Copyright 0 1990 by the Genetics Society of America
s r d l , a Saccharomyces cerevisiae Suppressor of the Temperature-Sensitive
Pre-rRNA Processing Defect
of w p l - 1
Gary
R.Fabian,' Suzanne
M.
Nessand Anita K. Hopper
Department of Biological Chemistry and The Program in Cellular and Molecular Biology, The Milton S. Hershey Medical Center, The Pennsylvania State University, Hershey, Pennsylvania 17033
Manuscript received September 8, 1989 Accepted for publication November 9, 1989
ABSTRACT
We define a new gene, SRDl, involved in the processing of pre-rRNA to mature rRNA. The SRDl gene was identified by selecting for second-site suppressors of the previously described rrpl-1 mutation. T h e r r p l - 1 mutation causes temperature-sensitive growth, a conditional defect in processing of 27s pre-rRNA to mature 25s rRNA, and a nonconditional increase in sensitivity to several aminoglycoside antibiotics. All s r d l alleles identified are recessive and apparently specific to the rrpl- I mutation. Although a mutation of SRDI suppresses the pre-rRNA processing defect, drug sensitivity and thermolethality of a point mutation of RRPl, it is unable to suppress a rrpl-disruption allele. We
suggest that the SRDl gene product either interacts with or regulates the RRPI product.
T
HE processing of precursor ribosomal RNA (pre- rRNA) species to mature rRNAs is complex and has been the subject of several review articles (HAD- JIOLOV 1985; HADJIOLOV and NIKOLAEV 1976; No-MURA, GOURSE and BAUCHMAN 1984; WARNER 1989).
There are multiple steps to this process including several nucleolytic cleavages and the participation of many proteins is required. In Saccharomyces cerevisiae, an initial 35s pre-rRNA is processed to 27s pre-rRNA and 20s pre-rRNA species. T h e
27s
pre-rRNA, in turn, is processed to 25s and 5.8s mature rRNAs and the 20s pre-rRNA to the mature 1 8 s species. This process requires the presence of proteins found on mature ribosomes (review: WARNER 1989) and small nuclear RNAs (TOLLERVEY 1987; LI, ZAGORSKI and FOURNIER, 1990). Furthermore, many nucleoside modifications are also known to occur before proper processing may proceed (review: WARNER 1989).Although much information about the eukaryotic processing pathway exists, to date not a single proc- essing enzyme has been identified nor is it known whether processing steps can be autocatalytic as is the case with rRNA splicing in Tetrahymena (review: CECH and BASS 1986). Our approach to studying pre-rRNA processing has been to identify mutants of S. cerevisiae
that are defective in rRNA processing and then to conduct a molecular analysis of the wild-type counter- part of the gene identified by mutation.
Previously we reported the characterization of the RRPl locus. A mutation of this locus, r r p l - I , simul-
nia.
' Present address: 350 Cambridge Avenue, Suite 100, Palo Alto, Califor-
of page charges. This article must therefore be hereby marked "advertisement"
T h e publication costs of this article were partly defrayed by the payment
in accordance with 18 U.S.C. $1734 solely t o indicate this fact. Genetics 1 2 4 497-504 (March, 1990)
taneously causes conditional lethality, hypersensitivity to aminoglycoside antibiotics, and abolishes the proc- essing of 27s pre-rRNA to the mature 25s and 5.8s rRNAs of the 6 0 s subunit at the nonpermissive tem- perature (ANDREW, HOPPER and HALL 1976; FABIAN and HOPPER 1987). A hypothesis that would have explained the role of RRPl in pre-rRNA processing is that RRPl could code for a component of the mature 60s subunit that assembled onto the RNA in the nucleus during pre-rRNA processing. T h e RRPl gene has been cloned and it was shown that the RRPl mRNA is in lower abundance than are the mRNAs for other ribosomal proteins (FABIAN and HOPPER
1987). These results indicated that the RRPl gene product probably does not code for a ribosomal pro- tein. T h e RRPI gene has been sequenced (L. LEMPER-
EUR and R. J. CROUCH, personal communication) but
an analysis of the ORF did not reveal the function of this gene product.
An alternative approach to gain an understanding of the role of RRPl in yeast pre-rRNA processing is to determine with what other cellular components the RRPl protein interacts. In pioneering studies, JARVIK
and BOTSTEIN (1973, 1975) showed that extragenic second-site suppressors can identify genes whose prod- ucts interact with the defective product of the first gene. We have used this strategy to identify genes encoding products that interact with the RRPl pro- tein. In this study we report mutations of a locus, SRDI, that can suppress the phenotypes of r r p l - I , but not a disruption of the RRPl locus.
