DOI: 10.1534/genetics.110.121194
A Two-Pathway Analysis of Meiotic Crossing Over and Gene
Conversion in
Saccharomyces cerevisiae
Franklin W. Stahl
1and Henriette M. Foss
Institute of Molecular Biology, University of Oregon, Eugene, Oregon 97403-1229
Manuscript received February 17, 2010 Accepted for publication July 31, 2010
ABSTRACT
Several apparently paradoxical observations regarding meiotic crossing over and gene conversion are readily resolved in a framework that recognizes the existence of two recombination pathways that dif-fer in mismatch repair, structures of intermediates, crossover interdif-ference, and the generation of non-crossovers. One manifestation of these differences is that simultaneous gene conversion on both sides of a recombination-initiating DNA double-strand break (‘‘two-sidedness’’) characterizes only one of the two pathways and is promoted by mismatch repair. Data from previous work are analyzed quantitatively within this framework, and a molecular model for meiotic double-strand break repair based on the concept of sliding D-loops is offered as an efficient scheme for visualizing the salient results from studies of crossing over and gene conversion, the molecular structures of recombination intermediates, and the biochemical competencies of the proteins involved.
E
UKARYOTES transit from the diplophase to thehaplophase via meiosis, which is associated with a number of interrelated processes, including crossing over and gene conversion. These processes involve meiosis-specific, programmed DNA double-strand breaks (DSBs) and their repair (DSBr). DSBr, in turn, is associated with mismatched base pairs and their rectification, re-ferred to as ‘‘mismatch repair’’ or MMR (Bishop et al. 1987). Current efforts to accommodate both the gene-tic and molecular phenomena associated with meio-tic DSBr in yeast (Saccharomyces cerevisiae) have been thoroughly reviewed (e.g., Hollingsworthand Brill 2004; Hoffmannand Borts2004; Surteeset al. 2004; Hunter 2007; Berchowitz and Copenhaver 2010), but none of the reviews commits to an overall picture with quantitative predictions. This work aims to remedy that lack. Specifically, we have made use of salient published studies to develop, step-by-step, a compre-hensive model of meiotic DSBr and MMR. The main features of this model are summarized in Table 1.
RESULTS
For readers who are unfamiliar with yeast genetics and/or the known details of MMR, we begin by review-ing (1) the basic principles and vocabulary of tetrad analysis in yeast, which expose the products of
individ-ual acts of meiosis, (2) the DSBr model of Szostaket al. (1983) as modified by Sun et al. (1991), which has provided a basic molecular interpretation of meiotic recombination, and (3) the known roles of mismatch-repair proteins such asMsh2andMlh1.
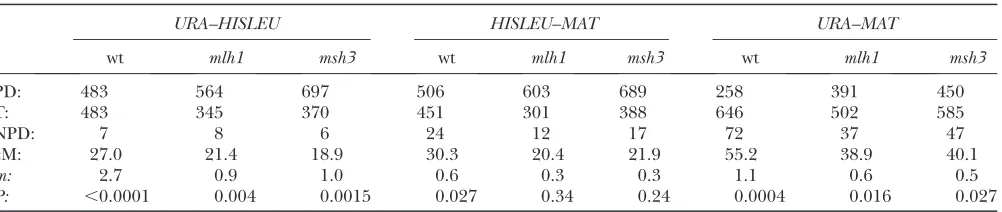
Relative frequencies of tetrad types provide meas-ures of linkage distance and crossover interference: Consider a population of diploid yeast cells heterozy-gous for two linked sites, A/a and D/d. When meiosis proceeds without a hitch, the resulting tetrads each contain four viable haploid spores. Because the geno-types of the spores are identifiable by the phenogeno-types of the colonies they give rise to, each spore in the tetrad can be characterized as a crossover or a noncrossover with respect to sites A/a and D/d. When the A/a and
D/d sites are closely linked, the most frequent tetrads contain only the two genotypes that characterized the parents;i.e., they have twoADand twoadspores and are therefore referred to as ‘‘parental ditype’’ tetrads (PD). Two other types of tetrads may be found in various fre-quencies: ‘‘tetratype’’ tetrads (T),i.e., those in which the spores are all different (AD, Ad, aD, ad), and ‘‘non-parental ditype’’ tetrads (NPD:Ad,Ad,aD,aD) represent-ing the two recombinant genotypes. This type of tetrad analysis—assessing the relative frequencies of PDs, T’s, and NPDs—allows a quantitative measure of crossing over (i.e.,linkage distances and crossover interference).
Gene conversion as evidence of DSBr: The same
diploid may be marked at one or two other sites, B/b
(and/orC/c), closely bracketing a ‘‘DSB hotspot,’’i.e., a site that receives a high frequency (e.g., 20%) of pro-grammed, meiosis-induced double-strand breaks. The
B/b (and C/c) sites serve to identify tetrads that have This article is dedicated to the Institute of Molecular Biology,
established January 1, 1959.
1Corresponding author:Institute of Molecular Biology, Onyx Room 287, 1229 University of Oregon, Eugene, Oregon 97403.
E-mail: [email protected]
undergone a DSBr event at the hotspot (Gilbertson and Stahl1996). Such tetrads are recognized by their failure to exhibit Mendelian (i.e., 2:2) segregation of the markers among the spores. Instead, a ‘‘conversion tetrad’’ may contain three spores with a marker derived from one parent, and one spore with the allele from the other parent (e.g., 3B:1b, or 3b:1B). This non-2:2 distribution of genotypes is also termed ‘‘aberrant’’ or ‘‘non-Mendelian.’’ Furthermore, because the chromo-some, a double-stranded DNA molecule, carries genetic information on each of its complementary strands, a haploid spore may give rise to a ‘‘mixed colony,’’ consis-ting of bothBandb(orCandc) cells. Tetrads contain-ing such a spore will be referred to as ‘‘half-conversions’’ (HCs) or 5:3. They can be diagrammed asBB,BB,Bb,
bb when B is in excess over b, or as bb, bb, bB, BB
whenbis in excess. In this article, we deal with the two most common types of conversions: the HCs or 5:3’s as described above, and the ‘‘full conversions’’ (FCs) or 3:1’s (aka6:2’s), which may be diagrammed asBB,BB,
BB,bborbb,bb,bb,BB). In general, tetrads with an excess ofBor ofbare found at similar frequencies. A signif-icant deviation from this expectation is referred to as ‘‘disparity.’’
Successful application of tetrad analysis requires careful placement of the markersABCD/abcd. Sites B/b and C/c must be located within a few hundred base pairs from a hotspot to efficiently register DSBr events as gene conversions. In contrast, markers A/a and D/d, designed to monitor crossing over and crossover inter-ference, must be far enough from the hotspot to rarely suffer gene conversion, but close enough to ensure that most crossovers betweenA/a and D/dare products of the DSBr events monitored byB/band/orC/c.
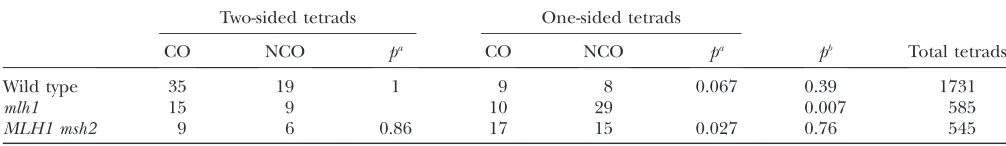
Because of limitations on the availability of markers, only one of the two sites closely bracketing a hotspot has typically been in a gene that determines a conveniently scored phenotype. Screening for conversions at that site has been used to select for analysis those tetrads that have undergone a DSBr event, involving nonsister chromatids, at the marked hotspot. Within that pop-ulation, the markers at the second, less conveniently scored site, have then been determined by DNA analysis. The use of conversion at one marked site, guaranteeing that homologous DSBr has occurred at the hotspot, allows more meaningful scoring of the other site for 2:2 segregation (as well as for FC or HC) of the marker. By this procedure, tetrads are identified as ‘‘two-sided’’ (conversions for both B/b and C/c) and ‘‘one-sided’’ (conversions for only the more conveniently scored site).
