HIGHLIGHTED ARTICLE
| INVESTIGATION
Gap Junctions and NCA Cation Channels Are Critical
for Developmentally Timed Sleep and Arousal in
Caenorhabditis elegans
Huiyan Huang,*,1Dustin J. Hayden,*,2Chen-Tseh Zhu,†,3Heather L. Bennett,*,‡,4
Vivek Venkatachalam,§,**,5Lukas L. Skuja,* and Anne C. Hart*,6
*Department of Neuroscience, Robert J. and Nancy D. Carney Institute for Brain Science,†Department of Ecology and Evolution, and‡Department of Molecular Biology, Cell Biology, and Biochemistry, Brown University, Providence, Rhode Island 02912, §Department of Physics and **Center for Brain Science, Harvard University, Cambridge, Massachusetts 02138
ORCID IDs: 0000-0003-1239-7191 (H.H.); 0000-0001-7239-4350 (A.C.H.)
ABSTRACTAn essential characteristic of sleep is heightened arousal threshold, with decreased behavioral response to external stimuli. The molecular and cellular mechanisms underlying arousal threshold changes during sleep are not fully understood. We report that loss ofUNC-7orUNC-9innexin function dramatically reduced sleep and decreased arousal threshold during developmentally timed sleep inCaenorhabditis elegans.UNC-7function was required in premotor interneurons andUNC-9function was required in motor neurons in this paradigm. Simultaneous transient overexpression ofUNC-7andUNC-9was sufficient to induce anachronistic sleep in adult animals. Moreover, loss ofUNC-7orUNC-9suppressed the increased sleep ofEGL-4gain-of-function animals, which have increased cyclic-GMP–dependent protein kinase activity. These results suggest C. elegansgap junctions may act downstream of previously identified sleep regulators. In other paradigms, the NCA cation channels act upstream of gap junctions. Consistent with this, di-minished NCA channel activity inC. elegansrobustly increased arousal thresholds during sleep bouts in L4-to-adult developmentally timed sleep. Total time in sleep bouts was only modestly increased in animals lacking NCA channel auxiliary subunitUNC-79, whereas increased channel activity dramatically decreased sleep. Loss ofEGL-4or innexin proteins suppressedUNC-79loss-of-function sleep and arousal defects. InDrosophila, the ion channel narrow abdomen, an ortholog of theC. elegansNCA channels, drive the pigment dispersing factor (PDF) neuropeptide release, regulating circadian behavior. However, inC. elegans, we found that loss of the PDF receptorPDFR-1did not suppress gain-of-function sleep defects, suggesting an alternative downstream pathway. This study empha-sizes the conservation and importance of neuronal activity modulation during sleep, and unequivocally demonstrates that gap junction function is critical for normal sleep.
KEYWORDSCaenorhabditis eleganssleep; gap junction; cGMP-dependent kinase; NCA channel
D
URING sleep, animals have heightened arousal thresh-olds due to decreased response and desensitization of specific neurons (Campbell and Tobler 1984; Schwarzet al.2011; Nicholset al.2017). How does this transient and pro-found state change occur at a molecular level? Previous work has revealed multiple players, including neurotransmitters, neuropeptides, receptors, and kinases (Cirelli 2009; Allada
et al.2017). However, genes regulating sleep act in diverse pathways, suggesting that additional critical genes are not yet known and that understanding the relationship between these genes will be critical. Conserved genes regulating sleep can be identified in many model organisms, including
Caenorhabditis elegans(Trojanowski and Raizen 2016). Copyright © 2018 by the Genetics Society of America
doi:https://doi.org/10.1534/genetics.118.301551
Manuscript received August 28, 2018; accepted for publication October 10, 2018; published Early Online October 15, 2018.
Supplemental material available at Figshare: https://doi.org/10.25386/genetics. 7187531.
1Present address: Department of Pediatrics, Washington University School of Medicine,
St. Louis, MO 63110.
2Present address: Picower Institute for Learning and Memory, Massachusetts Institute
of Technology, Cambridge, MA 02139.
3Present address: Global Breeding, Bayer Crop Science, Chesterfield, MO 63017. 4Present address: Department of Biology, Reem-Kayden Center for Science and
Computation, Bard College, Annandale-on-Hudson, NY 12504.
5Present address: Department of Physics, Northeastern University, Boston, MA
02115.
6Corresponding author: Brown University, Mailbox GL-N, 185 Meeting St., Providence,
C. eleganssleeps spontaneously at specific times or under specific conditions (Van Buskirk and Sternberg 2007; Ghosh and Emmons 2008; Raizenet al.2008; Hillet al.2014). The best characterized sleep states are developmentally timed sleep and stress-induced sleep. Molecular and genetic path-ways required for sleep in these paradigms do not completely overlap, although commonalities have been identified (Trojanowskiet al.2015). Here, we focus almost exclusively on developmentally timed sleep, which occurs during a 2.5-hr period, called lethargus, coincident with cuticle molt-ing durmolt-ing diapause. Durmolt-ing lethargus, C. elegans exhibits interspersed bouts of sleep and motion that vary in duration, lasting from seconds to minutes. During sleep bouts, animals spontaneously cease locomotion, assume a specific posture, and show diminished response to external stimuli. During motion bouts, animals resume sinusoidal locomotion and re-spond robustly to external stimuli (Raizenet al.2008; Singh
et al.2011; Iwaniret al.2013). The mechanisms underlying this state change remain elusive.
Three major components of sleep behavior can be defined: circadian regulation, sleep homeostasis, and the state of sleep itself (Campbell and Tobler 1984). At this point, cir-cadian regulation is well described, but the other two are less understood. The molecular pathways that regulate these three components of sleep are likely interlinked and complex. Using an evolutionary and ethological strategy, it may be possible to disentangle these behaviors by studying species that have fewer circadian behaviors. While the state of sleep and sleep homeostasis are conserved inC. elegans, many aspects of circadian biology are not conserved. There is no evidence for an endogenous, genetically encoded 24-hr internal clock. C. elegans lacks classical proteins in-volved in light entrainment, like Cryptochrome. However, proteins intimately involved in circadian rhythms in other species, like Period and timeless, have C. elegansorthologs that play essential roles in the developmental timing of dia-pause and lethargus, but not circadian behavior (Banerjee
et al.2005; Monsalveet al.2011). Thus far, endogenousC. elegans circadian behaviors have not been described, al-though cyclic gene expression changes can be artificially entrained (van der Linden et al.2010). Other components of sleep are well conserved and a decade ofC. eleganssleep research has established the utility of this model organism in thefield. There is deep conservation of genes and path-ways regulating sleep and sleep homeostasis between C. elegans and other species (Singh et al. 2014; Trojanowski and Raizen 2016). Studies inC. eleganshave contributed to a growing list of genes affecting sleep, includingegl-4 /cyclic-GMP (cGMP)–dependent kinase, lin-12/glp-1/Notch receptors, aptf-1/AP2-family transcription factor, and gpb-2/Gbprotein (Raizenet al.2008; Singhet al.2011; Huang
et al.2017b).
Here, we synthesize previous work in the sleepfield with the results of an unbiased forward genetic screen for sleep genes inC. elegans, revealing a hierarchy of genes involved in sleep and arousal. In a classic genetic screen, we identify a
new role forUNC-7andUNC-9innexins in sleep andfind that these gap junction proteins are likely required downstream of the C. elegans cGMP-dependent protein kinase G (PKG), EGL-4. We alsofind that the NCA cation channels, identified in a mouse genetic screen for sleep genes (Funatoet al.2016), regulate arousal and sleep inC. elegans. Finally, we determine thatEGL-4PKG, but notC. eleganspigment dispersing factor (PDF) signaling, is required downstream of NCA channels to regulate sleep.