MATERIALS AND METHODS
498 G. R. Fabian, S. M. Hess and A. K. Hopper
TABLE 1
Yeast strains
Strain name Genotype Source
GRF123 a trpl adel r r p l - 1 lys2gall FABIAN and HOPPER (1 987) aga17:4b a canl-100 lys2-1 gal7 leu- trp- ade2 This laboratory
G1/7-5-13a a his7 leul ade2 trpl lys2 tyrl ga17ga12 From GRF 123 X aga17:4b C1/7-5-13b a ade2 trpl lys2 tyrl gall gal2 From GRF 123 X aga17:4b G1/7-5-13c a leul ade2 trpl lys2 tyrl ga17ga12 r r p l - 1 From GRF 123 X aga17:4b G1/7-5-13d a his7 ade2 trpl lys2 tyrl gall gal2 r r p l - 1 From GRF 123 X aga17:4b
G4 a his7 ade2 t r p l lys2 tyrl gall gal2 srdl-2 r r p l - 1 Original isolate of s r d l - 2 in G1/7-5-13d
Y5 a leul ade2 trpl lys2 tyrl ga17ga12 srdl-1 rrpl-1 Original isolate of srdl-1 in G1/7-5-13c
Y 8 a leul ade2 trpl lys2 tyrl ga17ga12 srdl-4 r r p l - 1 Original isolate of srdl-4 in G1/7-5-13c
Y11 a leul ade2 t r p l lys2 tyrl ga17ga12 srdl-3 rrpl-I Original isolate of s r d l - 3 in G1/7-5-13c G1/7-6-7c a srdl-1 r r p l - 1 gal7 leul tyrl ade2 trpl his7 lys2ga12 Y5 X G1/7-5-13d
G1/7-6-3b a srdl-1 rrpl-1 gall tyrl ade2 trpl his7 lys2ga12 Y5 X G1/7-5-13d G1/7-7-5d a s r d l - 2 r r p l - l gal7 tyrl ade2 trpl his7 lys2ga12 G4 x G1/7-5-13c G1/7-7-lb a s r d l - 2 rrpl-1 gall tyrl ade2 t r p l his7 lys2ga12 G4 X G1/7-5-13~ 5d X I b MATaIMATa, his7/his7, ade2/ade2, t r p l l t r p l , lys2/lys2, t y r l l t y r l , This study
MAPl6D a r r p l - 1 s r d l - 2 This study
MAP15D a r r p l - 1 his1 leu2-3, 112 This study gal2/gal2, gall/+, gal7/+, srdl-2/srd1-2, rrpl-l/rrpl-l
and dropout media were prepared as previously described (HOPPER, BANKS and EVANGELIDIS 1978). YEPD
+
neomy- cin was YEPD plus 2 mg/ml Neomycin B (Sigma).Selection of diploid yeast strains was accomplished pri- marily by selection of strains able to ferment galactose on BTB-gal indicator plates (1 0% galactose, 1 % Bacto-peptone, 1% yeast extract, 0.04% adenine, 0.04% uracil, 2% agar and 10 ml/liter saturated bromothymol blue solution). Standard methods for sporulation, ascus dissection and tetrad analysis (MORTIMER and HAWTHORNE 1969) and the random spore method of DAVIDOW and BYERS (1 984) were used.
Selection of second-site revertants: We selected for var- iants of rrpl-1 strains able to grow at elevated temperatures or able to grow in the presence of neomycin. Two strains (G1/7-5-13c and G1/7-5-13d; Table 1) of opposite mating types each carrying the rrpl-1 mutation and either g a l l or gal7 were streaked for single colonies. Typically, 20 clones of each mating type were individually inoculated into 2 ml of YEPD broth and grown 2 days at 23" with aeration. Cultures were plated to YEPD at approximately 4 X 10" cells per plate and to YEPD
+
neomycin at 5 X lo' cells per plate. The YEPD plates were incubated at 37" for 2-3 days and the YEPD+
neomycin plates at 30" for 5 days. One colony from each clone was picked and streaked for single colonies.Analysis of RNA: RNA was prepared as previously de- scribed (FABIAN and HOPPER 1987). RNAs were resolved on 1.5% agarose formaldehyde gels as described by MAN-
IATIS, FRITSCH and SAMBROOK (1982) except that we em-
ployed 0.01 M NaP04 (pH 6.5), 10 mM EDTA for the electrophoresis buffer. Following electrophoresis the gels were rinsed with four changes of electrophoresis buffer minus formaldehyde and then transferred to autofluor (Na- tional Diagnostics) and gently shaken for 3 hr. The imbed- ded gels were dried under vacuum and then exposed to XAR-5 X-ray film at -70".
Exposures (6 hr, 16 hr, 20 hr) of the autoradiograms were scanned at 500 nm using a Beckman DU8 Spectropho- tometer and the Beckman DU8 Gel Scanning Program. The absorbances were plotted and peaks representing the 25s and 18s bands were identified. Background was subtracted, the area under the peak calculated for each lane, and the
ratio of the areas for the 25s and 18s rRNAs was deter- mined.