The Szostak/Sun model for meiotic DSBr: We can
use the Szostak/Sun DSBr model (Figure 1) to illustrate how DSBr could generate a variety of the observed tetrads. For example, Figure 1A illustrates how mismatched base pairs created by the loss of genetic information from the chromatid undergoing DSBr, and its replacement with information from the unbroken homolog, can create a
HC. Should a 39-end be degraded past the site of a marker near the DSB, the repair process is called ‘‘gap repair’’ and results, invariably, in FC for the marker, as shown in Figure 1B. Such gap repair presumably occurs independently of MMR proteins. MMR, too, can gener-ate FCs, but it opergener-ates only in the presence of known MMR proteins. MMR occurring at invasion or annealing will be directed by the invading or annealing termini and results only in FCs (Figure 1C). Should a mismatch escape MMR at invasion or annealing, it becomes sub-ject to MMR directed by the termini created by Holliday junction resolution. Because, in this model, both the Watson and the Crick chains are cut to effect resolution, MMR can, with presumed equal probability, result in an FC or in restoration to 2:2 segregation (Figure 1C and 1D). Insofar as 2:2 segregation for a marker close to a DSB hotspot (in a tetrad with conversion for a close marker on the other side of the DSB site) can be at-tributed to restoration, MMR directed by junction re-solution can be inferred. Note that Figure 1 does not show all four chromatids, but illustrates the fate of only the two chromatids directly involved in the DSBr process. Many basic features of the Szostak/Sun model have survived more than two decades’ worth of tests. These features include steps leading up to the formation of the joint-molecule, ligated DSBr intermediate (Figure 1A, step 4), and the existence of the intermediate is not in doubt (Schwachaand Kleckner 1995). The ligated intermediate, however, is now understood to give rise (by an, as-yet, unknown mechanism) to crossovers only (Allers and Lichten2001a; reviewed in Bishop and Zickler2004). In this work we present further depar-tures from the Szostak/Sun model inspired (1) by the demonstration (Getzet al.2008) that, in yeast, meiotic DSBr occurs via either a ‘‘pairing pathway,’’ first pro-posed by Zalevskyet al.(1999), or a ‘‘disjunction path-way’’ (see Stahlet al. 2004), and (2) by the extensive data of Hoffmannet al.(2005), which allow us to propose and quantify a set of DSBr-pathway-specific properties.
Since this analysis rests, to a large extent, on the known functions of MMR proteins in meiosis, and since much of our understanding of these proteins is based on studies of the bacterial anti-mutation proteins MutS and MutL, we summarize here the current understanding of MutS, MutL and their eukaryotic homologs.
Conventional skinny on MMR in meiosis: The
those of their bacterial cousins. For example, MutS homologs recognize mismatches and attract MutL ho-mologs to the region, and MutL hoho-mologs attract the remaining components of the mismatch-repair machin-ery. The expected manifestations of such MMR in meio-sis are dependent on the DSBr structures in which it is presumed to operate as well as on the intrinsic repair-ability of the particular mismatch.
Among MutS homologs, the Msh4–Msh5 hetero-dimer (Burnset al. 1994; Hollingsworthet al. 1995; Pochartet al. 1997) stands out in several respects. First, the Msh4–Msh5 heterodimer alone is meiosis specific (Ross-MacDonaldand Roeder1994) and required for wild-type levels of crossing over and interference (Novak et al. 2001) as well as for the formation of an intermedi-ate (Figure 1A, step 1) leading to (detectable) ligintermedi-ated joint-molecule DSBr intermediates (Figure 1A, step 4) (Bo¨rneret al. 2004). The meiosis specificity is consistent with the failure of Msh4–Msh5 to affect the mitotic mutation rate. However, the absence of Msh4–Msh5 also fails to manifest an increase in the frequency of HCs, the
expected consequence of defective meiotic MMR (Ross -MacDonaldand Roeder1994; Hollingsworthet al. 1995; Wanget al. 1999). This, plus the observation that Msh4–Msh5 protein has no identified mismatch recog-nition sequence like that of MutS and its yeast homolog Msh2(Culliganet al. 2000), has been taken as evidence that Msh4–Msh5 lacks the ability to effect MMR. (We revisit this view below.) Themsh4–msh5-induced loss of crossing over, without a detected loss of either DSBr or MMR, suggests that the DSBs that were fated to have been interhomolog crossovers have, instead, been repaired using the sister chromatid as jig and template. Armed with this background, we first review the observations that force revision of the Szostak/Sun model of DSBr and, second, develop the alternative model (Table 1) for DSBr and MMR, along with arguments demonstrating the adequacy and economy of the model.
Two pathways for meiotic DSBr:In an effort to reveal
the relationship between DSBr and MMR, Getz et al. (2008) used tetrad analysis to monitor, at each of two hotspots, conversion of a palindrome marker (Naget al.
Figure1.—Classic DSBr. The model of Szostaket al. (1983) as modified by Sunet al. (1991) related double-strand break repair,
1989). DSBr-induced mismatches involving such palin-drome markers often fail to undergo MMR in the pre-sence of normal MMR proteins (Figure 1A). We refer to such mismatches as ‘‘poorly repairable mismatches’’ or PRMs. In addition, Getzet al. (2008) monitored cross-ing over between markers bracketcross-ing the palindrome and the DSB site. They conducted these crosses in the presence or absence of Msh4, a protein required for wild-type levels of crossing over and interference as described above. Their data show that the frequencies of only those crossover tetrads with FC or with nonconver-sion for the palindrome marker (in many of which a PRM had presumably undergone restorational repair) were dependent on the presence ofMsh4 protein. In contrast, the frequencies of both crossover and non-crossover tetrads with HC for the marker (i.e., those in which MMR of the PRM had failed) were, to a good ap-proximation,Msh4independent. Similarly, in wild-type yeast, the crossover tetrads with FC or nonconversion for the palindrome marker showed positive interference, while the crossover tetrads with HC for the palindrome lacked such interference.
Pathway-specific rules for MMR:The observations by
Getzet al. (2008) provide good evidence that wild-type yeast, unlikeS. pombe(Cromieand Smith2008), has two DSBr pathways with different MMR properties, both of which yield crossovers. Specifically, when DSBr creates a PRM via theMsh4-dependent, interference-generating disjunction pathway, the PRM undergoes efficient re-pair, yielding a crossover with either FC or nonconver-sion for the palindrome marker. In contrast, when the PRM is created by DSBr in theMsh4-independent, non-interference, ‘‘pairing pathway,’’ the PRM is refractory to repair, yielding either a HC crossover or a HC non-crossover. (Note the implication that if any FCs for the palindrome marker arise in the absence ofMsh4, they must have resulted from gap repair.)
Msh2 deletion and PRMs are equivalent ways of
eliminating Mmr from the pairing pathway:The results
of Getzet al. (2008) provide an incentive to revisit, in the framework of two pathways for DSBr, the powerful study
by Hoffmannet al. (2005), which was also designed to explore relationships between DSBr and MMR. The methods used by Hoffmann et al. (2005) differ from those used by Getzet al. (2008). Specifically, Hoffmann et al. (2005) monitored DSBr as conversion for markers that make rather ‘‘well-repairable mismatches’’ (WRMs) in a wild-type background, and they created MMR deficiencies by deletion of the MutS homologMsh2or the MutL homolog Mlh1. Getz et al. (2008), instead, monitored DSBr events as conversions for a marker that makes PRMs near a DSB site. They noted that failedvs. successful MMR for the marker was associated with Msh4independence or dependence, respectively, which enabled them to assign DSBr events to one or the other DSBr pathway. In particular, they noted that MMR was effective only in the disjunction pathway (Getz et al. 2008). Lest the reader protest that comparing Hoffmannet al. (2005) with Getzet al. (2008) is com-paring apples and oranges, we argue, on the basis of the following observations, that failure to undergo repair of
PRMs on the one hand and deletion of Msh2 on the
other are equivalent manifestations of failed MMR in the pairing pathway, and that pathway only.
Stone and Petes (2006) demonstrated that, for
a WRM near a DSB site, deletion of MSH2 causes a
shift from FCs to HCs independently of Msh4. This implies thatmsh2-deletion mutants lack MMR in the Msh4-independent, pairing pathway, but have no
con-spicuous MMR in the Msh4-dependent, disjunction
pathway. Getz et al. (2008) demonstrated that, in a
MSH2background, failure of the PRM to be repaired is also independent of Msh4. This implies that the observed HCs for the PRM, too, represent failure of MMR in the pairing pathway. Together, these obser-vations suggest that a PRM involving a palindrome in
a MSH2 background may be defined as a mismatch
that is refractory toMsh2-dependent MMR. The work of Wanget al. (2003), showing that Msh2interacts ab-errantly with a palindrome mismatch, further supports our thesis that the methods of Hoffmannet al. (2005) and those of Getzet al. (2008) support each other. TABLE 1
Proposed properties of two DSBr pathways
Features Pairing pathway Disjunction pathway
Products Crossovers and noncrossovers Crossovers only Crossover Interference No positive interference Positive interference
Msh4–Msh5 dependence None Total
Bimolecular intermediate Long with junctions not fully ligated Short with fully ligated Holliday junctions
Invasion heteroduplex Partly ephemeral Ephemeral MMR at invasion and annealing Dependent on Msh2 and Mlh1 None MMR near the DSB site Directed by 39invading and annealing
ends
Mlh1 dependent; directed by junction resolution
Role of Msh2 in MMR Recognizes mismatches and attracts Mlh1 None
Deletion of Mlh1 causes greater loss of MMR than
does deletion of Msh2:In their exploration of DSBr and
MMR, Hoffmannet al. (2005) had analyzed tetrads of the ABCD/abcd-type illustrated above, with markers his
(which makes rather well repairable mismatches) and
BIK(which makes WRMs) representing the genericB/b
and C/c sites bracketing the hotspot. Accordingly, the authors registered DSBr events as conversions forhisand/ orBIK. Scoring, initially, conversion forhis, they showed that the elimination of eitherMlh1 orMsh2resulted in a shift from full- to half-conversion, as predicted by the
Szostak/Sun model. However, the absence of Mlh1
caused a greater MMR deficiency,i.e.,a greater increase in HC/(FC1HC), than did the absence ofMsh2 (Appen-dix A). In a one-pathway model for DSBr, this might imply thatMlh1simply affects MMR more efficiently than does Msh2. In a two-pathway context, on the other hand, the quantitative difference in phenotypes suggests a require-ment forMlh1 to affect MMR not only in the pairing pathway in conjunction with Msh2, but also, without Msh2, in the disjunction pathway. Further analysis of their data supports the hypothesis of Mlh1-dependent MMR in the disjunction pathway, as described below.