Materials and Methods
C. elegans culture and strain information
C. eleganswere cultured on standard nematode growth me-dia (NGM) seeded withOP50E. coli. Animals were grown at 25°and assayed at 22°unless noted otherwise. Strains used are listed in Supplemental Material, Table S1.
Microfluidic chamber-based assessment of L4/A sleep
The microfluidic, chamber-based sleep assay was adapted from previous studies (Singhet al.2011). Briefly, kanamycin-treatedOP50was resuspended as described in NGM (without agar) as a food source for the animals in the chambers. Mid-L4 stage animals, L4 substages L4.3–L4.4 (Moket al.
2015), were loaded into each chamber and covered with a glass coverslip, which was sealed to the chip-containing chambers using molten 2% agar. Images were recorded every 10 sec for 12 hr and analyzed using a MATLAB (MathWorks, Natick, MA) script for image subtraction (Singhet al.2011) and a custom Python script for calculating total time in sleep bouts (Huanget al.2017a).
Arousal threshold assessment (touch and light)
Arousal threshold assays with blue light stimulation were conducted as previously described (Huang et al. 2017b). Briefly, L4/A lethargus animals were picked to freshly made assay plates with food and allowed to recover for 15 min. During the assay, the plates remained unperturbed with the lids off. A blue light laser pointer was used to stimulate the animal over the head region. Response latency for animals during both sleep bouts and motion bouts was recorded. Ex-amination ofunc-7(e5)animals suggested possible defects in their response to blue light stimulation during motion bouts. Therefore, arousal thresholds determinations for unc-7and unc-9 studies tested latency to respond to posterior body gentle touch (Singh et al. 2011) in both sleep bouts and motion bouts. For all arousal determinations, at least eight animals per genotype and per condition (asleep or in motion) were tested in each trial. At least three independent trials were conducted. The experimenter was blinded to genotype.
PLM imaging
PLM GCaMP6 imaging in roaming animals was done with a custom-made system published earlier (Venkatachalamet al.
lethargus, was picked onto each assay plate (NGM in 100 mm plate seeded with 10mlOP50). Animals were covered with a 48365 mm #1 coverslip and left unperturbed for between 10 and 20 min before imaging. When imaging, the agar plate wasfit into a groove and secured with a custom-made metal plate screwed down on the plate. An automated tapping de-vice will exert controlled tapping when triggered. To avoid desensitization, the interval between tapping for an individ-ual animal was at least 10 min.
Plasmids and transgenic strains
Transgenic strains were generated by microinjecting plasmids into nematodes using standard methods (Evans 2006). Plas-mids used and transgene concentrations are listed in Table S2. Phenotypic rescue studies showed dose dependence. For unc-7 transgenes, high concentrations caused paralysis. For results herein, only transgene concentrations that did not cause paralysis were used.
Transient overexpression using heat shock
Transgenic animals with transgenes under the control of heat shock promoter were picked onto fresh plates at the young adult stage to induce transgene expression. Plates were sealed with Parafilm (Bemis Company) and heat shocked for 75 min in a 33.5°water bath, agar side down. Parafilm was removed and animals allowed to recover in a 20° incubator for 60 min. Then, animals were scored in the next 20-min in-terval. To avoid disturbing animals, plate lids were kept closed and plates were placed on the dissection scope agar side up (to avoid condensation on the inner surface of the plate lid when scoring). Animals were scored as Ans (in anachronistic sleep) if they did not move spontaneously or pump for 5 sec; either pharyngeal pumping or spontaneous locomotion was sufficient to score an animal as nonAns (not in anachronistic sleep).
To score locomotion and pharyngeal pumping separately, animals were treated as described above. Animals were scored as not moving if they did not move spontaneously for 5 sec, independent of the pumping status. Animals were scored as not pumping if they did not pump for 5 sec, regardless of locomotion. For all heat shock studies, at least 10 animals per genotype were tested for each trial and at least three in-dependent trials were conducted. The experimenter was blinded to genotype.
Statistics
Statistical analyses used Student’st-test for most of the studies. Mann–Whitney–Wilcoxon tests were performed for analyses of arousal thresholds with blue light because of the non-normally distributed data.
Data and reagent availability
Strains and plasmids are available upon request, unless other-wise stated. Full data tables are available upon request. Supplemental material available at Figshare:https://doi.org/ 10.25386/genetics.7187531.
Results
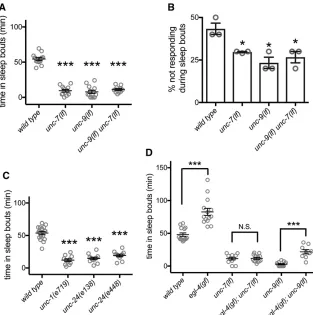
Loss of UNC-7 or UNC-9 gap junction proteins dramatically decreases sleep bouts during lethargus
To identify conserved genes required forC. elegans develop-mentally timed sleep during the transition to adulthood (L4/ A lethargus), we previously undertook a forward genetic screen (Huanget al.2017b). Subsequently, we determined that theunc-7(rt212)mutant allele, a premature stop muta-tion (W365Opal, isoform a), caused decreased total time in sleep bouts during L4/A lethargus (total sleep time, Figure S1A). The unc-7 gene encodes a C. elegans gap junction/ innexin protein with at least 10 alternative splice forms, dif-fering at the N terminus. Theunc-7(rt212)missense allele is predicted to prematurely truncate allunc-7splice forms. unc-7(rt212)decreased endogenous L4/A lethargus sleep defects (Figure S1A) and suppressed adult anachronistic sleep in-duced by Notch coligand OSM-11 overexpression (Figure S1B).
UNC-7normally functions withUNC-9to form gap junc-tions between cells. Loss of eitherUNC-7orUNC-9causes the animals to have uncoordinated locomotion with inappropri-ate posture predominantly during forward locomotion, but activity levels of adult animals are not decreased (Starich
et al. 1993, 1996, 2009). We found thatunc-7(e5) loss-of-function orunc-9(e101)loss-of-function animals had dramat-ically reduced L4/A lethargus total sleep (Figure 1A) and decreased lethargus sleep bout duration (Figure S1C). Intro-duction of a transgene containing theunc-9promoter driving unc-9 complementary DNA (cDNA) almost completely re-stored total sleep quantity in unc-9(e101) animals (Figure S1D). To corroborate the sleep defects observed based on motion detection, we also examined another important as-pect of sleep: altered arousal thresholds. During C. elegans
L4/A lethargus sleep bouts, response to a variety of external stimuli is diminished (Raizenet al.2008; Singhet al.2011; Huanget al.2017b); animals carrying mutations that perturb sleep quantity usually also have perturbed arousal thresholds during sleep bouts (Singhet al.2014). Consistent with this general rubric, we found arousal defects inunc-7(lf)and unc-9(lf)animals. While animals lacking unc-7orunc-9always responded to gentle touch outside of lethargus or during lethargus motion bouts, loss of either unc-7 or unc-9 decreased arousal thresholds specifically during sleep bouts (Figure 1B, response to gentle touch). Additionally, we undertook a small pilot study and found that changes in stimulus-evoked, intracellular calcium signals in mechano-sensory neurons that are characteristic of lethargus (Schwarz and Bringmann 2013; Cho and Sternberg 2014; Nicholset al.
andUNC-9function is important for modulating sensory neu-ron response, setting arousal thresholds during sleep bouts, and normal sleep quantity during lethargus.