DNA manipulations: Restriction enzymes were obtained from a variety of manufacturers and digestions with these enzymes were per the manufacturer's instructions. Specific DNA fragments derived from restriction enzyme-digested plasmids were purified by utilization of an Elutrap (Schleicher 8c Schuell). T 4 DNA ligase was obtained from Bethesda Research Laboratories and was used according to the manufacturer's instructions. Analysis of yeast genomic DNA by Southern (1975) analysis was as previously reported (ATKINSON, DUNST and HOPPER 1985).
Generation of a disruption allele of R R P I : Figure l a shows the construction of plasmid pUC4T. The PstI, frag- ment from pMac56 1 (MCKNIGHT and MCCONAUGHY 1983) containing the yeast TRPl gene was inserted into pUC4K (VIEIRA and MESSING 1982) that had been digested with PstI. The pUC4T vector has the TRPl gene flanked by paired PstI, SalI, BamHI and EcoRI sites. Figure l b shows the construction of pSH1. T h e 4.3-kb ClaI/BamHI DNA fragment from YEpRRP1-2 (FABIAN and HOPPER 1987) was inserted into the AccI/BamHI sites of the polylinker region of pUCI9. pSHl contains the entire RRPl sequence and a unique SalI site within a region necessary for complemen- tation of the rrpl-1 mutation (FABIAN and HOPPER 1987). Fig. I C shows the restriction map of pSH2. T o generate pSH2, pUC4T was digested with Sal1 and the T R P l frag- ment was purified and ligated into the Sal1 site of pSH1. This resulted in placement of the TRPI gene within an essential region of R R P l . T o generate a genomic disruption of R R P l , pSH2 was linearized with BarnHI and used to generate a gene replacement (ORR-WEAVER, SZOSTAK and ROTHSTEIN 1983) of a srdl-2 rrpl-1 trpllsrdl-2 rrpl-1 trpl diploid strain (5d X lb, Table 1 ) .
RESULTS
Genetic characterization of S R D l : Variants of pa- rental strains G1/7-5-13c, and G1/7-5-13d (Table l ) carrying the rrpl-1 mutation were selected for t h e ability to grow at 37" or to grow in the presence of neomycin as described in MATERIALS AND METHODS.
Suppressor of rRNA Processing Defect
P R P
pMac561
499
a.
b.
B P P B
c"GGA TCC GTC GAC CTG CAG GGG
p V C 4 K I I I p U C 4 K
CCC CTG CAG GTC GAC GGA TCC
-
I I
8
E C R E E G B
CCA AGC TTG CAT GCC TGC AGG TCG ACT CTA GAG GAT CCC GGG GTA CCG AGC TCG AAT T C A T
p U C l 9
I
1,
1
'
I
Hind I11
I
ma I Sma I 'pn I Eco ISph I Xma I Sst 1
Acc
I
BamHIS a l I
Hfnc I1
R R S P R P S E E G B
C .
.
. 5 kb
FIGURE 1 .-Construction of a disruption allele of R R P I . Plasmid pSH2 was generated by a three-step procedure as detailed in MATERIALS A N D METHODS. a , Construction of pUC4T; b, Construction of pSH1; c, Restriction map of pSH2. T h e Sal1 fragment from pUC4T (see highlighted S) containing the yeast TRPI gene was ligated into the unique SalI site (see highlighted S) of pSHl located within an essential region of RRP1. Symbols and abbreviations: 0, TRPI sequences; W, kanamycin resistance gene sequences from pUC4K; M, RRPI sequences from YEpRRPI-2; P, PstI; R, EcoRV; B, BamHI; E, EcoRI; C , ClaI; S, S a l I ; G , BglII. DNA from part of the polylinker region of pUC4K is shown i n a. DNA from the polylinker region of pUCI9 is shown in b. Note: DNA sequences between Sal1 and PstI shown as a part of the
TRPI gene are actually part of the pUC4K polylinker sequence. T h e ClaI site is destroyed upon construction of pSH 1.
analyses. The reversion frequency to temperature- resistance (ie., able to grow at 37 ") was approximately 1.2 X 1 0-' and the reversion frequency to neomycin resistance (growth on YEPD
+
neomycin at 30") was approximately 6.5 X T h e apparent high rever- sion frequency to neomycin resistance may be an artifact of the selection since rrpl-1 strains divide before their growth is arrested by neomycin.T h e variants that were able to grow on media containing neomycin were also able to grow at 37 O .
Similarly, cells selected by restoration of growth at 3 7 " were able to grow in the presence of neomycin. No phenotypic differences among the suppressors was observed regardless of the nature of their selection.