Evidence for disjunction-pathway-specific,
Mlh1-dependent MMR:Likemsh4–msh5mutants, but unlike
msh2 mutants, mlh1 mutants show reduced meiotic
crossing over (Hunterand Borts1997) and reduced interference (Abdullah et al. 2004; Appendix B). Moreover, these mlh1 phenotypes are observed in a
MSH4–MSH5 background only (Wang et al. 1999;
Arguesoet al. 2004). Thus, at least with respect to cross-ing over and interference,MLH1functions in the dis-junction pathway. The data of Hoffmannet al.(2005) indicate that the observed mlh1-induced gain in HCs mentioned above representstwicethemlh1-induced loss in FCs (Appendix A). Data in the same article suggest (1) that the disjunction-pathway-specific,Mlh1-dependent FC tetrads were crossovers, while themlh1-induced HCs were noncrossovers and (2) that these noncrossovers represented twice the number ofMlh1-dependent cross-overs (Appendix C). These observations imply thatMlh1 does, indeed, play a role in MMR in the disjunction path-way and that50% of the time,Mlh1-dependent MMR in the disjunction pathway restores Mendelian (2:2) se-gregation of the marker.
In the disjunction pathway, Mlh1-dependent MMR
occurs only in response to junction resolution: The
evidence that Mlh1-dependent, disjunction-pathway-specific MMR yields restoration and FC tetrads at equal frequencies implies that such MMR was directed by resolution of the Holliday junctions of the ligated DSBr intermediate (which is the molecular hallmark of the disjunction pathway) (Figure 1D). A corollary of this view is that mismatches created at the invasion and/or annealing phases of disjunction-pathway DSBr fail to undergo MMR prior to being incorporated in the ligated intermediate. Work by Allers and Lichten
(2001b) supports this interpretation. These authors
used gel electrophoresis of DNA from aMSH2 MLH1
strain to characterize DSBr intermediates with respect to a palindrome marker that makes PRMs near a DSB site. As expected, they found that the intermediate frequently contained the marker in mismatched, het-eroduplex DNA, indicating a paucity of MMR prior to ligation of the intermediate. Moreover, in none of the intermediates were all four of the marked DNA strands derived from only one parent or the other, which would have been an indication of MMR.
The results of Allersand Lichten(2001b) indicate that PRMs in the disjunction pathway generally escape MMR at the invasion and annealing phases of DSBr. As discussed above, the data of Hoffmann et al. (2005), who used markers that make rather well-repairable mismatches, show that disjunction-pathway-specific MMR of WRMs, too, is directed by junction resolution. Thus, in wild-type yeast, the lack of MMR at the invasion and annealing phases appears to be a regular feature of disjunction-pathway DSBr. Yet, as demonstrated by Getzet al. (2008), mismatches induced by DSBr in the disjunction pathway are invariably repaired. Thus, MMR in this pathway occurs always, and only, in response to Holliday junction resolution.
msh2-induced lack of Mmr reduces two sidedness:As
shown above, lack of MMR in the disjunction pathway revealed DSBr events that would not have been detected in the presence of MMR. The data of Hoffmann et al. (2005) allow us to ask if the absence ofMsh2-dependent (i.e.,pairing-pathway specific) MMR would produce the same result. These authors screened for tetrads with conversion athis. Within thishis-conversion population, they compared conversion frequencies for their second marker, BIK,in msh2vs. MSH2strains. Their results showed that deletion ofMsh2caused an increase in HCs among conversion tetrads, as expected. At the same time, however, two sidedness (the conversion frequency ofBIKamonghisconversions) wasdecreased, rather than increased in response to loss ofMsh2. Getzet al. (2008) reported equivalent results from crosses in a MMR-proficient background. To select for DSBr events at the hotspot, they used a ‘‘B/b’’ site (close to a DSB hotspot) that made WRMs. The crosses contained a ‘‘C/c’’site (on the other side of the DSB hotspot) to assess two sidedness. When theC/csite made WRMs, most of theC/c conver-sions tetrads were FCs, as expected. When they replaced the WRMC/csite with aC/csite that made PRMs, most of the conversions at the C/csite were now HC, also as ex-pected, but two sidedness was significantly reduced.
Migrating D-loops and transient heteroduplex: How
en-larges the D-loop, in Figure 2 extension of the invading end causes the lagging as well as the leading end of the D-loop to move toward the DSB site (Ferguson and Holloman 1996; Hoffmann and Borts 2005). As a result, a mismatch formed at invasion is undone as the invading strand is extruded to reunite with its original partner. If the mismatch undergoes MMR before it is ‘‘undone’’ by the migrating D-loop, full conversion for the marker will indicate that a mismatch had been created. If the mismatch fails to be repaired promptly, evidence that a mismatch had been created at invasion may be erased by migration of the D-loop.
A role for Msh4–Msh5 in disjunction-pathway MMR: The concept of transient heteroduplex at invasion, together with ‘‘use-it-or-lose-it’’ conversion opportunities, satisfactorily accounts for MMR-dependent two sided-ness in the pairing pathway. [Unwinding of the in-vasion heteroduplex (SDSA, Paquesand Haber1999) could provide a second ‘‘use-it-or-lose-it’’ route to MMR-dependent two sidedness. As such, one sidedness due to lack of MMR would be enriched among noncrossovers relative to crossovers (Merkeret al. 2003; Getz et al. 2008)]. A migrating D-loop, causing transient hetero-duplex, may well characterize disjunction pathway DSBr also. Indeed, data from Allersand Lichten(2001b) and Schwacha and Kleckner (1995) demonstrated that a conspicuous fraction of ligated (i.e., disjunction-pathway) DSBr intermediates had both Holliday junctions on the same side of the DSB site. If the disjunction pathway does, in fact, have transient invasion heteroduplex, mismatches created at invasion will be lost without a trace, because the disjunction pathway appears to routinely forego MMR at invasion and annealing, even in wild-type crosses.
What could prevent MMR in wild-type strains from acting at invasion and annealing in the disjunction
pathway? By way of answer, we suggest that the lack of MMR prior to completion of the ligated intermediate in this Msh4–Msh5-dependent pathway is due to the ab-sence of MutS function required for recognizing
mis-matches in duplex DNA. Msh2 and Msh4–Msh5, the
only known candidates for this role, are both disquali-fied, although for different reasons—Msh2, which does recognize mismatches in duplex DNA, does not operate in the disjunction pathway (see Appendix A), while Msh4–Msh5, which, by definition, does operate in the disjunction pathway, fails to recognize mismatches in duplex DNA. It remains to be considered how, in the absence of mismatch recognition, PRMs, and WRMs too, in the disjunction pathway are nevertheless invari-ably repaired to yield either FC or nonconversion tet-rads (Getzet al. 2008). Work by Snowdenet al. (2004) suggests how the unique properties of Msh4–Msh5 might allow the dimer to promote MMR in a joint-molecule double-Holliday-junction DSBr intermediate. Snowdenet al. (2004), working with human MutS and MutL homologs, concluded that the behavior of Msh4– Msh5 protein at a Holliday junction is like that of a MutS protein at a mismatch in duplex DNA—Msh4–Msh5 binds to a Holliday junction and then slides away (Acharyaet al. 2003). The high concentration of Msh4–Msh5 could then attract the MutL homolog. In the case of the double-Holliday-junction intermediate, reiteration of such behav-ior, with sliding in either direction, could lead to a traffic jam of Msh4–Msh5 in the region between the junctions, attractingMlh1to the entire region. Now, when nicks are introduced to resolve a junction, every mismatch between the junctions is rectified. Whether the DNA removal and replacement required for rectification is an inevitable consequence of junction cutting or is dependent on the mismatch is not answerable at this time.