Gap junction accessory proteins are also required for normal lethargus sleep bouts
The stomatin-like proteinsUNC-1andUNC-24are thought to regulate UNC-7/UNC-9 gap junction function via an un-known mechanism (Chen et al.2007);unc-24(lf)and unc-1(lf)animals have severe locomotion defects that are almost identical tounc-7(lf)andunc-9(lf)animals. Both of the unc-24(lf)alleles examined resulted in decreased total sleep time during L4/A lethargus (Figure 1C). Additionally, we tested the loss-of-function allele unc-1(e719) and the dominant gain-of-function allele unc-1(e1598) (Park and Horvitz 1986). unc-1 loss of function reduced total sleep time and unc-1gain of function increased total sleep time (Figure 1C and Figure S1F). Combined, these results confirm that re-duced gap junction accessory protein function results in de-creased developmentally timed sleep.
Gap junction proteins likely act downstream of EGL-4 PKG
Orthologs of theC. elegansEGL-4PKG, regulate sleep in ver-tebrates and inverver-tebrates (Raizen et al.2008; Langmesser
et al. 2009). InC. elegans, loss ofEGL-4function decreases sleep quantity and lowers arousal thresholds during devel-opmentally timed sleep. Further, increased EGL-4 activity
increases both sleep quantity and arousal thresholds (Raizen
et al. 2008). We confirmed these changes in sleep quantity and arousal thresholds for egl-4(n479) and egl-4(ad450), which are loss- and gain-of-function alleles, respectively. To our knowledge, no downstream targets ofEGL-4 PKG per-tinent to sleep have been identified in any species. To determine ifC. elegansgap junction proteins might act stream, we examined double mutant animals, as the down-stream gene usually suppresses in this context (Huang and Sternberg 1995). We found that the increased sleep of egl-4(ad450) gain-of-function animals was completely sup-pressed by loss ofunc-7and thategl-4(ad450)increased sleep was almost completely suppressed by loss of unc-9(Figure 1D). This suggests thatEGL-4PKG acts upstream in a genetic pathway and that EGL-4 PKG requiresUNC-7gap junction function, and is heavily dependent on UNC-9 function, to increase sleep.
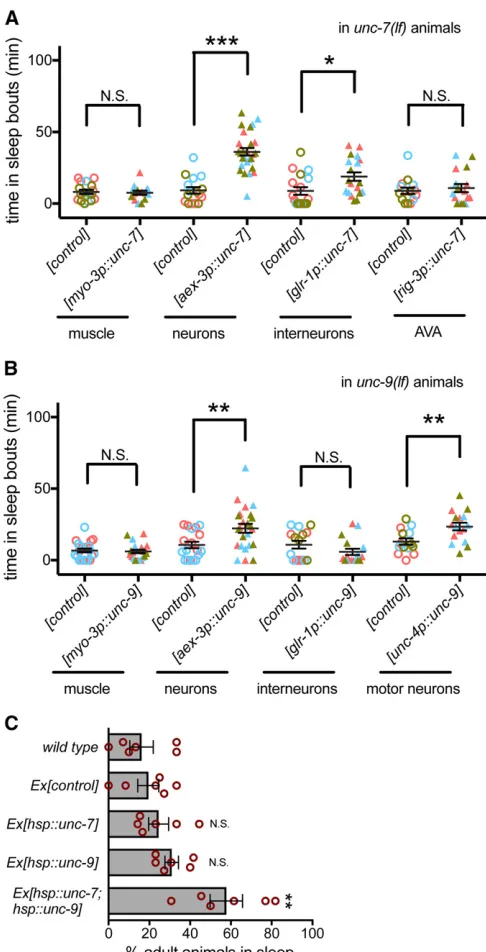
UNC-7 and UNC-9 act in interneurons and motor neurons, respectively
not restore wild-type level sleep; this may be a consequence of chimeric extrachromosomal array expression.
UNC-7function is required in premotor AVA interneurons for coordinated locomotion (Kawanoet al.2011). To deter-mine if UNC-7function is also required in AVA for normal sleep, we expressedUNC-7using theglr-1promoter, which drives expression in 17 classes of interneurons, including AVA interneurons (Hartet al.1995). This partially rescued sleep defects (Figure 2A). Expression of UNC-7 in AVA neurons alone using the rig-3promoter did not rescue sleep. Com-bined, these results suggest thatUNC-7function is required in neurons, and that UNC-7 function in a subset of glr-1 -expressing neurons plays a role in developmentally timed sleep.
UNC-9function is required in A-class motor neurons for coordinated locomotion (Kawano et al. 2011). Consistent with this, expression ofUNC-9cDNA using theglr-1promoter did not restore sleep (Figure 2B). However, expression of UNC-9 in cholinergic neurons, which includes the A-class motor neurons, partially restored sleep. These results suggest thatUNC-7/UNC-9gap junctions may function between pre-motor interneurons and pre-motor neurons to promote develop-mentally timed sleep, but their function in other neurons is likely also important.UNC-7andUNC-9act, at least in part, in interneurons and motor neurons to promote lethargus sleep bouts. The partial rescue of lethargus sleep bouts ob-served here may suggest that these gap junction proteins act at other sites to facilitate activity changes associated with sleep.
Simultaneous expression of both UNC-7 and UNC-9 induces anachronistic sleep in adult animals
UNC-7 andUNC-9are required for developmentally timed sleep, but is their activation sufficient to induce sleep? We tested this by inducing expression ofunc-7,unc-9, or both in adult animals using the heat-induciblehsp-16.2promoter to drive cDNA expression. After heat shock, C. elegans enter stress-induced quiescence, but in our hands, most wild-type animals resume locomotion and pharyngeal pumping 1 hr after heat shock ends. However, when UNC-7 and UNC-9 were simultaneously expressed, anachronistic sleep bouts were observed in adult animals 1 hr after heat shock (Figure 2C). Expression of eitherUNC-7orUNC-9was not sufficient, demonstrating that increasingUNC-7andUNC-9gap junc-tions is sufficient to induce anachronistic sleep in adult C. elegans and that both are required.UNC-7andUNC-9 can function as heterotypic gap junction channels or as hemi-channels; homomeric channels have been observed in ectopic expression systems (Starichet al.2009; Bouhourset al.2011; Menget al.2016). Heterotypic channels may be critical for the anachronistic sleep observed here.
The relationship between lethargus sleep and anachronis-tic sleep observed in adult animals after heat shock induced expression ofunc-7andunc-9may be complicated by contri-bution of stress-induced sleep, ectopic overexpression, and uncoupling of this behavior from the developmental context Figure 2 UNC-7 and UNC-9 is partially required in premotor
of diapause. Nevertheless, we were able to examine the consequences ofegl-4PKG loss of function in anachronistic sleep caused byUNC-7andUNC-9ectopic overexpression. Loss ofegl-4 did not significantly alter the fraction of ani-mals not pumping afterUNC-7/UNC-9ectopic overexpres-sion (Figure S2A), suggesting thatEGL-4is irrelevant or acts upstream in this paradigm. However,egl-4loss did suppress the locomotion changes caused by UNC-7/UNC-9 ectopic overexpression (Figure S2B), suggesting that EGL-4 may act downstream in this anachronistic paradigm. We are hes-itant to draw conclusions about developmentally timed sleep from anachronistic sleep, but unc-7 and unc-9 may act downstream ofegl-4for cessation of pumping in anach-ronistic sleep.