T h e revertants obtained from the selections were mated to rrpl-1-bearing strains. We employed the tightly linked noncomplementing gall and gal7 mu- tations to select for diploid cells and to assess haploidy in random-spore analysis. Therefore, gall rrpl-1 or
500 G. R. Fabian, S. M. Hess and A . K . Hopper
TABLE 2
Analysis of revertants of rrpl-1
No. of alleles Suppressor segregating phenotype independent of
rrpl-1
Selective Strain Condition
Domi- Reces- Domi- Reces- nant sive nant sive
G1/7-5-13~ YEPD/37" 44 15 0/44 15/15 G1/7-5-13d 47 13 0/47 13/13
G1/7-5-13c YEPD
+
neo/ 34 1 1 0/34 1 1 / 1 1 30"G1/7-5- 13d
" 35 10 0/35 10/10
160 49 0/160 49/49
gall
r r p l - 1 strains (G1/7-5-13c and G1/7-5-13d, Table l), respectively, and diploids were selected on BTB-gal medium. T h e diploids were tested for the ability to grow under the selective condition used to select the original revertant, thus testing the dominant or recessive nature of the lesion.A total of 49 recessive and 160 dominant revertants were isolated. The revertants were then crossed to wild-type
gal7
RRPl orgall
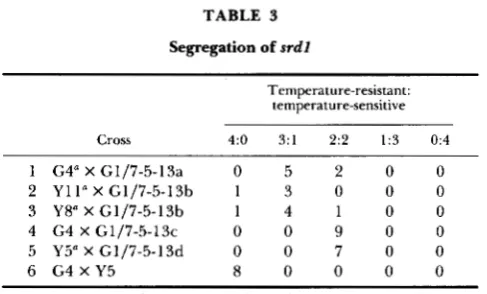
RRPl strains (G1/7-5-13a and G1/7-5-13b, Table l), diploids were selected and sporulated, and the Gal- progeny analyzed by random spore analysis. If the suppressor were a sec- ond-site suppressor and not closely linked to the RRPl locus, then temperature-sensitive or neomycin-sensi- tive progeny would be obtained when rrpl-1 and the suppressor segregated to separate meiotic progeny. All of the recessive suppressors were shown to be unlinked to r r p l - I . None of the dominant suppressors segregated independently of RRPl (data summarized in Table 2) and therefore were most likely intragenic revertants at the RRPl locus or suppressors that re- sulted in lethality in the absence of the rrpl-1 muta- tion. The extragenic location of the suppression of three isolates (G4, Y 1 1 and Y8) was verified by tetrad analysis (Table 3, crosses 1,
2
and 3). If the suppressor were intragenic or tightly linked to the RRPl locus, then 4:O (temperature-resistant:temperature-sensi-tive) segregation would have been expected. How- ever, in each case the suppressor segregated inde- pendently of RRPl.
In order to categorize the mutations into comple- mentation groups, the mutant derivatives of G1/7-5- 13c were mated to mutant derivatives of the MATa
r r p l - 1 strains ( i e . , G1/7-5-13d) and diploids were obtained by replica-plating to selective plates. If the recessive suppressors were unable to complement, the diploids should grow under selective conditions (37" or in the presence of neomycin). By this analysis only one complementation group was identified. Further, MATa and MATa progeny of the crosses G4 X G1/7- 5-13c and Y5 X G1/7-5-13d (see below) were also used for complementation tests and by this analysis all
TABLE 3
Segregation of srdl
Temperature-resistant: temperature-sensitive
Cross 4:O 3:l 2 2 1:3 0:4
1 G4" X G1/7-5-13a 0 5 2 0 0
2 Y11" X G1/7-5-13b 1 3 0 0 0
4 G4 X G1/7-5-13c 0 0 9 0 0
5 Y5" X G1/7-5-13d 0 0 7 0 0
6 G4 X Y5 8 0 0 0 0
G4, Y5, Y8 and Y11 carry independent suppressors of r r p l - 1 .
3 Y8" X G1/7-5-13b 1 4 1 0 0
49 suppressors belong to a single complementation group. We have named this complementation group SRDl for suppressor of rRNA processing defect.
T w o of the isolates, G4 (an isolate of G1/7-5-13d) and Y5 (an isolate of G1/7-5-13c) were crossed to
rrpl-1 bearing strains to determine if a single gene was responsible for the suppression phenotype (Table 3, crosses 4 and 5). In each case, there was
2:2
segre- gation of temperature-resistant:temperature-sensitiveprogeny, indicating that the suppressors behaved as single Mendelian genes.
T o confirm that the suppressors in Y5 and G4 were indeed at the same genetic locus, the progeny of a Y5
X G4 diploid were analyzed. If the suppressors were at genetically unlinked loci, temperature-sensitive progeny would be expected to segregate. If the sup- pressors were at the same genetic locus, all progeny were expected to be temperature resistant. T h e latter result was obtained; 8/8 tetrads segregated 4:O for temperature-resistance to temperature-sensitivity. This indicated that srdl-1 from Y5 and srdl-2 from G4, are at or near the same genetic locus (Table 3, cross 6).