The dog that didn’t bark: If, as we propose, there is
such a thing as Msh4–Msh5/Mlh1-dependent MMR in the disjunction pathway, why then would msh4–msh5
deletion mutants not have a MMR-deficiency pheno-type? This lack ofmsh4–msh5-induced increase in HCs is economically explained by the requirement of Msh4– Msh5 for the establishment of the disjunction pathway (Bo¨ rneret al. 2004). Without the disjunction pathway and its products to register the presence or absence of MMR, there can be no msh4-induced increase in HCs to signal that MMR had failed. Hence, the apparent paradox—any MMR-deficiency phenotype ofmsh4–msh5
mutants should be detectable only in the presence of Msh4–Msh5 protein.Msh4–Msh5’s vital contribution to the establishment of the disjunction pathway makes it impossible to challenge directly the proposal that, within the disjunction pathway, MMR is Msh4–Msh5 dependent. A critical test of Msh4–Msh5’s involvement in MMR may require an (as-yet hypothetical)msh4–msh5mutant that has retained the ability to form/stabilize the double-Holliday-junction DSBr intermediate, but has lost the ability to recruit Mlh1. Perhaps such a
separation-of-Figure2.—The traveling D-loop. (Step 1) A D-loop is
function mutant will be found and will exhibit the typical meiotic MMR-deficiency phenotype, viz., an increase in HCs.
Where the rubber meets the road: The DSBr and
MMR data from the extensive study by Hoffmannet al. (2005) were reported in terms of HCs and FCs forhis
and BIK in wild-type,msh2 and mlh1 crosses. In addi-tion, as noted above, these authors screened tetrads for two sidedness. These data can be used to test whether the model is capable of generating those observed values. Such a test requires that we first identify and evaluate (see Appendix A) the probabilities for each of the steps that can lead to a specified outcome. They are of two kinds—those whose values are preset by the model (nonadjustable parameters) and those that are specific to the HIS4 DSB hotspot and the markers in Hoffmann’s strains (adjustable parameters).
Our model implies the following values for the four nonadjustable parameters:
d The probability that hisis on the annealing side of
any DSBr intermediate¼½ (Whenhisis not so sit-uated,BIKis)
d The probability that a mismatch on the annealing side
of the DSBr intermediate in the disjunction path-way is repaired¼1
d The probability that repair in the disjunction pathway
leads to FC¼½
d The probability that a mismatch on the invading side
of DSBr in the disjunction pathway remains within the migratory D-loop¼0.
The adjustable parameters are:
d g, the probability, specific for each marker, that a
mis-match in the pairing pathway becomes FC by gap TABLE 2
Summary of Sudoku
Parameter Valuea
A. Values for adjustable parameters athis
D, disjunction pathway events per 1000 tetrads 154
P, pairing pathway events per 1000 tetrads 171
E, fraction ofPretaining heteroduplex on invasion side 0.3
R, probability that heteroduplex rejection does not occur 0.643
ghis, probability of gap repair athis 0.127
gBIK,probability of gap repair atBIK 0.04
mhis, probability of mismatch rectification to FC athisin pairing pathway 0.726
mBIK, probability of mismatch rectification to FC atBIKin pairing pathway 0.96
Class Obs.b Calc.c
B. Conversion athis
mlh1HC 171 174
mlh1FC 21 21.8
msh2HC 97 97
msh2FC 61 60.3
WT HC 17 17.1
WT FC 120 122
Cross Obs.d Calc.e
C. Fraction of two-sidedhisconversions
mlh1 0.26–0.47 0.31
msh2 0.22–0.46 0.39
WT 0.58–0.77 0.71
Cross Obs.f Calc.e
D. FC/(FC1HC) forBIKamonghisconversions
mlh1 4/41 0.10
msh2 3/51 0.10
WT 89/92 0.97
a
From Tables A2, A3, and A4.
b
From Table A1.
c
From Tables A2 and A3.
d
95% confidence interval on data from Table 5 of Hoffmannet al. (2005, Table 5).
eFrom Table A4. fFrom Hoffmann
repair (Szostak et al. 1983) or FC-biased ‘‘short-patch repair’’ (Coı¨cet al. 2000)
d R, the probability that heteroduplex rejection in the
pairing pathway (Chamberset al. 1996; Goldfarb and Alani2005) does not occur (see Appendix A) d m, the probability, specific for each marker, that
mismatches in the pairing pathway are repaired (always to FC rather than 2:2)
d E, the probability that, on the invasion side of a
pair-ing pathway DSBr event, the mismatch remains covered by the traveling D-loop so that it appears as an HC in MMR-deficient crosses
d P, the probability (expressed as number of DSBs per
thousand tetrads) that a DSB is repaired by way of the pairing pathway
d D,the probability (expressed as number of DSBs per
thousand tetrads) that a DSB is repaired by way of the disjunction pathway.
This list of parameters intentionally excludes the possibility of restoration of 2:2 segregation by MMR acting on a mismatch that is close to an initiating DSB. In so doing we minimize the number of parameters.
Sudoku:To estimate values for the adjustable
param-eters, we adopted the conventional strategy of starting with the parts of the puzzle that look easiest. For example, to obtain a value for D, we made use of the model’s feature that DSBr products contributed by the pairing pathway should be strictly the same formlh1as formsh2crosses. Hence, any differences between those two crosses should lead directly to estimates ofD, the only adjustable parameter in the disjunction pathway. To determine the values ofghis, P, and E, we first esti-matedEon the basis of the frequency of tetrads, in the MMR mutants, that were simultaneously conversions for
BIKandhis(two-sided tetrads). We assume a single value forE, rather than assuminghis- andBIK-specific values, on the grounds tha, if we can fit the data with a single value, we could surely fit them with separate values. The remaining two parameters were then chosen to give satisfactory fits to the FC and HC data for the MMR mutants (Appendix A).
To obtain ahis-specific value formand a value ofR, we turned to the HC and FC frequencies in the wild-type cross. We heldD,ghis, E,andP at the values deduced from the MMR-mutant crosses, and, for simplicity, as-sumedmto apply equally to mismatches created at in-vasion or annealing (Appendix A). The strategy for obtaining values for the only remaining parameters,mbik
andgbik, is described in Appendix A.
With a value for each of the parameters, it was then possible to compare the expected values for FC, HC, and two sidedness with the observed values. The summary (Table 2) demonstrates that a single set of plausible parameter values satisfies both the HC and FC data, as well as the two sidedness, for each of the three genotypes,msh2,mlh1, and WT. It is gratifying that, with
eight adjustable parameters, the model can account for 12 observations.
DISCUSSION
The data of Hoffmannet al. (2005) and Getzet al. (2008) call for an updated view of DSBr and MMR. Our interpretation of these data, above, has allowed us to
Figure3.—Proposed features of the disjunction pathway of
assign specific attributes to each of two pathways for DSBr in wild-type yeast (Table 1), leading to a molecular model that illustrates how repair of the programmed, meiotic double-strand breaks might occur in each of the two pathways (Figures 3 and 4).
The model invites us to revisit several meiotic phe-nomena previously published and interpreted. These include the wide variation in the estimated lengths of regions of heteroduplex resulting from DSBr, MMR-dependent two sidedness, the apparent mutual
exclu-sivity of interference and gene conversion in Sordaria fimicola, and an unexpected DSBr intermediate. Below, we review these phenomena and offer interpretations within the framework of our two-pathway model with traveling D-loops.