Normal NCA channel activity is required during C. elegans sleep bouts
In C. elegans, UNC-7 and UNC-9 act downstream of NCA cation channels to modulate response to anesthetics (Sedensky and Meneely 1987). The NCA ion channels (NALCN channels in mammals) are conserved cation chan-nels that are a member of the voltage-gated sodium and cal-cium channel family (Leeet al.1999; Ren 2011; Liebeskind
et al.2012). The NALCN/NCA channels used to be consid-ered sodium leak channels, although more functions have been proposed for these channels. (Senatore et al. 2013; Senatore and Spafford 2013; Boone et al. 2014) The NALCN/NCA channels have been implicated in various as-pects of sleep in flies and mice (Lear et al. 2005; Funato
et al. 2016). In flies, antiphase cycles in resting potassium and sodium conductance in clock neurons drive daily rhythms of activity, arousal, and sleep. This rhythmic sodium conductance is driven by sodium leak channel activity, which regulates circadian pacemaker neuron activity (Flourakis
et al. 2015). We hypothesized that, inC. elegans, the NCA cation channels might also regulate arousal and sleep. TheC. elegans NCA channel comprises the functionally redundant
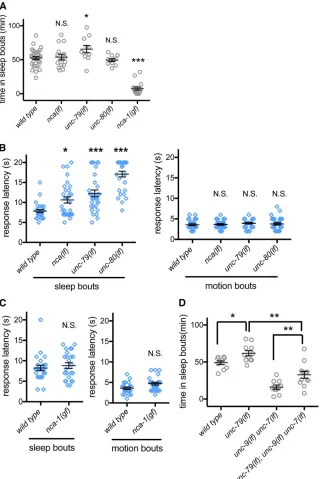
a1 subunitsNCA-1andNCA-2, which are encoded bynca-1 andnca-2genes, hence the name NCA channels.NCA-1and NCA-2act redundantly to regulate resting membrane poten-tial (Gaoet al.2015). Increased NCA channel function causes exaggerated body bends during locomotion, while NCA chan-nel loss causes short transient pauses inC. eleganslocomotion (Yehet al.2008). To assess the effect of NCA channels onC. eleganssleep during L4/A lethargus, we first examined the gain-of-function allele ofnca-1(e625): sleep bouts during L4/ A lethargus were almost completely eliminated in these an-imals (Figure 3A), suggesting that heightened NCA channel activity disturbs sleep.
Animals lacking both nca-1andnca-2are referred to as
nca(lf) animals in the literature (Yeh et al. 2008) and we adopt that convention here. We found thatnca(lf) animals had normal total time in sleep bouts during L4/A lethargus (Figure 3A). Other aspects of sleep/motion bout timing and duration were also normal when compared to wild-type an-imals (Figure S3, A and B). This discordance led us to more
closely examine the role of NCA channels and associated proteins inC. eleganssleep.
Functional NCA channels require two conserved auxiliary subunits, called UNC-79and UNC-80in C. elegans. These two genes do not function redundantly, as loss of either unc-79 or unc-80 perturbs NCA channel function with defects equal to nca(lf) animals (Yeh et al.2008). When we examined L4/A lethargus sleep in unc-79(lf)animals, we observed a small increase in total sleep time for two different mutant allele strains (Figure 3A and Figure S3C). However, total sleep time and other measures of L4/A sleep were normal inunc-80(lf)animals (Figure 3A and Figure S3, A–C). Given the dramatic effect ofnca(gf)on sleep, we considered the possibility that nca(lf) altered other aspects of L4/A lethargus sleep.
NCA loss-of-function animals had normal total sleep time, allowing us to assess arousal thresholds during sleep bouts. We previously established that blue light can be used to wake
C. elegans from sleep bouts and that time to respond (re-sponse latency) is an effective measure of arousal threshold (Huanget al.2017b). Loss-of-function alleles of NCA channel genes, including nca(lf),unc-79(lf), and unc-80(lf), caused increased latency to respond during sleep bouts (Figure 3B, left panel). To rule out the possibility that NCA channel func-tion is required for response to blue light, we also examined latency to respond during L4/A motion bouts. Time to re-spond to blue light was normal in nca(lf),unc-79(lf), and unc-80(lf)animals (Figure 3B, right panel). Assessing arousal thresholds innca-1(gf)animals was more challenging. Sleep bouts are rare and the average sleep bout duration in these animals was roughly 50% shorter (Figure S3B). With these caveats, we found that arousal thresholds in nca-1(gf) ani-mals were not different from wild-type aniani-mals (Figure 3C). Combined, these results suggest that appropriateC. elegans
NCA activity is critical for normal sleep.
We also examined sleep and arousal in animals defective in nlf-1function;NLF-1is an endoplasmic reticulum-associated protein that promotes, but is not absolutely required for, ap-propriate C. elegansNCA channel localization and function (Xie et al. 2013). The locomotion defects in adultnlf-1(lf)
animals were not as severe as those of unc-79(lf), unc-80(lf), ornca(lf)animals (Xieet al.2013). We did not observe changes in L4/A lethargus sleep quantity or in arousal thresh-olds for either sleep or motion bouts (Figure S3, D and E). Combined, all of these results confirm the conserved role of NCA channels in sleep-associated behaviors extends to C. elegans. Loss of NCA function leads to aberrantly increased arousal thresholds only during sleep bouts and increased NCA function prevents sleep bouts. Regulation of NCA chan-nel activity might be critical for entry into or maintenance of the sleep state with appropriately increased arousal thresholds.
NCA in any of these neuron populations rescued the arousal defects during lethargus (Figure S4C). These results are rem-iniscent of the distributed sites of action forC. elegansegl-4in sleep bout arousal (Raizenet al.2008) and consistent with the hypothesis that arousing factors can act at various loca-tions to awaken animals.
Gap junction protein function contributes to NCA channel modulation of sleep
Either NCA channel gain of function or gap junction loss of function results in decreased sleep. We examined the possi-bility that these genes act in the same pathway for sleep, as observed in other contexts (Sedensky and Meneely 1987; Bouhourset al.2011). For this analysis, we usedunc-79(lf)
animals, which have increased sleep, and introduced loss-of-function alleles forunc-7andunc-9. Loss ofUNC-7,UNC-9, or simultaneous loss of both gap junction proteins suppressed unc-79(lf)defects; total sleep time in triple mutant animals was not as low as sleep time in unc-9(lf)unc-7(lf)animals (Figure 3D), but was dramatically lower than the quantity of sleep observed in wild-type animals. We found that loss of onlyunc-7orunc-9also decreased sleep inunc-79(lf)animals (Figure S3F), to levels comparable to the triple mutant in Figure 3D. The incomplete suppression means either that these genes function in independent pathways or that gap junctions act genetically downstream of NCA channels in a pathway critical for lethargus sleep, along with other effectors.
C. elegansNCA channel loss of function leads to“fainting bouts,” which cause transient cessation of locomotion in adult animals (Yeh et al. 2008). We examined the conse-quences ofnca-1(gf)in anachronistic sleep caused by UNC-7andUNC-9ectopic overexpression in adult animals.nca-1(gf)
did not suppress pumping or locomotion changes observed in animals ectopically overexpressing UNC-7 and UNC-9 (Figure S2, A and B), suggesting that NCA channels either act upstream of gap junctions or are irrelevant in this paradigm.
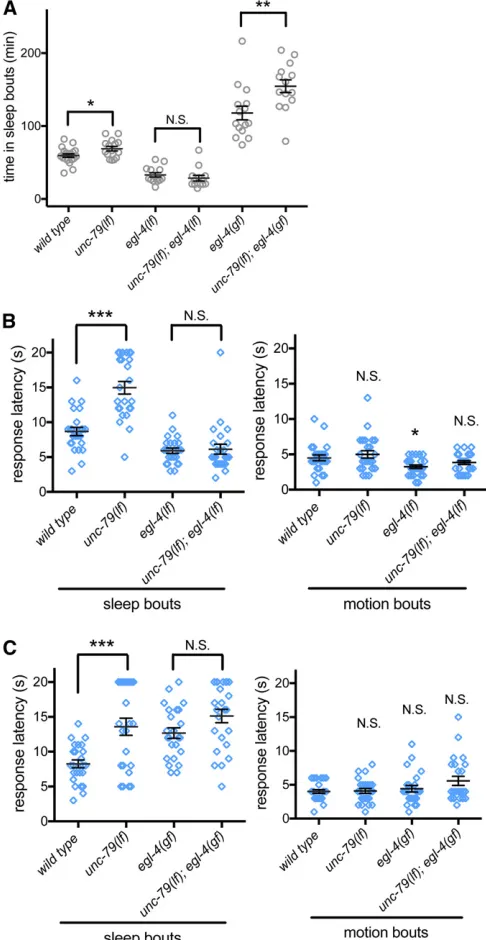
EGL-4 PKG may act downstream of NCA channels in C. elegans sleep
We also examined the relationship betweenEGL-4and NCA channels duringC. elegansL4/A lethargus, focusing initially on sleep quantity. We found that the increased sleep of unc-79(ec1)loss-of-function animals was completely eliminated by loss ofegl-4; total time in sleep bouts of animals lacking both unc-79 and egl-4was indistinguishable from animals with loss ofegl-4function alone (Figure 4A). Therefore, egl-4likely acts downstream ofunc-79in a genetic pathway that regulates sleep quantity. Next, we examined the arousal thresholds in the double mutant animals. Again,unc-79(ec1)
arousal changes during sleep bouts were completely sup-pressed by loss ofegl-4(Figure 4B). Combined, these results suggest thatEGL-4likely acts downstream of NCA channels for both sleep bouts and arousal threshold changes in a single pathway.