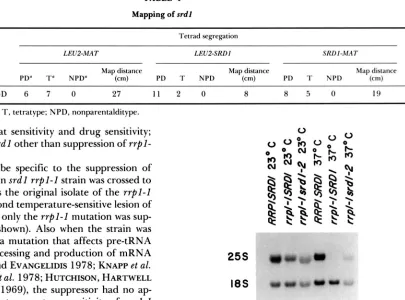
Inspection of the segregation of markers in crosses 4 and 5 (Table 3), indicated that SRDl is centromere linked, possibly to chromosome I11 ( i e . , there were no NPD tetrads observed for MAT-srdl segregation, data not shown). T h e assignment to chromosome I11 was confirmed by the analysis of progeny from the diploid formed by the mating of MAPl6D (relevant genotype: MATa srdl LEU2 r r p l - I ) to MAP15D (rel- evant genotype: MATa SRDl leu2 r r p l - I ) . This analy- sis placed SRDl near the centromere of chromosome 111 (Table 4). Random spore analysis of the segrega- tion of the three linked genes, MAT, LEU2 and SRDI, showed that SRDl is located on the right arm of chromosome 111 between the centromere and MAT.
The srdl r r p l - 1 strains were tested for cold sensi- tivity, heat sensitivity and drug sensitivity/resistance (aminoglycosides, trichodermin, crytopleurine and cy- cloheximide), but no differences between srdl rrpl-1
Suppressor of rRNA Processing Defect 50 1
TABLE 4
Mapping of srdl
Tetrad segregation
LEU2-MAT LEU2-SRDI SRDI-MAT
Cross PD" T" NPD" Map distance (cm) PD T NPD Map distance (cm) PD T NPD Map distance (cm)
MAPIGDX MAP15D 6 7 0 27 1 1 2 0 8 8 5 0 19
a PD, parental ditype; T, tetratype; NPD, nonparentalditype.
3 (Table 3) for heat sensitivity and drug sensitivity; no phenotype for srdl other than suppression of rrpl- 1 was found.
srdl apepars to be specific to the suppression of rrpl-1 since when an srdl rrpl-1 strain was crossed to ts351 which carries the original isolate of the rrpl-1
mutation and a second temperature-sensitive lesion of unknown function, only the rrpl-1 mutation was sup- pressed (data not shown). Also when the strain was crossed to rnal-1, a mutation that affects pre-tRNA and pre-rRNA processing and production of mRNA (HOPPER, BANKS and EVANGELIDIS 1978; KNAPP et al.
1978; O'FARRELL et al. 1978; HUTCHISON, HARTWELL and MCLAUCHLIN 1969), the suppressor had no ap- parent effect on the temperature sensitivity of rnal-1 (data not shown). Furthermore, srdl showed no evi- dence of suppressing any missense or nonsense auxo- trophic markers present in our strains (Table 1).
The effect of srdZ on rRNA processing: The in-
corporation of ['Hluracil into mature rRNAs was studied to determine whether srdl corrected the pre- r R N A processing defect as well as the temperature- sensitivity and drug sensitivity defects of rrpl-1 cells. Three related strains were employed: G1/7-5-13a (rel- evant genotype, R R P l SRDl), G1/7-5-13c (relevant genotype, rrpl-1 SRDl) and G1/7-7-5d (relevant gen- otype, rrpl-1 srdl-2) (Table 1). The ability of each strain to generate 25s mature rRNA at permissive and nonpermissive temperatures was determined (Fig- ure 2). As expected, the ratio of radioactivity in 25s: 18s rRNAs for wild-type cells at both tempera- tures was about 2. As previously reported (FABIAN and HOPPER 1987), this ratio is aberrant in rrpl-1 cells. At permissive temperatures, the ratio of radio- active 25s to 18s rRNA is about 1. At the nonper- missive temperature, no mature 25s rRNA is gener- ated. Interestingly, the ratio of 25s to 18s RNA in the rrpl-1 srdl-2 cells at the permissive temperature is not a 2: 1 ratio as seen with wild-type cells. The ratio (1.3:l) is similar to what is seen in the rrpl-1 cells at permissive temperature. A t non-permissive tempera- ture, the ratio is less (about 0.6:l) indicating that srdl-
2 causes a partial, but not a complete suppression of the rrpl-1 processing defect at 37".
A mutation of SRDZ fails to suppress a disruption
25s
18s
FIGURE 2.-Pre-rRNA processing in rrpl-1 srdl-2 cells. RNAs of strains G1/7-5-13a (wt), G1/7-5-13c (rrpl-I), and G1/7-7-5d
(rrpl-I srdl-2) were pulse-labeled for 1 hr with ['H]uracil 2 0 min after a shift to 37" and then chased for 30 min with an excess of unlabeled uracil. RNA from strains grown at both 23" and 37" was isolated and approximately 1 X IO6 cpm of RNA from each sample was resolved on a 1.5% agarose formaldehyde gel. Lanes containing RNA from rrpl-1 SRDl and rrpl-1 srdl-2 cells incubated at 37" have apparently less radioactivity incorporated into 2 5 s and 18s
rRNA probably because mRNAs, tRNA and 5s RNA comprise a large percentage of total RNAs in these cells.