Heteroduplex lengths: Several studies of DSBr in
yeast have yielded data that create impressions of the length of the regions of heteroduplex created at the invasion and/or the annealing stages of DSBr. Some estimates were based on physical analyses (e.g.,
micros-Figure4.—Proposed features of the pairing pathway of DSBr inmsh2andmlh1meioses. (A) Noncrossovers: (Step 1) A D-loop is
copy or gel electrophoresis), others on genetic analyses. The problem is that the impressions appear to contradict each other. For example, genetic analyses suggest that conversion tracts (which depend on regions of hetero-duplex) may often be ‘‘long’’ (Detloffet al. 1992; Foss et al. 1999). Microscopy by Belland Byers(1983), on the other hand, indicates that double-Holliday-junction in-termediates tend to be ‘‘short.’’ This estimate by micros-copy is consistent with gel electrophoresis data reported by Schwachaand Kleckner(1995), which imply that the region of DNA between two Holliday junctions in observed intermediates is usually short. Our model suggests that these apparent discrepancies reflect the differences between the pairing and disjunction path-ways. Both the gel electrophoresis and the microscopy focus on double-Holliday-junction intermediates, i.e. on disjunction-pathway intermediates, which, in our model, are short (Figure 3, and see below). In contrast, the genetic studies indicating long regions of hetero-duplex (Figure 4; Detloffet al. 1992; Fosset al. 1999; Hillers and Stahl1999) were based on HC tetrads, which are manifestly products of the pairing pathway (Getzet al. 2008; and see above). This interpretation is also consistent with themsh4-induced increase in the average length of conversion tracts, as deduced from genetic data of Mancera et al. (2008)—according to our model, the absence ofMsh4 would eliminate the disjunction pathway with its short conversion tracts, thereby increasing the average conversion tract length. It should be noted that electrophoresis studies have yielded little evidence on the structure of pairing-pathway intermediates. While this could reflect an ephemeral nature, we suggest that it reflects (instead or also) the variable and ‘‘unfinished’’ nature of these intermediates (Figure 4) and/or a length that frequently exceeds the distance between the restriction sites used by the investigator to liberate the intermediates from the chromosome. Furthermore, frequent erosion of the 39-single-strand ends at the DSB in the pairing pathway (g.0) could confound the detection of DSBs specific to that pathway.
MMR-dependent two sidedness: Hoffmann et al.
(2005) explained the phenomenon of MMR-dependent two sidedness with the suggestion ‘‘... that in wild-type cells the initial DSB repair event is two sided. The absence of MMR, by either mutation or use of poorly repaired palindromes, allows a second, unbiased mis-pair removal pathway to restore a proportion of hetero-duplexes, leading to apparent one-sided events.’’ This proposal suffers from several problems, one of which is that the short-patch system hypothesized by Hoffmann et al. (2005) to be responsible for this unbiased MMR is claimed by its discoverers in yeast (Coı¨cet al. 2000) to be biased against the marker on the invading strand, thus favoring FCs over restorations (and see Appendix A,
Disparity between the two classes of HCs). Our model suggests, instead, that a traveling D-loop in the pairing
pathway allows a mismatch created at invasion only a transient opportunity to enjoy MMR (Figure 4). Thus, the one sidedness (fraction of conversions for one of the bracketing markers that are 2:2 for the other) reflects the failure of MMR to turn such a mismatch into an FC before the heteroduplex containing the mismatch is undone.
Gene conversion in Sordaria: Our model for DSBr
obliges us to revisit the observation by Kitani(1978) that, unlike yeast crossovers, Sordaria crossovers that exhibit gene conversion have no crossover interference, and vice versa. Stahland Foss(2008) had suggested that, in both organisms, MMR in the disjunction pathway is directed by junction resolution. In yeast such MMR would result in interfering crossovers with FC or 2:2 segregation for the marker with equal probability while, in Sordaria, junction-directed MMR would yield interfering crossovers with only 2:2 segregation. This proposal rested on the assumption that the disjunction
Figure 5.—The JM2 DSBr intermediate of Allers and
Lichten(2001a,b). Steps 1 and 2 are as in Figure 3. (Steps
3 and 4) If the traveling D-loop overshoots the marker to the right of the DSB site before being halted by annealing, the marker is in heteroduplex composed of an old red strand and a new blue strand. (Steps 5 and 6) Prior to the comple-tion of the Holliday junccomple-tion on the right, the unligated junc-tion, posing as a nicked juncjunc-tion, directs Mlh1-dependent MMR of the mismatch, restoring the parental state (speckled) of this disjunction pathway intermediate. Schwacha and
Kleckner(1995) reported a high frequency of JM2s (their
pathways for the two organisms differ from each other in the same manner as do the pairing pathways;viz. yeast has predominantly asymmetric heteroduplexes, revealed by 5:3 segregation, while Sordaria has a high frequency of symmetric heteroduplex, revealed as aberrant 4:4 segregation (Kitaniand Olive1967; Stahland Hillers 2000; Stahland Foss2008). We now suggest a second possibility as to why interference and conversion appear mutually exclusive in Sordaria. This possibility is based on our understanding that Kitani’s monitored markers were not selected for proximity to a DSB hotspot. If, as for yeast, Sordaria’s regions of heteroduplex in disjunction-pathway intermediates are relatively short, Kitani’s markers might only rarely generate disjunction-pathway mismatches, in which case the interfering crossovers would usually lack conversion regardless of whether the intermediate had symmetric or asymmetric heterodu-plex. In contrast, crossovers in the pairing pathway, with its longer conversion tracts, would be relatively more likely to involve a marker far from the DSB site in heteroduplex.
This explanation (for the frequent lack of detectable DSBr participation in the disjunction pathway for marked sites relatively remote from the DSB site) may apply also to the apparent lack ofBIK’s participa-tion in disjuncparticipa-tion pathway DSBr (Appendix A,The BIK data).
An unexpected DSBr intermediate: Allers and
Lichten(2001b) reported a DSBr intermediate that they labeled JM2. JM2 had two Holliday junctions, identifying it as a disjunction-pathway intermediate. The junctions, however, were on the same side of the DSB site, beyond a palindrome marker located near the DSB site (on the right in Figure 5). Moreover, the palindrome marker was in parental configuration, which the authors had some difficulty explaining. Within the framework of our model featuring a traveling D-loop and junction-directed repair in the disjunction pathway, we offer the scheme illustrated in Figure 5. A most attractive feature of the scheme is that, in harmony with the observations of Allersand Lichten (2001a), it yields restoration intermediates but never FC ones. We note that if JM2s had been frequent in the data of Hoffmannet al. (2005), our model would demand that the excess of HCs seen in themlh1cross would be more than twice the excess of FCs seen in theMLH1 msh2cross. The data do not lean that way.
Predictions: Our view of the roles of the various
Mut homologs in meiotic MMR makes a number of predictions:
1. Crosses carried out with a poorly repairable mis-match close to a DSB site in a MSH2 background should have the same HC/(FC1 HC) ratios as do crosses in amsh2background with a marker, at the same site, that makes well-repairable mismatches. 2. The proposal that deletion ofMSH4in yeast
elimi-nates a crossover pathway that yields only
noncon-versions and one-sided FCs implies thatmsh4mutants should show a modest increase not only in the HC/ (FC1HC) ratio for a palindrome marker atHIS4(by decreasing FC crossovers as shown by Getz et al. 2008), but also in the fraction of DSBr events that are two sided (by decreasing the one-sided tetrads). 3. Getzet al. (2008) noted that, inmsh4mutants, the
loss of conversion crossovers appeared to result conspicuously in a gain in nonconversion noncross-overs (presumably by sister repair). The studies of Hoffmann et al. (2005) suggest that, in contrast, deletion of MLH1 appears to turn FC interfering crossovers into HC noncrossovers. The possibility that these HC noncrossovers would interfere with each other is a heady one, but difficult to test. 4. Another test of the model concerns the effect ofMsh4
on HC/(FC1HC) ratios inmsh2andmlh1mutants. Although these two single mutants differ appreciably with respect to HC/(FC1HC) ratios at his, when the disjunction pathway is removed by deletion ofMSH4
themsh2 msh4andmlh1 msh4double mutants should be seen to have identical values for that ratio. 5. According to our model, only the pairing pathway
produces two-sided tetrads, implying that two-sided crossovers should not manifest positive interference. Expected and observed phenotypes for a variety of relevant genotypes are summarized in Table 3.
A prudent investigator aiming to challenge these predictions would probably choose to work at HIS4 and with the strains of Hoffmannet al. (2005).
We thank Eva Hoffmann, John Fowler, Rhona Borts, Nancy Hollingsworth, Elizabeth Housworth, Michael Lichten and anony-mous reviewers for helpful comments. Jette Foss provided invaluable editorial assistance on Appendix A. Unless otherwise specified, statis-tical calculations were conducted with the aid of VassarStats (http:// faculty.vassar.edu/lowry/VassarStats.html).
LITERATURE CITED
Abdullah, M. F. F., and R.H. Borts, 2001 Meiotic recombination
frequencies are affected by nutritional states inSaccharomyces
cer-evisiae.Proc. Natl. Acad. Sci. USA98:14526–14529.
Abdullah, M. F. F., E.R. Hoffmann, V.E. Cottonand R. H. Borts,
2004 A role for the MutL homologueMLH2in controlling
het-eroduplex formation and in regulating between two different crossover pathways in budding yeast. Cytogenet. Genome Res.
107:180–190.
Acharya, S., P. L. Foster, P. Brooksand R. Fishel, 2003 The
co-ordinated functions of the MutS and MutL proteins in mismatch
repair. Mol. Cell12:233–246.
Allers, T., and M. Lichten, 2001a Differential timing and control
of noncrossover and crossover recombination during meiosis.
Cell106:47–57.