Sometimes, additional information can be gained by ex-amining gain-of-function alleles in epistasis studies. There-fore, we constructed double mutant unc-79(lf); egl-4(gf)
animals. There was no additivity in arousal changes between unc-79(lf)andegl-4(gf)animals (Figure 4C), consistent with EGL-4 and NCA acting in the same pathway for arousal threshold changes during L4/A sleep. However, we noted thatunc-79(lf);egl-4(gf)double animals had increased sleep compared with unc-79(lf) and egl-4(gf) single mutant ani-mals (Figure 4A). Because the increased sleep in double mu-tant animals is more than additive, interpretation is difficult, but not inconsistent with these two genes acting in the same pathway. Loss of NCA channel function may increase down-stream EGL-4 kinase activity. Alternatively, NCA channels may act throughEGL-4, as well as other mechanisms. Com-bined, these genetic epistasis studies suggest that NCA chan-nels likely act through theEGL-4PKG to regulate sleep and arousal.
NCA channels act downstream or in parallel to PDF signaling
Previous studies inDrosophilahave suggested that NCA chan-nels and PDF receptors act coordinately in DN1pcircadian neurons to regulate circadian behavior (Zhanget al.2010). In C. elegans, there are two PDF neuropeptide-encoding genes (pdf-1 andpdf-2) and one gene encoding a receptor (pdfr-1). Previous work inC. eleganshas shown in animals lacking neuropeptide Y signaling, loss ofPDF-1or thePDFR-1
receptor restores L4/A lethargus sleep quantity (Choi et al.
2013).PDF-1orPDFR-1loss also decreases motility in adult
C. elegans(Meelkopet al.2012; Choiet al.2013). However, a requirement for PDF signaling in L4/A lethargus sleep and arousal has not been reported previously.
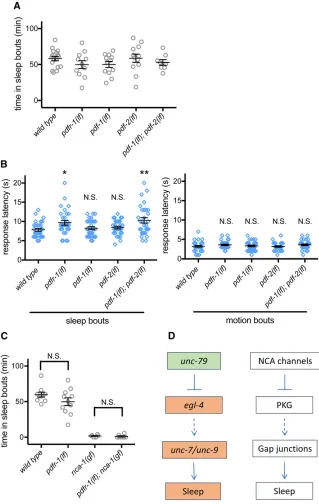
To directly determine the role of PDF signaling in lethargus sleep, we measured the total time in sleep bouts in pdfr-1(ok3425),pdfr-1(lst34),pdf-1(tm1996), orpdf-2(tm4393)
loss-of-function animals and inpdf-1(lf);pdf-2(lf)double mu-tant animals. All of these mumu-tant strains had normal total sleep during L4/A lethargus (Figure 5A and Figure S4A). We also examined arousal thresholds during L4/A lethargus sleep and motion bouts. Bothpdfr-1(lf)animals andpdf-1(lf); pdf-2(lf) double mutant animals had slightly increased arousal thresholds only during sleep bouts (Figure 5B and Figure S4B). Response latency was normal during motion bouts for all genotypes. pdf-1(lf) and pdf-2(lf)may act re-dundantly in this scenario, through thepdfr-1receptor. While the effect of altered PDF signaling was less dramatic than that of NCA channel perturbations, these results suggest that PDF neuropeptide signaling contributes to arousal during lethar-gus sleep bouts. Next, we examined the relationship between PDF signaling and NCA channels based on genetic epistasis. We generatedpdfr-1(lf); nca-1(gf)double mutant animals. PDFR-1loss did not ameliorate the sleep defects ofnca-1(gf)
animals (Figure 5C). Combined, these results suggest that PDF signaling is not required for NCA channel modulation ofC. eleganslethargus sleep and that loss of PDF signaling has a modest effect on arousal threshold during lethargus sleep bouts.
In summary, we have unequivocally established a require-ment forUNC-7andUNC-9gap junctions in L4/A lethargus sleep and arousal thresholds. We alsofind that NCA cation channels andEGL-4PKG likely act upstream ofUNC-7and UNC-9gap junction proteins in this behavior. NCA and PKG play critical roles in sleep and arousal across species, includ-ingC. elegans,Drosophila, and mice. We suggest that a path-way including these proteins and gap junctions may play a conserved role regulating sleep across all animal species (Fig-ure 5D).
Discussion
Here, we report that loss of C. elegans UNC-7 or UNC-9 innexin gap junction proteins resulted in dramatically re-duced time in sleep bouts during L4/A lethargus, de-creased bout duration, and dede-creased arousal thresholds during these sleep bouts.UNC-7function was partially re-quired in premotor interneurons andUNC-9function was partially required in motor neurons during sleep. More-over, transient, simultaneous ectopic overexpression of UNC-7 and UNC-9 induced anachronistic sleep in adult animals. Based on genetic epistasis studies in lethargus sleep,unc-7andunc-9acted downstream of egl-4, which encodes the C. elegans PKG. Additionally, we found that loss of the C. elegans NCA channel increased arousal thresholds during L4/A lethargus sleep bouts and in-creased cation channel activity dein-creased sleep in this con-text. NCA channels acted upstream of egl-4 and gap junctions in lethargus sleep. Loss ofC. elegansPDF neuro-peptides or receptors had no effect on sleep quantity
during lethargus, but their loss slightly increased arousal thresholds during these sleep bouts. Our results suggest that inC. eleganslethargus sleep, PDF signaling likely does not act downstream of NCA channels. This unequivocally demonstrates a requirement for gap junction proteins in sleep and demonstrates thatEGL-4PKG regulation of sleep is dependent on gap junction function.
A recent forward genetic screen in mice for genes involved in sleep, identified a dominant, missense mutation in murine NALCN, which increases channel activity and leads to de-creased rapid eye movement sleep (Funatoet al.2016). Mu-rineNalcnencodes the direct ortholog ofC. elegansnca-1and nca-2. Our work inC. elegansfinds that increased NCA chan-nel activity decreases sleep bout quantity during lethargus, which is consistent with a requirement for NALCN channels in murine sleep (Funato et al. 2016). We speculate that if these channels act in wake-promoting neurons or sensory neurons, their loss might increase arousal thresholds during sleep. In Drosophila, PDF signaling and cation channels co-ordinately regulate Drosophilacircadian behavior, based on activity and entrainment to light/dark cycles (Zhang et al.
2010).DrosophilaPDF-expressing neurons play critical roles in activating neurons that express NCA channels, which in turn helps drive circadian behaviors. InC. elegans, PDF sig-naling regulates activity levels in adult animals and sup-presses sleep quantity defects in animals lacking neuropeptide Y signaling (Meelkop et al.2012; Choi et al.
