Ins plasmon produces indehiscent flowers in the presence of one of three dominant genes I n , In', or
Znz; pLs plasmon leads to a pollenless condition in the presence of the p l p l geno-
type, and Lb' plasmon leads to lobed sterile microspores in the presence of the gene Lb. The plasmons Df', inr, PIr, and lb', by contrast, produce normal fertile flowers in interaction with either dominant or recessive alleles of the genes con- cerned. Among varying biotypes of 1 1 species, three of the sensitive (s) plasmons appear to have a relatively narrow range of distribution as compared with that of the corresponding resistant ( r ) plasmons. The data can be interpreted as show- ing that a change to the sensitive form of the plasmon is a n important step in formation of internal barriers to gene exchange between species in this genus.
A C K N O W L E D G M E N T S
We wish to express o u r sincere thanks to DR. R. W. HOUGAS and the U. S. Department of Agriculture for supplying the plant material used in this study.
DR. YATES BORDEN was of great assistance in programming use of I. B. _W. equip- ment for summarizing data. We are indebted to DR. HARLAN LEWIS for his critical review of the manuscript. The study was supported by a grant from the National Science Foundation.
L I T E R A T U R E C I T E D
BUCK, R. W., JR., 1960 Male sterility in interspecific hybrids of Solanum. J. Heredity 51 : 13-14.
CASPARI, E., 1948 CORRELL, D. S., 1952
Central America. U.S. Dept. Agr. Monograph 11. Washington, D.C.
EPHRUSSI, B., 1953 England. GRUN, P., 1961
diploid species of Solanum. Am. J. Botany 48: 79L89. GRUN, P., and A. RADLOW, 1961
self-compatible species of Solanum. Heredity 16 : 137-143. HAWKES, J. G., 1956a
and W. RUDORF. Handbuch der Pflanzenziichtung 3: 1-43. 37-109.
HIORTH, G., 1948
HOUGAS, R. W., S. J. PELOQUIN, and R. W. Ross, 1958 HOWARD, H. W., 1960
KOOPMANS, A., 1951
Cytoplasmic inheritance. Advan. Genet. 2 : 1-66.
Selection Tuberarium of the Genus Solanum of North America and Nucleo-cytoplasmic Relations in Micro-organisms. Clarendon Press. Oxford,
Early stages in the formation of internal barriers to gene exchange between
Evolution of barriers to crossing of self-incompatible with The potato. Taxonomy, cytology and crossability. Edited by H. KAPPERT, A revision of the tuber-bearing Solanums. Scot. Plant Breeding Sta. Ann. Rept. 1956:
'Uber Hemmungssysteme bei Godetia whitneyi. 11. Z. Ind. Abst. Vererb. 82:
1956b
276-330.
Haploids of the common potato. J. Heredity 49 : 103-1 06,.
Potato cytology and genetics. Bibl. Genet. 19: 87-216.
Cytogenetic studies on Solanum tuberosum L. and some of its relatives. Changes i n sex in the flowers of the hybrid Solanum rybinii x S . chacoense. Genetica Genetica 25: 193-337.
P L A S M O N A N D G E N E I N T E R A C T I O N 1333
1954 1955 1959
Changes i n sex in the flowers of the hybrid Solanum rybinii
x
S. chacoense. 11. Plas- matic influences upon gene action. Genetica 27: 273-285.Changes i n sex in the flowers of the hybrid Solanum rybinii x S. chmoense. 111. Data about the reciprocal cross Solanum chcoense x S. rybinii. Genetica 2 7 : 46511.71.
Changes in sex in the flowers of the hybrid Solanum rybinii x S. chacoense. IV. Further data from the reciprocal cross S . chacosnse
x
S. rybinii. Genetica 30: 384-390.LAVFN, H., 1959 Speciation by cytoplasmic isolation i n the Culex pipiens complex. Cold Spring Harbor Symp. Quant. Biol. 4: 166-173.
MICHAFLIS, P., 1954 Cytoplasmic inheritance i n Epilobium and its theoretical significance. Advan. Genet. 6: 287-4Q1.
OEHLKERS, F., 1938 Bastardierungsversuche in der Gattung Streptocarpus Lindl. I. Plasmatische Vererbung und die Geschlechtsbestimmung von Zwitterpflanzen. Z. Botan. 32 : 305-393. OEHLKERS, F., and U. EBELL, 1958 Bastardierungsversuche in der Gattung Streptocarpus Lindley. VII. Die genetischen Gmndlagen fur die Pollenste~rilitat verschiedener Strepto- carpus-bastarde. Z. Vererb. 89: 559-586.
PFLOQUIN, S. J., and R. W. HOUGAS, 1960 Genetic variations among haploids of the common potato. Am. Potato J. 37: 289-297.
STINSON, H. T., JR., 1960 Extranuclear barriers to interspecific hybridization between Oeno- thera hoakeri and Oenothera argillicola Genetics 45 : 81 9-838.
STUBBE, W., 1960 Untersuchungen zur genetischen Analyse des Plastoms von Oenothera. Z. Botan. 48: 191-218.