allele of RRPZ: Three mechanisms by which muta-
tions of SRDl might suppress the rrpl-1 lesion are: (1) SRDl might code for a product that interacts with the RRPl protein, (2) SRDl might regulate the level of R R P l product and (3) mutations of SRDI could cause a bypass of the RRPl-mediated pre-rRNA proc- essing pathway. Mechanisms 1 and 2 predict that the srdl lesion could not suppress a rrpl-null allele, whereas mechanism 3 predicts that srdl would sup- press a rrpl-null allele. T o distinguish model 3 from
1 or 2, we constructed a rrpldisruption allele in a rrpl-1 srdl-2 trpl/rrpl-l srdl-2 trpl diploid strain, (5d X lb, Table 1) by a gene replacement (MATERIALS
AND METHODS). The diploid strain was transformed with plasmid pSH2 (Figure IC) linearized with BamHI and Trp+ prototrophs were selected.
502 G. R. Fabian, S. M. Hess and A. K. Hopper
kb
W X I5.6
4.6
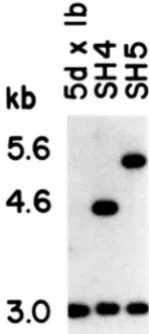
FIGURE 3.-Confirmation of rrpl-disrupted allele. Genomic DNA from 5d X Ib, the parental strain, and two potential
rrpl::TRPl strains, SH4 and SH5, was analyzed by Southern analy- sis. DNA was digested with EcoRI, resolved on agarose gels, trans- ferred to membranes and hybridized with a nick-translated '*P- labeled gel purified 3-kb EcoRl fragment from YEpRRPl-2.
to Gene Screen Plus (DuPont Co.). A 3-kb gel purified EcoRI fragment (Figure lb) from YEpRRPl-2 (FA-
BIAN and HOPPER 1987) was nick translated with ['*PI -dTTP and employed as a probe. Three hybridization patterns were detected: two were unique (Figure 3, lanes 2 and 3) and the third was a composite of the first two (not shown). If a disruption of RRPl had been obtained, we would have expected two bands, the 3.0-kb parental band and a 4.6-kb band resulting from the insertion of TRPl in the RRPl coding re- gion. If an integration of the rrp1::TRPl and the vector sequences at the RRPl locus had been ob- tained, we would have expected the 3.0-kb parental band and a -6.8-kb novel band. The pattern seen for SH4 (Figure 3, lane 2) meets the criteria for a gene replacement; the pattern for SH5 (Figure 3, lane 3) is not consistent with either a gene replacement or an integration event at R R P l , but could be due to an integration at TRP1. Subsequent digestions with EcoRV and BglII (not shown) confirmed that SH4 contained the rrp1::TRPl gene replacement. AI- though the origin of the rearrangement in SH5 was not pursued, we have used SH5 as a control for our genetic analysis (see below).
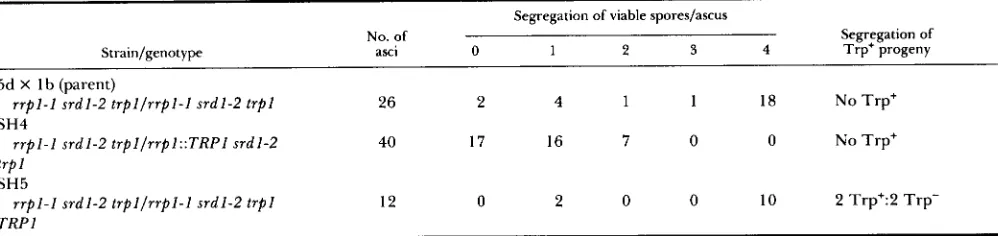
The undisrupted parent, SH4 and SH5 were spor- ulated and the progeny were dissected. Of 26 asci from 5d X lb, 18 yielded 4:O segregation for viable spores; all progeny were temperature resistant (Table 5). SH5 generated very similar viability and the asci segregated 2:2 for Trp+:Trp-. SH4, on the other hand, yielded only 0, 1 or 2 viable spores/ascus and no Trp+ segregants were obtained. All spores ob- tained from SH5 and SH4 were temperature resistant. Therefore SH4 possesses a recessive-lethal lesion linked to TRPI. These results indicate that srdl-2 is unable to suppress a disruption allele of R R P l .
DISCUSSION
The purpose of selecting extragenic second-site sup- pressors of rrpl-1 was two-fold: to learn more about the RRPl gene and its product and to identify other genes encoding products involved in pre-rRNA proc- essing. A single locus identified as being able to give
rise to recessive suppressors of rrpl-1 was named SRDl (suppressor of rRNA processing defect). Al- though our analysis is not complete, it would appear that 49 independent isolates of SRDl were obtained. Thus it seems unlikely that any other gene could be identified by searching for additional second-site s u p pressors of rrpl-1. No dominant second-site suppres- sors of rrpl-1 were isolated. Independent of whether the selection was for suppression of neomycin-sensitiv- ity or temperature-sensitivity of r r p l - I , all isolates were capable of suppressing either phenotype. srdl appears to be specific for the suppression of rrpl-1.