2013). Our work confirms that loss of PDF signaling does not change C. elegans lethargus sleep quantity (Choiet al.
2013), but reveals that PDF signaling regulates arousal thresholds during sleep bouts, confirming a cross-species role for PDF signaling in arousal and sleep. We note that loss of nlf-1, which encodes an ER-localized protein required for NCA channel trafficking, did not perturb sleep or arousal thresholds (Figure S3, D and E). As the fainting locomotion defects innlf-1loss-of-function animals are less severe than those seen inncaloss-of-function animals (Xieet al.2013), NLF may also interact with or regulate additional, unidenti-fied channels that affect both locomotion and sleep, antago-nizing NCA channel function. Combined, these studies in disparate animal species highlight the conserved require-ment for NACLN channels in sleep, although the molecular mechanisms underlying sleep regulation may require further study.
Invertebrate gap junctions comprise innexin proteins, while vertebrate gap junctions comprise connexin proteins. Despite their analogous function in electrically coupling ad-jacent cells, innexin and connexin protein amino acid se-quences cannot be easily aligned, which makes drawing conclusions about orthology or orthologous function difficult (Abascal and Zardoya 2013). Vertebrates also have genes that encode innexin protein orthologs, called pannexins. Innexin and pannexin proteins share sequence similarity and both can function as hemichannels (Bruzzone et al.
Prior work is not inconsistent with a conserved role for gap junctions in sleep. Rapid eye movement sleep in rats is dis-rupted by quinine, a nonspecific blocker of gap junctions and various other channels and solute pumps (Franco-Pérez and Paz 2009; Connors 2012). Further, mice lacking Pannexin 1 function show increased wake and decreased slow-wave sleep, possibly due to the depleted extracellular adenosine (Kovalzon et al. 2017). Also, Connexin 43 knockout mice show excessive sleepiness and fragmented wakefulness, which may be attributed to connexin loss in astrocytes caus-ing impaired lactate shuttlcaus-ing that impairs orexin neuron function (Clasadonte et al. 2017). Additionally, Connexin
36 knockout mice show dampened circadian activity rhythms that may be attributed to loss of synchronized spiking in the suprachiasmatic nucleus (Longet al.2005). Finally, a recent study in Drosophila has demonstrated requirement for
in circuits, allowing normalC. eleganssleep. Or, the synchro-nized neuronal activity characteristic of sleep in many species may require gap junction function. Additional studies are clearly required to understand how gap junction proteins act to regulate sleep and arousal across species.
DoC. elegansUNC-7andUNC-9proteins function as hemi-channels or as gap junctions proteins in the context of C. eleganssleep? Previous work has established thatC. elegans
UNC-7may function as a hemichannel in some contexts, but must function as a gap junction for normal locomotion (Bouhourset al.2011). We do not directly test ifUNC-7acts as a hemichannel in the context of sleep. However, our over-expression data showed that only by overexpressing both UNC-7andUNC-9together were we able to induce sleep in adult animals. This suggests that either collaboration ofUNC-7 andUNC-9in heterotypic gap junctions is required for normal sleep, or that in the ectopic overexpression paradigm, high levels of bothUNC-7andUNC-9hemichannels are required.
During lethargus,C. elegansstop feeding and pharyngeal pumping ceases. Why pumping normally ceases during leth-argus is unclear. Previous work has demonstrated a require-ment forUNC-7in EGF-signaling induced pumping cessation during sleep bouts (Van Buskirk and Sternberg 2007). Fur-ther, a recent report found that optical silencing of body wall muscles can induce pumping inhibition in adult animals via UNC-7gap junctions between the I1 neuron and RIP neurons, the only intra- and extrapharyngeal neural connection (Takahashi and Takagi 2017). However, even in animals lackingunc-7function, we observed that pharyngeal pump-ing ceases durpump-ing lethargus, suggestpump-ing that other factors are at work. We note that lethargus-specific decreases in pharyn-geal muscle excitability may play a major role in suppressing pharyngeal pumping and feeding during lethargus, as optical stimulation of pharyngeal muscles cannot drive contraction (Trojanowskiet al.2016). Gap junction proteins may contrib-ute to cessation of feeding during lethargus, but other regu-latory pathways are likely major players.
Results presented here suggest that gap junctions function downstream of other proteins known to regulateC. elegans
sleep. Indeed,C. elegansgap junctions function downstream of the cGMP-dependent protein kinaseEGL-4in sleep, suggest-ing a model in whichEGL-4directly or indirectly regulates gap junction function. Gap junction activity could be regulated at multiple levels, including protein local expression, membrane protein endocytosis and exocytosis, and/or protein modifi ca-tions that might alter gap junction permeability (Kjensethet al.
2010; Axelsenet al.2013; Pogodaet al.2016). Phosphoryla-tion and redox changes can regulate connexin activity and trafficking (Pogoda et al.2016), but a connection between gap junction proteins and cGMP-dependent kinase activity has not been examined to our knowledge. The only exception is onein vitrostudy showing that nitric oxide can lead to PKG phosphorylation of connexins (Patelet al.2006). Further work examining how gap junctions are regulated might provide novel insights into how cGMP-dependent protein kinases reg-ulate sleep and arousal across species.
Acknowledgments
We thank Mei Zhen (Lunenfeld–Tanenbaum Research Insti-tute) and Dong Yan (Duke University) for numerous con-structs and strains, Gary Silverman and Stephen Pak (Washington University in St. Louis) and Barry Connors (Brown University) for manuscript discussion, and Aravinthan Samuel (Harvard University) for critical advice and support. Some strains were provided by the Caenorhabditis
Genetics Center, funded by National Institutes of Health Office of Research Infrastructure Programs (grant P40 OK010440). We acknowledge support from the Brown In-stitute for Brain Science/Norman Prince Neurosciences Institute (BIBS/NPNI) Postdoctoral Fellowship in Transla-tional Neuroscience (to H.H.), Karen T. Romer Undergrad-uate Teaching and Research Awards (to D.J.H. and L.L.S.), Burroughs Wellcome Fund Career Award (to V.V.) and National Institute of Neurological Disorders and Stroke (NINDS) grant NS055813 (to A.C.H.).
Literature Cited
Abascal, F., and R. Zardoya, 2013 Evolutionary analyses of gap junction protein families. Biochim. Biophys. Acta 1828: 4–14.
https://doi.org/10.1016/j.bbamem.2012.02.007
Allada, R., C. Cirelli, and A. Sehgal, 2017 Molecular mecha-nisms of sleep homeostasis inflies and mammals. Cold Spring Harb. Perspect. Biol. 9. https://doi.org/10.1101/cshperspect. a027730
Axelsen, L. N., K. Calloe, N. H. Holstein-Rathlou, and M. S. Nielsen, 2013 Managing the complexity of communication: regulation of gap junctions by post-translational modification. Front. Phar-macol. 4: 130.https://doi.org/10.3389/fphar.2013.00130
Banerjee, D., A. Kwok, S. Y. Lin, and F. J. Slack, 2005 Developmental timing in C. elegans is regulated by kin-20 and tim-1, homologs of core circadian clock genes. Dev. Cell 8: 287–295.https://doi.org/ 10.1016/j.devcel.2004.12.006
Boone, A. N., A. Senatore, J. Chemin, A. Monteil, and J. D. Spaf-ford, 2014 Gd3+ and calcium sensitive, sodium leak currents are features of weak membrane-glass seals in patch clamp re-cordings. PLoS One 9: e98808.https://doi.org/10.1371/journal. pone.0098808
Bouhours, M., M. D. Po, S. Gao, W. Hung, H. Liet al., 2011 A co-operative regulation of neuronal excitability by UNC-7 in-nexin and NCA/NALCN leak channel. Mol. Brain 4: 16.