Allers, T., and M. Lichten, 2001b Intermediates of yeast meiotic
recombination contain heteroduplex DNA. Mol. Cell 8:225–
231.
Argueso, J. L., D. Smith, J. Yi, M. Waase, S. Sarin et al.,
2002 Analysis of conditional mutations in theSaccharomyces
cer-evisiae MLH1gene in mismatch repair and in meiotic crossing
over. Genetics160:909–921.
Argueso, J. L., J. Wanat, Z. Gemiciand E. Alani, 2004 Competing
crossover pathways act during meiosis inSaccharomyces cerevisiae.
Bell, L. R., and B. Byers, 1983 Homologous association of
chromo-somal DNA during yeast meiosis. Cold Spring Harbor Symp.
Quant. Biol.47:829–840.
Berchowitz, L. E., and G. P. Copenhaver, 2010 Genetic
interfer-ence: don’t stand so close to me. Curr. Genomics11:91–102.
Bishop, D. K., and D. Zickler, 2004 Early decision: meiotic
cross-over interference prior to stable strand exchange and synapsis.
Cell117:9–15.
Bishop, D. K., M. S. Williamson, S. Fogeland R. D. Kolodner,
1987 The role of heteroduplex correction in gene conversion
inSaccharomyces cerevisiae.Nature328:362–364.
Bo¨rner, G. V., N. Klecknerand N. Hunter, 2004
Crossover/non-crossover differentiation, synaptonemal complex formation, and regulatory surveillance at the leptotene/zygotene transition of
meiosis. Cell117:29–45.
Burns, N., B. Grimwade, P. B. Ross-MacDonald, E-Y. Choi, K. Finberg
et al., 1994 Large-scale analysis of gene expression, protein
local-ization, and gene disruption inSaccharomyces cerevisiae. Genes. Dev.
8:1087–1105.
Chambers, S. R., N. Hunter, E. J. Louisand R. H. Borts, 1996 The
mismatch repair system reduces meiotic homeologous recombi-nation and stimulates recombirecombi-nation-dependent chromosome
loss. Mol. Cell. Biol.16:6110–6120.
Coı¨c, E., L. Gluckand F. Fabre, 2000 Evidence for short-patch
mis-match repair inSaccharomyces cerevisiae.EMBO J.19:3408–3417.
Cotton, V. E., E. R. Hoffmann, A. F. F. Abdullahand R. H. Borts,
2009 Interaction of genetic and environmental factors in
Sac-charomyces cerevisiaemeiosis: the Devil is in the details. Methods
Mol. Biol.557:3–20.
Cromie, G. A., and G. R. Smith, 2008 Meiotic recombination in
Schizzosacharomyces pombe: a paradigm for genetic and molecular
analysis. Genome Dyn. Stab.3:195–230.
Culligan, K. M., G. Meyer-Gauen, J. Lyons-Weilerand J. B. Hays,
2000 Evolutionary origin, diversification and specialization of
eukaryotic MutS homolog mismatch repair proteins. Nucleic
Acids Res.28:463–471.
Detloff, P., M. A. Whiteand T. D. Petes, 1992 Analysis of a gene
conversion gradient at theHIS4locus inSaccharomyces cerevisiae.
Genetics132:113–123.
Ferguson, D. O., and W. K. Holloman, 1996 Recombinational
re-pair of gaps in DNA is asymmetric inUstilago maydisand can be
explained by a migrating D-loop model. Proc. Natl. Acad. Sci.
USA93:5419–5424.
Foss, E., R. Lande, F. W. Stahland C.M. Steinberg, 1993 Chiasma
in-terference as a function of genetic distance. Genetics133:681–691.
Foss, H. M., K. J. Hillersand F. W. Stahl, 1999 The conversion
gradient atHIS4inSaccharomyces cerevisiae. II. A role for mismatch
repair directed by biased resolution of the recombinational
inter-mediate. Genetics153:573–583.
Getz, T. J., S. A. Banse, L. S. Young, A. V. Banse, J. Swansonet al.,
2008 Reduced mismatch repair of heteroduplexes reveals
‘‘non’’-interfering crossing over in wild-typeSaccharomyces
cerevi-siae.Genetics178:1251–1269.
Gilbertson, L. A., and F. W. Stahl, 1996 A test of the double
strand break model for meiotic recombination inSaccharomyces
cerevisiae.Genetics144:27–41.
Goldfarb, T., and E. Alani, 2005 Distinct roles for the
Saccharomy-ces cerevisiaemismatch repair proteins in heteroduplex rejection,
mismatch repair and nonhomologous tail removal. Genetics169:
563–574.
Hillers, K. J., and F. W. Stahl, 1999 The conversion gradient at
HIS4ofSaccharomyces cerevisiae.I. Heteroduplex rejection and
res-toration of Mendelian segregation. Genetics153:555–572.
Hoffmann, E. R., and R. H. Borts, 2004 Meiotic recombination
in-termediates and mismatch repair proteins. Cytogenet. Genome
Res.107:232–248.
Hoffmann, E. R., and R. H. Borts, 2005 Transevents associated
with crossovers are revealed in the absence of mismatch repair
genes inSaccharomyces cerevisiae.Genetics169:1305–1310.
Hoffmann, E. R., E. Eriksson, B. J. Herbert and R. H. Borts,
2005 MLH1andMSH2promote the symmetry of double-strand
break repair events at theHIS4hotspot inSaccharomyces cerevisiae.
Genetics169:1291–1303.
Hollingsworth, N. M., and S. J. Brill, 2004 The Mus81 solution
to resolution: generating meiotic crossovers without Holliday
junctions. Genes Dev.18:117–125.
Hollingsworth, N. M., L. Ponteand C. Halsey, 1995 Msh5, a
novel MutS homolog, facilitates meiotic reciprocal
recombina-tion between homologs in Saccharomyces cerevisiaebut not
mis-match repair. Genes Dev.9:1728–1739.
Hotchkiss, R. D., 1971 Toward a general theory of genetic
recom-bination in DNA. Adv. Genet.16:325–348.
Hunter, N., 2007 Meiotic recombination, pp. 381–442 inMolecular
Genetics of Recombination, edited by A. Aguileraand R. Rothstein.
Springer, New York.
Hunter, N., and R. H. Borts, 1997 Mlh1 is unique among mismatch
repair proteins in its ability to promote crossing-over during meiosis.
Genes Dev.11:1573–1582.
Hunter, N., and N. Kleckner, 2004 The single-end invasion: an
asym-metric intermediate at the double-strand break to double-Holliday
junction transition of meiotic tecombination. Cell106:59–70.
Jessop, L., and M. Lichten, 2008 Mus81/Mms4 endonuclease and
Sgs1 helicase collaborate to ensure proper recombination
inter-mediate metabolism during meiosis. Mol. Cell31:313–323.
Kitani, Y., 1978 Absence of interference in association with gene
conversion inSordaria fimicola, and presence of interference in
association with ordinary recombination. Genetics89:467–497.
Kitani, Y., and L. S. Olive, 1967 Genetics ofSordaria fimicola. VI.
Gene conversion at theglocus in mutant3wild type crosses.
Genetics57:767–782.
Lamb, B. C., and M. R. T. Wickramaratne, 1973
Corresponding-site interference, synaptinemal complex structure, and 81:0m
and 71:1m octads from wild-type3mutant crosses ofAscobolus
immersus.Genet. Res.22:113–124.
Maloisel, L., J. Bhargavaand G. S. Roeder, 2004 A role for DNA
polymerase in gene conversion and crossing over during meiosis inSaccharomyces cerevisiae.Genetics167:1133–1142.
Mancera, E, R. Bourgon, A. Brozzi, W. Huberand L. M. Steinmetz,
2008 High-resolution mapping of meiotic crossovers and
non-crossovers in yeast. Nature454:479–485.
Merker, J. D., M. Dominskaand T. D. Petes, 2003 Patterns of
hetero-duplex formation associated with the initiation of meiotic
recom-bination in the yeastSaccharomyces cerevisiae.Genetics165:47–63.
Modrich, P., 1991 Mechanisms and biological effects of mismatch
repair. Annu. Rev. Genet.25:229–253.
Nag, D. K., M. A. Whiteand T. D. Petes, 1989 Palindromic
sequen-ces in heteroduplex DNA inhibit mismatch repair in yeast. Nature
340:318–320.
Nakagawa, T., A. Dattaand R. D. Kolodner, 1999 Multiple
func-tions of MutS- and MutL-related heterocomplexes. Proc. Natl.
Acad. Sci. USA96:14186–14188.
Novak, J. E., P. B. Ross-MacDonaldand G. S. Roeder, 2001 The
bud-ding yeast Msh4 protein functions in chromosome synapsis and the
regulation of crossover distribution. Genetics158:1013–1025.