No independent phenotype of the srdl mutations (such as cold-sensitivity, temperature-sensitivity, drug resistance/sensitivity) has been identified; however, it was seen that for a rrpl-1 srdl-2 strain the ratio of 25s to 18s rRNAs was not 2: 1 as seen for wild-type cells. SRDl was shown to be centromere linked and located on the right arm of chromosome 111.
Although we have not yet learned its function, we have learned more about R R P l . Our earlier studies showed that the rrpl-1 lesion either caused hypersen- sitivity to aminoglycoside antibiotics or was tightly linked to another mutation that caused this sensitivity [i.e., no rrpl-1 segregants that were resistant to amino- glycosides were obtained (FABIAN and HOPPER
1987)]. The fact that both putative revertants at RRPl and extragenic lesions at SRDl simultaneously correct the temperature-sensitivity and the antibiotic sensitiv- ities strongly suggest that a single lesion at RRPl causes both phenotypes. Whereas there are several models to explain this phenomenon, we favor one that, according to the conventional means by which aminoglycoside antibiotics affect ribosomes, would im- plicate the RRPl gene product in the structure of ribosomes.
Previously only a single lesion of RRPl, rrpl-1, existed. This allele causes an inability of cells to grow at elevated temperatures. The interpretation of this phenotype was that RRPl is an essential gene. How- ever, this phenotype is also consistent with the models that RRPl encodes a protein needed only at high temperatures or that the rrpl-1 product titrates out an independent essential gene product. The fact that a rrpl-1 srdl-2/rrpl::TRPl srdl-2 diploid cell yields no more than two viable spores per ascus suggests that RRPl is an essential gene. We do not understand why
Suppressor of r R N A Processing Defect 503
TABLE 5
Analysis of the ability of srdl-2 to suppress wp1::TRPl
Segregation of viable spores/ascus
No. of Segregation of
Strain/genotype asci 0 1 2 3 4 Trp+ progeny
5d X I b (parent)
SH4
r r pl-I s r d l- 2 t r p l / r r p l- l sr d l- 2 t rp l 26 2 4 1 1 18 No Trp+
r r pl -l s r d l - 2 tr p l /r r p l: :TR PI srd l - 2 40 17 16 7 0 0 No Trp+
trp 1
SH5
r r p l - l t r pl / r r p l -l s r d l- 2 tr pl s r dl - 2 12 0 2 0 0 10 2 Trp+:2 Trp- TRP 1
independently derived disruptions of the 5d X l b parental strain.
In order to begin to understand how mutations of SRDI suppress r r p l - I , we sought to determine whether s r d l - 2 could suppress a disruption allele of the RRPl locus. We found that srdl-2 is able to suppress a nitrosoguanidine-induced mutation of the RRPl locus, but not a disruption allele. This suggests that a mutation of SRDl does not bypass the necessity for a functional RRPI gene product. This result leads us to consider two general classes of models for SRDl function: (1) SRDl encodes a product that interacts with the RRPl protein and (2) SRDl regulates the level of RRPI protein. There are several possible specific functions for SRDl in the first model. One possible explanation of s r d l suppression of rrpl-1 is that the conformational change of s r d l corrects an aberrant scaffolding of the pre-rRNA caused by a mutant rrpl protein. Alternatively, SRDl and RRPl may code for components of a multimeric enzyme complex involved in rRNA processing as has been demonstrated in Bacillus subtilis (STAHL et al. 1984; PACE, STAHL and PACE 1984). In these two cases, we would account for the recessiveness of srdl mutations by assuming that the product of the wild-type SRDl gene interacts more strongly, but ineffectively, with the rrpl-1 gene product than does the product of the mutant s r d l locus. Under the second class of models there are also several possibilities. For example, SRDl could code for a negative regulator of RRPl transcrip- tion. In this case we would assume that rrpl-I pro- duces a product that is less active than the RRPl protein. Alteration of the regulatory component would then result in greater production of the rrpl-1
protein. Alternatively, regulation could be mediated at a posttranscriptional step. For example, rrpl-1
could encode a pre-RNA processing activity that is labile at non-permissive temperatures. In this case, SRDl could encode, for example, a protease that is inactive or less active in s r d l cells. T h e isolation of the SRDl gene will provide information on another product involved in pre-rRNA processing and should
ultimately help elucidate the function of RRPI. T h e task of identifying a clone encoding SRDl should not be difficult since the region of chromosome 111 where SRDl maps has been thoroughly studied (NEWLON et al. 1986).
This work was supported by a U.S. Public Health Service grant from the National Institutes of Health to A.K.H. We thank S. WANG and H. TRACLIA for valuable comments on the manuscript and F. CRAMER for preparation of the manuscript.
LITERATURE CITED
ANDREW, C., A. K. HOPPER and B. D. HALL, 1976 A yeast mutant defective in the processing of 2 7 s r-RNA precursor. Mol. Gen. Genet. 144: 29-37.