https://doi.org/10.1186/1756-6606-4-16
Bruzzone, R., S. G. Hormuzdi, M. T. Barbe, A. Herb, and H. Monyer, 2003 Pannexins, a family of gap junction proteins expressed in brain. Proc. Natl. Acad. Sci. USA 100: 13644–13649.https:// doi.org/10.1073/pnas.2233464100
Campbell, S. S., and I. Tobler, 1984 Animal sleep: a review of sleep duration across phylogeny. Neurosci. Biobehav. Rev. 8: 269–300.https://doi.org/10.1016/0149-7634(84)90054-X
Chen, B., Q. Liu, Q. Ge, J. Xie, and Z. W. Wang, 2007 UNC-1 regulates gap junctions important to locomotion inC. elegans. Curr. Biol. 17: 1334–1339. https://doi.org/10.1016/j.cub. 2007.06.060
Cho, J. Y., and P. W. Sternberg, 2014 Multilevel modulation of a sensory motor circuit duringC. eleganssleep and arousal. Cell 156: 249–260.https://doi.org/10.1016/j.cell.2013.11.036
for behavioral quiescence in C. elegans. Neuron 78: 869–880.
https://doi.org/10.1016/j.neuron.2013.04.002
Cirelli, C., 2009 The genetic and molecular regulation of sleep: from fruit flies to humans. Nat. Rev. Neurosci. 10: 549–560.
https://doi.org/10.1038/nrn2683
Clasadonte, J., E. Scemes, Z. Wang, D. Boison, and P. G. Hay-don, 2017 Connexin 43-mediated astroglial metabolic networks contribute to the regulation of the sleep-wake cycle. Neuron 95: 1365–1380.e5. https://doi.org/10.1016/j.neuron. 2017.08.022
Connors, B. W., 2012 Tales of a dirty drug: carbenoxolone, gap junctions, and seizures. Epilepsy Curr. 12: 66–68. https://doi. org/10.5698/1535-7511-12.2.66
Evans, T. C., 2006 Transformation and microinjection (April 6, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.108.1, http://www. wormbook.org.
Flourakis, M., E. Kula-Eversole, A. L. Hutchison, T. H. Han, K. Arandaet al., 2015 A conserved bicycle model for circadian clock control of membrane excitability. Cell 162: 836–848.
https://doi.org/10.1016/j.cell.2015.07.036
Franco-Pérez, J., and C. Paz, 2009 Quinine, a selective gap junc-tion blocker, decreases REM sleep in rats. Pharmacol. Biochem. Behav. 94: 250–254.https://doi.org/10.1016/j.pbb.2009.09.003
Funato, H., C. Miyoshi, T. Fujiyama, T. Kanda, M. Sato et al., 2016 Forward-genetics analysis of sleep in randomly muta-genized mice. Nature 539: 378–383.https://doi.org/10.1038/ nature20142
Gao, S., L. Xie, T. Kawano, M. D. Po, S. Guanet al., 2015 The NCA sodium leak channel is required for persistent motor circuit ac-tivity that sustains locomotion. Nat. Commun. 6: 6323 (erratum: Nat. Commun. 6: 7191).https://doi.org/10.1038/ncomms7323
Ghosh, R., and S. W. Emmons, 2008 Episodic swimming behavior in the nematode C. elegans. J. Exp. Biol. 211: 3703–3711.
https://doi.org/10.1242/jeb.023606
Hart, A. C., S. Sims, and J. M. Kaplan, 1995 Synaptic code for sensory modalities revealed byC. elegansGLR-1 glutamate re-ceptor. Nature 378: 82–85.https://doi.org/10.1038/378082a0
Hill, A. J., R. Mansfield, J. Lopez, D. M. Raizen, and C. Van Buskirk, 2014 Cellular stress induces a protective sleep-like state inC. elegans. Curr. Biol. 24: 2399–2405. https://doi.org/10.1016/j. cub.2014.08.040
Huang, H., K. Singh, and A. C. Hart, 2017a Measuring
Caenorhabditis elegans sleep during the transition to adulthood using a microfluidics-based system. Bio Protoc. 7: e2174.
https://doi.org/10.21769/BioProtoc.2174
Huang, H., C. T. Zhu, L. L. Skuja, D. J. Hayden, and A. C. Hart, 2017b Genome-wide screen for genes involved inCaenorhabditis elegansdevelopmentally timed sleep. G3 (Bethesda) 7: 2907–2917.
https://doi.org/10.1534/g3.117.300071
Huang, L. S., and P. W. Sternberg, 1995 Genetic dissection of developmental pathways. Methods Cell Biol. 48: 97–122.
https://doi.org/10.1016/S0091-679X(08)61385-0
Iwanir, S., N. Tramm, S. Nagy, C. Wright, D. Ishet al., 2013 The microarchitecture ofC. elegans behavior during lethargus: ho-meostatic bout dynamics, a typical body posture, and regulation by a central neuron. Sleep (Basel) 36: 385–395. https://doi. org/10.5665/sleep.2456
Kawano, T., M. D. Po, S. Gao, G. Leung, W. S. Ryuet al., 2011 An imbalancing act: gap junctions reduce the backward motor circuit activity to bias C. elegans for forward locomotion. Neuron 72: 572–586. https://doi.org/10.1016/j.neuron.2011. 09.005
Kjenseth, A., T. Fykerud, E. Rivedal, and E. Leithe, 2010 Regulation of gap junction intercellular communication by the ubiquitin sys-tem. Cell. Signal. 22: 1267–1273. https://doi.org/10.1016/j.cell-sig.2010.03.005
Kovalzon, V. M., L. S. Moiseenko, A. V. Ambaryan, S. Kurtenbach, V. I. Shestopalovet al., 2017 Sleep-wakefulness cycle and be-havior in pannexin1 knockout mice. Behav. Brain Res. 318: 24– 27.https://doi.org/10.1016/j.bbr.2016.10.015
Langmesser, S., P. Franken, S. Feil, Y. Emmenegger, U. Albrecht
et al., 2009 cGMP-dependent protein kinase type I is impli-cated in the regulation of the timing and quality of sleep and wakefulness. PLoS One 4: e4238. https://doi.org/10.1371/ journal.pone.0004238
Lear, B. C., J. M. Lin, J. R. Keath, J. J. McGill, I. M. Ramanet al., 2005 The ion channel narrow abdomen is critical for neural output of the Drosophila circadian pacemaker. Neuron 48: 965– 976.https://doi.org/10.1016/j.neuron.2005.10.030
Lee, J. H., L. L. Cribbs, and E. Perez-Reyes, 1999 Cloning of a novel four repeat protein related to voltage-gated sodium and calcium channels. FEBS Lett. 445: 231–236. https://doi.org/ 10.1016/S0014-5793(99)00082-4
Liebeskind, B. J., D. M. Hillis, and H. H. Zakon, 2012 Phylogeny unites animal sodium leak channels with fungal calcium chan-nels in an ancient, voltage-insensitive clade. Mol. Biol. Evol. 29: 3613–3616.https://doi.org/10.1093/molbev/mss182
Long, M. A., M. J. Jutras, B. W. Connors, and R. D. Burwell, 2005 Electrical synapses coordinate activity in the suprachias-matic nucleus. Nat. Neurosci. 8: 61–66. https://doi.org/ 10.1038/nn1361
Meelkop, E., L. Temmerman, T. Janssen, N. Suetens, I. Beetset al., 2012 PDF receptor signaling inCaenorhabditis elegans modu-lates locomotion and egg-laying. Mol. Cell. Endocrinol. 361: 232–240.https://doi.org/10.1016/j.mce.2012.05.001
Meng, L., C. H. Chen, and D. Yan, 2016 Regulation of gap junc-tion dynamics by UNC-44/ankyrin and UNC-33/CRMP through VAB-8 in C. elegans neurons. PLoS Genet. 12: e1005948.
https://doi.org/10.1371/journal.pgen.1005948
Mok, D. Z., P. W. Sternberg, and T. Inoue, 2015 Morphologically defined sub-stages ofC. elegansvulval development in the fourth larval stage. BMC Dev. Biol. 15: 26. https://doi.org/10.1186/ s12861-015-0076-7
Monsalve, G. C., C. Van Buskirk, and A. R. Frand, 2011 LIN-42/ PERIOD controls cyclical and developmental progression of
C. elegans molts. Curr. Biol. 21: 2033–2045. https://doi.org/ 10.1016/j.cub.2011.10.054
Nichols, A. L. A., T. Eichler, R. Latham, and M. Zimmer, 2017 A global brain state underlies C. eleganssleep behavior. Science 356: eaam6851.https://doi.org/10.1126/science.aam6851
Park, E. C., and H. R. Horvitz, 1986 Mutations with dominant effects on the behavior and morphology of the nematode
Caenorhabditis elegans. Genetics 113: 821–852.