Oh, S. D., J. P. Lao, A. F. Taylor, G. R. Smith and N. Hunter,
2008 RecQ Helicase, Sgs1, and XPF family endonuclease,
Mus81-Mms4, resolve aberrant joint molecules during meiotic
recombination. Mol. Cell31:324–336.
Papazian, H. P., 1952 The analysis of tetrad data. Genetics37:175–188.
Paques, F., and J. E. Haber, 1999 Multiple pathways of
recombina-tion induced by double-strand breaks inSaccharomyces cerevisiae.
Microbiol. Mol. Biol. Rev.63:349–404.
Perkins, D., 1949 Biochemical mutants of the smut fungusUstilago
maydis. Genetics34:607–626.
Pochart, P., D. Wolteringand N. M. Hollingsworth, 1997
Con-served properties between functionally distinct MutS homologs in
yeast. J. Biol. Chem.272:30345–30349.
Radford, S. J., S. McMahan, H. L. Blanton and J. Sekelsky,
2007 Heteroduplex DNA in meiotic recombination in
Dro-sophilamei-9mutants. Genetics176:63–72.
Ray, A., N. Machin and F.W. Stahl, 1989 A DNA double-chain
break stimulates triparental recombination inSaccharomyces
cere-visiae.Proc. Natl. Acad. Sci. USA86:6225–6229.
Ross-MacDonald, P., and G. S. Roeder, 1994 Mutation of a
meiosis-specific MutS homolog decreases crossing over but not mismatch
Schwacha, A., and N. Kleckner, 1995 Identification of double
Holliday junctions as intermediates in meiotic recombination.
Cell83:783–791.
Shinohara, M., K. Sakai, A. Shinohara and D. K. Bishop,
2003 Crossover interference inSaccharomyces cerevisiaerequires
a TID1/RDH54- and DMC1-dependent pathway. Genetics163:
1273–1286.
Snowden, T., S. Acharya, C. Butz, M. Berardini and R. Fishel,
2004 hMSH4–hMSH5recognizes Holliday junctions and forms a meiosis-specific sliding clamp that links homologous
chromo-somes. Mol. Cell15:437–451.
Stahl, F. W., 2008 On the ‘‘NPD ratio’’ as a test for crossover
inter-ference. Genetics179:701–704.
Stahl, F. W., and H. M. Foss, 2008 But see Kitani(1978). Genetics
178:1141–1145.
Stahl, F. W., and K. J. Hillers, 2000 Heteroduplex rejection in
yeast? Genetics154:1913–1916.
Stahl, F. W., and E. A. Housworth, 2009 Methods for analysis of
cross-over interference inS. cerevisiae.Methods Mol. Biol.557:35–53.
Stahl, F. W., and R. Lande, 1995 Estimating interference and linkage
map distance from two-factor tetrad data. Genetics139:1449–1454.
Stahl, F. W., H. M. Foss, L. S. Young, R. H. Borts, M. F. F. Abdullah
et al., 2004 Does crossover interference count inSaccharomyces cerevisiae?Genetics168:35–48.
Stone, J. E., and T. D. Petes, 2006 Analysis of the proteins involved
in thein vivorepair of base–base mismatches and four-base loops
formed during meiotic recombination in the yeastSaccharomyces
cerevisiae.Genetics173:1223–1239.
Sun, H., D. Trecoand J. W. Szostak, 1991 Extensive 39-overhanging,
single-stranded DNA associated with the meiosis-specific
double-strand breaks at theARG4recombination initiation site. Cell64:
1155–1161.
Surtees, J. A., J-L Arguesoand E. Alani, 2004 Mismatch repair
proteins: Key regulators of genetic recombination. Cytogenet.
Genome Res.107:146–159.
Szostak, J., T. L. Orr-Weaver, R. J. Rothstein and F.W. Stahl,
1983 The double-strand-break repair model for recombination.
Cell33:25–35.
Terasawa, M., H. Ogawa, Y. Tsukamoto, M. Shinohara, K.
Shirahige et al., 2007 Meiotic recombination-related DNA
synthesis and its implications for cross-over and
non-cross-over recombinant formation. Proc. Natl. Acad. Sci. USA104:
5965–5970.
Wang, H., Y. Yang, M. J. Schofield, C. Du, Y. Fridman et al.,
2003 DNA bending and unbending by MutS govern mismatch
recognition and specificity. Proc. Natl. Acad. Sci. USA 100:
14822–14827.
Wang, T-F., N. Klecknerand N. Hunter, 1999 Functional
specific-ity of MutL homologs in yeast: Evidence for three Mlh1-based heterocomplexes with distinct roles during meiosis in recombi-nation and mismatch correction. Proc. Natl. Acad. Sci. USA
96:13914–13919.
Zalevsky, J., A. J. MacQueen, J. B. Duffy, K. J. Kemphues and
A. M. Villeneuve, 1999 Crossing over duringCaenorhabditis
elegans meiosis requires a conserved MutS-based pathway
that is partially dispensable in budding yeast. Genetics 153:
1271–1283.
Communicating editor: M. Johnston
APPENDIX A: ASSIGNING PARAMETER VALUES (F.W.S.)
Hoffmannet al. (2005) reported the values for the relative frequencies of two-vs.one-sidedness as well as those of HCs and FCs forhisandBIKin wild-type,msh2,
and mlh1 crosses. To test whether the DSBr model
proposed above (Table 1 and Figures 3 and 4) can accommodate these values it was necessary to evaluate the parameters that were identified as defining the model (Table 2). This appendix describes how such evaluation was achieved.
Parameter values were assigned on the basis of data in Hoffmannet al. (2005, Table 5). The key observation is that the fraction of FCs among conversions in themlh1
mutant strain (12/112) is significantly less than that in themsh2strain (33/86;P,0.0001). These data provide the statistical support for the view that, whereasMlh1is required for MMR in both the pairing and disjunction pathways,Msh2has no role in the disjunction pathway. To allow comparison between the observed and expectedhisconversion frequencies (Table 2) inmsh2,
mlh1,and wild-type strains, we expressed the entries in Hoffmann et al. (2005 Table 5) as events per 1000 tetrads, rounding to whole numbers (Table A1; a convenience that exaggerates the significance of the
msh2and mlh1 data, while reducing that of the wild-type data). In addition, we combined the four conver-sion classes (6:2, 2:6, 5:3, 3:5) of Hoffmann et al. (2005) into two classes, FC and HC, ignoring for now the implications of some conversion disparities (Ap-pendix D). Finally, we assumed that theBIKandhisends
of the broken chromosome are equally likely to initiate invasion.
We now have the tools to evaluate the adjustable parameterD. Following the model, which assumes that
msh2andmlh1have identical phenotypes in the pairing pathway, the excess of HCs in themlh1strain over HCs
in the MLH1 msh2 strain (171 97 ¼ 74: Table A1)
corresponds toD/2, the number of tetrads in whichhisis involved on the annealing side of a disjunction-pathway event. Also by the model, the excess of FCs inMLH1 msh2
over FCs inmlh1should beD/4. That excess is 6121¼
40 (Table A1). We arrive at a value forDby comparing the two mutants with respect to HCs (D/2¼74;D¼148) and, independently, by comparing them with respect to FCs (D/4¼40;D¼160). We average these values to get a working estimate ofD¼154 (Table A2).
To obtain values forPandghisfor each of the mutant strains, msh2 and mlh1, we write expressions for the number of tetrads demanded by the model to result in FC or HC forhis:
ForMLH 1 msh2: FC ¼ P ghis 1D=4;
HC ¼ ðP=2ÞðE11Þð1ghisÞ:
Formlh1: FC ¼P ghis;
HC¼ ðP=2ÞðE11Þð1 ghisÞ1D=2
are also conversions forBIK) and the rarity ofBIKFCs among these two-sided events (Hoffmann et al. 2005, Table 5; Table A4A), we setE, the fraction of pairing pathway events in which the sliding D-loop comes to rest over theBIKmarker on the invasion side of the DSB at
E¼0.3. Absent an estimate ofEforhis, we assume the same 0.3 value. With D¼154 andE ¼0.3, values for
Pcan be extracted from the equations. Solving the FC and HC equations formsh2, we getP¼172, while solving the equations formlh1givesP ¼166. We can then get values for ghisfrom both themsh2 and the mlh1 data. These are 0.131 and 0.127, respectively. ThePandghis
values for the two strains are, as expected, similar, and we settled on P ¼ 171 and ghis ¼ 0.127 for personal
reasons. (Recall that our modest goal is to determine whether there is a set of parameters that allows the model to fit the data, rather than to determine best estimates of those parameters.) Calculating the values for the FCs and HCs in the wild-type crosses (Table A3) required the evaluation of two additional parameters—one for the probability (mhis) that Msh2–Mlh1-dependent MMR leads to full conversion in the pairing pathway and one for the probability (R) that heteroduplex rejection does not occur. The need for R in the wild-type cross is signaled by the otherwise puzzling observation (Table A1) that total conversions at hisare lower in wild type than in the MMR mutants despite the greater two sidedness in wild type. In limiting the model to eight TABLE A1
Conversions athis
Conversion type per thousand tetradsa
6:2 2:6 FCb 5:3 3:5 HCc Tetrads (FC1HC)% HC/(FC1HC)%
Wild type 56 64 120 8 9 17 1731 13.7 12.4
msh2 28 33 61 31 66 97 545 15.8 61.4
mlh1 9 12 21 60 111 171 585 19.2 89.1
Conversions of the markerhis4–ATC(his), close to the DSB site. Adapted from Hoffmannet al. (2005, Table 5).
a
Ignores a few rare tetrad classes. Conversion types are rounded to the nearer whole number.
b
6:212:6.
c
5:313:5.