ATKINSON, N. S., R. W. DUNST and A. K. HOPPER, 1985 Characterization of an essential Saccharomyces cerevisiae gene related to RNA processing: cloning of RNA1 and generation of a new allele with a novel phenotype. Mol. Cell. Biol. 5: 907- 915.
CECH, T . R., and B. L. BASS, 1986 Biological catalysis by RNA.
DAVIDOW, L. S., and B. BYERS, 1984 Enhanced gene conversion and post meiotic segregation in pachytene-arrested Saccharo- myces cerevisiae. Genetics 106: 165-183.
FABIAN, G. R., and A. K. HOPPER, 1987 R R P I , A Saccharomyces cerevisiae gene affecting rRNA processing and production of mature ribosomal subunits. J. Bacteriol. 169: 1571-1578. HADJIOLOV, A. A., 1985 T h e nucleolus and ribosome biogenesis.
Cell Biol. Monogr. 12: 1-268.
HADJIOLOV, A. A., and N. NIKOLAEV, 1976 Maturation of ribo- somal ribonucleic acids and the biogenesis of ribosomes. Progr. Biophys. Mol. Biol. 31: 95-144.
HOPPER, A. K., F. BANKS and V. EVANGELIDIS, 1978 A yeast mutant which accumulates precursor tRNAs. Cell 14: 21 1- 219.
HUTCHINSON, H. T . , L. H. HARTWELL and C. S. MCLAUGHLIN, 1969 A temperature sensitive yeast mutant defective in ri- bonucleic acid production. J. Bacteriol. 9 9 807-814.
JARVIK, J., and D. BOTSTEIN, 1973 A genetic method for deter- mining the order of events in a biological pathway. Proc. Natl. Acad. Sci. USA 7 0 2046-2050.
JARVIK, J.. and D. BOTSTEIN, 1975 Conditional-lethal mutations that suppress genetic defects in morphogenesis by altering structural proteins. Proc. Natl. Acad. Sci. USA 72: 2738-2742. KNAPP, G., J. S. BECKMANN, P. F. JOHNSON, S. A. FUHRMAN and J.
ABELSON, 1978 Transcription and processing of intervening sequences in yeast tRNA genes. Cell 14: 221-236.
504 G. R. Fabian, S. M. Hess and A. K. Hopper essential snRNA disrupts processing of 18s ribosomal RNA in
Saccharomyces cerevisiae. Mol. Cell. Biol. 10 (in press).
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular
Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MCKNIGHT, G . L., and B. L. MCCONAUGHY, 1983 Selection of functional cDNAs by complementation in yeast. Proc. Natl. Acad. Sci. USA 8 0 4412-4416.
MORTIMER, R. K., and D. C. HAWTHORNE, 1969 Yeast genetics, pp. 385-460 in The Yeasts, edited by A. H. ROSE and J. S. HARRISON. Academic Press, New York.
NEWLON, C. S., R. P. GREEN, K. J. HARDEMAN, K. E. KIM, L. R. LIPCHITZ, T . G. PALZKILL, S. SYNN and S. T. WOODY, 1986 Structure and organization of yeast chromosome 111, pp. 21 1-223 in Yeast Cell Biology, edited by J. HICKS. Alan R. Liss, New York
NOMURA, M., R. GOURSE and G. BAUGHMAN, 1984 Regulation of the synthesis of ribosomes and ribosomal components. Annu. Rev. Biochem. 53: 75-1 17.
O'FARRELL, P., B. CORDELL, P. VALENZUELA, W. RUTTER and H. GOODMAN, 1978 Structure and processing of yeast precursor rRNAs containing intervening sequences. Nature 274: 438- 445.
ORR-WEAVER, T. L., J. W. SZOSTAK and R. J. ROTHSTEIN, 1983 Genetic applications of yeast transformation with linear and gapped plasmids. Methods Enzymol. 101: 228-245. PACE, B., D. A. STAHLand N. R. PACE, 1984 The catalytic element
of a ribosomal RNA-processing complex. J. Biol. Chem. 259:
11454-1 1458.
SOUTHERN, E. M., 1975 Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98: 503-5 17.
STAHL, D. A., B. PACE, T. MARSH and N. R PACE, 1984 T h e ribonucleoprotein substrate for a ribosomal RNA-processing nuclease. J. Biol. Chem. 2 5 9 11448-1 1453
TOLLERVEY, D., 1987 A yeast small nuclear RNA is required for normal processing of pre-ribosomal RNA. EMBO J. 6: 4169- 4175.
VIEIRA, J., and J. MESSING, 1982 The pUC plasmids, a M I 3 mp7 derived system for insertion mutagenesis and sequencing with a synthetic universal primer. Gene 1 9 259-268.
WARNER, J. R., 1989 Synthesis of ribosomes in Saccharomyces cerevisiae. Microbiol. Rev. 53: 256-271.