Patel, L. S., C. K. Mitchell, W. P. Dubinsky, and J. O’Brien, 2006 Regulation of gap junction coupling through the neuronal connexin Cx35 by nitric oxide and cGMP. Cell Commun. Adhes. 13: 41–54.https://doi.org/10.1080/15419060600631474
Pogoda, K., P. Kameritsch, M. A. Retamal, and J. L. Vega, 2016 Regulation of gap junction channels and hemichannels by phosphorylation and redox changes: a revision. BMC Cell Biol. 17: 11.https://doi.org/10.1186/s12860-016-0099-3
Raizen, D. M., J. E. Zimmerman, M. H. Maycock, U. D. Ta, Y. J. You
et al., 2008 Lethargus is a Caenorhabditis elegans sleep-like state. Nature 451: 569–572 (erratum: Nature 453: 952).https:// doi.org/10.1038/nature06535
Ren, D., 2011 Sodium leak channels in neuronal excitability and rhythmic behaviors. Neuron 72: 899–911. https://doi.org/ 10.1016/j.neuron.2011.12.007
Schwarz, J., I. Lewandrowski, and H. Bringmann, 2011 Reduced activity of a sensory neuron during a sleep-like state in
Caenorhabditis elegans. Curr. Biol. 21: R983–R984.https://doi. org/10.1016/j.cub.2011.10.046
Sedensky, M. M., and P. M. Meneely, 1987 Genetic analysis of halothane sensitivity in Caenorhabditis elegans. Science 236: 952–954.https://doi.org/10.1126/science.3576211
Senatore, A., and J. D. Spafford, 2013 A uniquely adaptable pore is consistent with NALCN being an ion sensor. Channels (Austin) 7: 60–68.https://doi.org/10.4161/chan.23981
Senatore, A., A. Monteil, J. van Minnen, A. B. Smit, and J. D. Spafford, 2013 NALCN ion channels have alternative selectivityfilters resembling calcium channels or sodium channels. PLoS One 8: e55088.https://doi.org/10.1371/journal.pone.0055088
Singh, K., M. Y. Chao, G. A. Somers, H. Komatsu, M. E. Corkins
et al., 2011 C. elegansNotch signaling regulates adult chemo-sensory response and larval molting quiescence. Curr. Biol. 21: 825–834.https://doi.org/10.1016/j.cub.2011.04.010
Singh, K., J. Y. Ju, M. B. Walsh, M. A. DiIorio, and A. C. Hart, 2014 Deep conservation of genes required for bothDrosophila melanogasterandCaenorhabditis eleganssleep includes a role for dopaminergic signaling. Sleep (Basel) 37: 1439–1451.https:// doi.org/10.5665/sleep.3990
Starich, T. A., R. K. Herman, and J. E. Shaw, 1993 Molecular and genetic analysis of unc-7, aCaenorhabditis elegansgene required for coordinated locomotion. Genetics 133: 527–541.
Starich, T. A., R. Y. Lee, C. Panzarella, L. Avery, and J. E. Shaw, 1996 eat-5 and unc-7 represent a multigene family in
Caenorhabditis elegansinvolved in cell-cell coupling. J. Cell Biol. 134: 537–548.https://doi.org/10.1083/jcb.134.2.537
Starich, T. A., J. Xu, I. M. Skerrett, B. J. Nicholson, and J. E. Shaw, 2009 Interactions between innexins UNC-7 and UNC-9 medi-ate electrical synapse specificity in the Caenorhabditis elegans
locomotory nervous system. Neural Dev. 4: 16. https://doi. org/10.1186/1749-8104-4-16
Takahashi, M., and S. Takagi, 2017 Optical silencing of body wall muscles induces pumping inhibition in Caenorhabditis elegans. PLoS Genet. 13: e1007134. https://doi.org/10.1371/journal. pgen.1007134
Trojanowski, N. F., and D. M. Raizen, 2016 Call it worm sleep. Trends Neurosci. 39: 54–62. https://doi.org/10.1016/j. tins.2015.12.005
Trojanowski, N. F., M. D. Nelson, S. W. Flavell, C. Fang-Yen, and D. M. Raizen, 2015 Distinct mechanisms underlie quiescence dur-ing twoCaenorhabditis eleganssleep-like states. J. Neurosci. 35: 14571–14584. https://doi.org/10.1523/JNEUROSCI.1369-15.2015
Trojanowski, N. F., D. M. Raizen, and C. Fang-Yen, 2016 Pharyngeal pumping inCaenorhabditis elegansdepends on tonic and phasic signaling from the nervous system. Sci. Rep. 6: 22940.https://doi. org/10.1038/srep22940
Troup, M., M. H. Yap, C. Rohrscheib, M. J. Grabowska, D. Ertekin
et al., 2018 Acute control of the sleep switch in Drosophila reveals a role for gap junctions in regulating behavioral re-sponsiveness. eLife 7: e37105.https://doi.org/10.7554/eLife. 37105
Van Buskirk, C., and P. W. Sternberg, 2007 Epidermal growth factor signaling induces behavioral quiescence inCaenorhabditis elegans. Nat. Neurosci. 10: 1300–1307.https://doi.org/10.1038/ nn1981
van der Linden, A. M., M. Beverly, S. Kadener, J. Rodriguez, S. Wasserman et al., 2010 Genome-wide analysis of light- and temperature-entrained circadian transcripts in Caenorhabditis elegans. PLoS Biol. 8: e1000503.https://doi.org/10.1371/journal. pbio.1000503
Venkatachalam, V., N. Ji, X. Wang, C. Clark, J. K. Mitchell et al., 2016 Pan-neuronal imaging in roaming Caenorhabditis ele-gans. Proc. Natl. Acad. Sci. USA 113: E1082–E1088.https:// doi.org/10.1073/pnas.1507109113
Xie, L., S. Gao, S. M. Alcaire, K. Aoyagi, Y. Wanget al., 2013 NLF-1 delivers a sodium leak channel to regulate neuronal excitabil-ity and modulate rhythmic locomotion. Neuron 77: 1069–1082.
https://doi.org/10.1016/j.neuron.2013.01.018
Yeh, E., S. Ng, M. Zhang, M. Bouhours, Y. Wanget al., 2008 A putative cation channel, NCA-1, and a novel protein, UNC-80, transmit neuronal activity in C. elegans. PLoS Biol. 6: e55.
https://doi.org/10.1371/journal.pbio.0060055
Zhang, L., B. Y. Chung, B. C. Lear, V. L. Kilman, Y. Liu et al., 2010 DN1(p) circadian neurons coordinate acute light and PDF inputs to produce robust daily behavior in
Drosophila. Curr. Biol. 20: 591–599. https://doi.org/10.1016/ j.cub.2010.02.056