TABLE A2
Worksheet for confirming parameter values forhisin MMR mutants
Pairing pathway:P¼171a Disjunction pathway:D¼154b
Invasion: 1/2 Annealing: 1/2 Invasion: 1/2 Annealing:1/2
FC HC FC HC FC HC FC HC
msh2 g
0.127
E(1g) 0.262
g
0.127
1g
0.873
0 0 1/2 0
N 10.9 22.4 10.9 74.6 0 0 38.5 0
Summed conversions: Calculated: Observed:
FC 10.9110.9138.5¼60.3 61
HC 22.4174.6¼97 97
mlh1 g
0.127
E(1g) 0.262
g
0.127
1g
0.873
0 0 0 1
N 10.9 22.4 10.9 74.6 0 0 0 77
Summed conversions: Calculated: Observed:
FC 10.9110.9¼21.8 21
HC 22.4174.6177¼174 171
a
Estimated number of tetrads per thousand that enjoyed a DSB atHIS4and were repaired on a homologviathe pairing path-way.
b
adjustable parameters, we ignore a variety of possible additional factors with the hope that in so doing we are better exposing the skeletal features of the model. For instance, we are assuming that, for our markers, MMR in the pairing pathway is directed by the invading and annealing ends created by the DSB, with the result that all MMR in that pathway results in FC. Justification for this simplification is found in the adequacy of our simple model (Table 2).
We next address the features of the model that link two sidedness directly to MMR (Table A4). SinceBIK
and his are simultaneously present in all the crosses, their conversions must depend on common values ofP,
D,andR. Because we have assumed thatE, also, is the same forhisandBIK, we need pick onlymandgvalues forBIK. If there were a large body ofBIKdata analogous to thehisdata, we might have estimatedmBIKandgBIK
as we didmhisandghis. However, the best data come from theBIK conversions among tetrads selected for being
hisconversions, forcing a change in strategy. Thus, for both theMLH1 msh2andmlh1crosses, we sought and found agBIKvalue (0.04, by trial and error) that gave satisfactory fits to these two-sidedness data as well as to the FC/(FC1HC) value forBIKamonghisconversions in the two MMR-defective strains (Table A4A). For the wild-type cross, the only remaining parameter to be estimated ismBIK, which we chose to fit the two-sidedness data exactly (Table A4B). ThismBIKvalue proved to give a good fit to the wild-type HC/(FC1HC) ratio forBIK
amonghisconversions, supporting the view that theMlh1 -and Msh2-dependent two sidedness is a reflection of MMRper se.
Disparity between the two classes of HCs: Data of
Hoffmannet al. (2005) showed disparities in the rates of conversion tohisandHIS. In our Sudoku, we ignored the disparity, raising the possibility that in doing so we have concealed important information. The default hypothesis for disparity is differential rates of DSBs on the two homologs, and disparity so caused would be without consequence for our analysis. However,
Hoffmann et al. (2005), noting that the disparity was statistically significant only for the HCs, attributed it to different rates of restoration, by short-patch repair (Coı¨c et al. 2000), for the two different mismatches. This interpretation appeared to strengthen the authors’ proposal that short-patch repair, operating primarily in the absence ofMsh2andMlh1, was responsible for the
one sidedness seen in the msh2 and mlh1 crosses.
Shortcomings of this proposal, along with support for the differential DSB hypothesis, are detailed in Appen-dix D. The significance of a well-supported proposal for differential DSBs is that it undermines restoration by short-patch repair as an explanation for the one sidedness.
The BIK data: Hoffmann et al. (2005) noted that
the rate of conversion at BIK, especially in the MMR mutants, is less than that athis. Other aspects of theBIK
data combine with this observation to suggest that our Sudoku is not quite finished. Most of the BIKdata, in Hoffmann et al. (2005, Table 5), were collected from tetrads that were preselected ashisconversions. As such, they were two sided and, according to our model, must be from the pairing pathway. Consequently, thoseBIK
data were expected to be the same for themlh1and the
msh2 crosses, which they seem to be (mlh1—3 FC, 48 HC;msh2—4 FC, 37 HC;P¼0.7). That’s cool, butBIK, in unselected tetrads, fails to show the HC and FC differences that characterize thehisdata;i.e., for BIK, there is no evidence of mlh1-specific HCs or ofMLH1 msh2-specfic FCs. (mlh1—1 FC, 10 HC;msh2—0 FC, 9 HC). The numbers are small and could be ignored for that reason. However, the 0:9 ratio for BIK in themsh2
cross is significantly different (P ¼ 0.02) from the corresponding ratio, 33:53, for his. An economical interpretation for both the relatively low conversion rate of BIK (Hoffmann et al. 2005) and its failure to show properties characteristic ofhisin the disjunction pathway is that the disjunction pathway conversion tracts usually fail to include BIK. This may be simply because conversion tracts in the disjunction pathway are short andBIKis farther (maximum600 bp) from the TABLE A3
Worksheet for confirming parameter values forhisin wild type
Pairing Pathway:RP¼0.6433171 Disjunction Pathway:D¼154
Invasion: 1/2 Annealing: 1/2 Invasion: 1/2 Annealing: 1/2
FC HC FC HC FC HC FC HC
g1m(1g) 0.760
E(1g)(1m) 0.0718
g1m(1g) 0.760
(1g)(1m) 0.239
0 0 1/2 0
N 41.8 3.9 41.8 13.2 0 0 38.5 0
Summed conversions: Calculated: Observed:
FC 41.8141.8138.5¼122 120
HC 3.9 113.2¼17.1 17
HIS4 hotspot than is his (maximum 266 bp) (E. R. Hoffmann, personal communication). This possibility, which is of no consequence for our basic Sudoku, predicts that most or all BIK conversion crossovers come from the pairing pathway and, consequently, will have weaker interference than do his conversion crossovers.
Multiple events? In all studies of conversion at
re-combination hotspots in yeast there are tetrads that would be interpreted on the basis of any model as due to multiple DSBr events, and these tetrads are usually exempted from interpretation. However, as pointed out by Merker et al. (2003), the identification of the less obvious mul-tiple events is unavoidably model dependent. Further-more, the estimation of expected frequencies of multiple events is confounded by the possibility of negative in-terference between DSBs at the same level (Lamband Wickramaratne1973). Among other candidates for com-plex events we note two-sided HCs that are crossovers with heteroduplex on the same chromatid in thetrans
configuration (Hoffmannand Borts2005). Such tetrads
T ABLE A4 (Continued) Properties of BIK conversions Calculated Obser ved Freq. two-sided tetrads a mong his conversions (53.5 1 45.7)/ (45.7 1 55.0 1 38.5) ¼ 0.71 61/90 ¼ 0.57 0.77 a FC/ (FC 1 HC) BIK among his conversions 1 (0.6/53.5)(1/1.831) (2.1/55.0)(0.831/1.831) ¼ 0.97 89/92 ¼ 0.97 b HC/ (FC 1 HC} BIK in unselected tetrads (2.7)/(108.5 1 38.5) ¼ 0.02 0/9 b (FC 1 HC) BIK /(FC 1 HC) his (108.5 1 38.5)/ (100.7 1 38.5) ¼ 1.06 (10/107)/ (243/1731) ¼ 0.67 6 0.40 b V alues for P , D , R, and E, as well as for mhis and ghis , are as in T ables A2 and A3. E ¼ 0.3 and gBIK ¼ 0.04 were picked to fit the (FC 1 HC) and the FC/ (FC 1 HC) data for BIK among his conversions. mBIK ¼ 0.96 was then selected to fit the WT sidedness data. Expected numbers ( N ) are in parentheses after each expected conversion frequency . a H OFFMANN et al . (2005, T able 6). b H OFFMANN et al . (2005, T able 5).
Figure A1.—Crossovers that are trans-HC on the same