Visual illusion of tool use recalibrates tactile perception
Full text
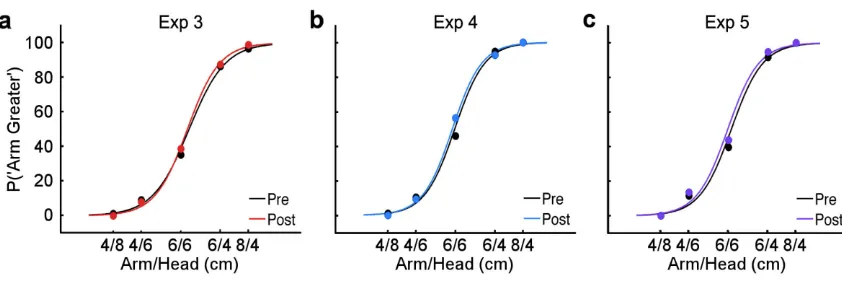
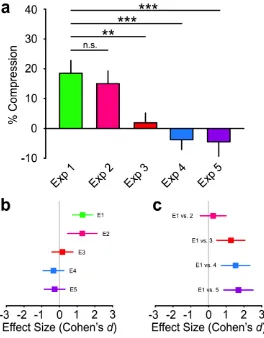
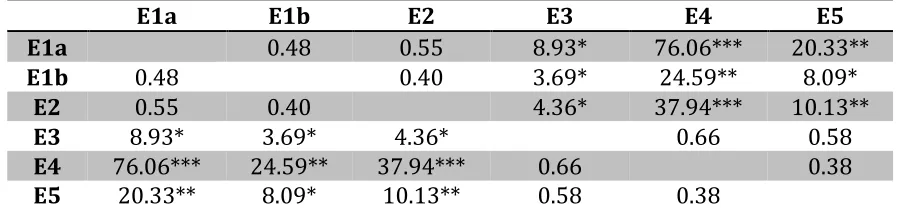
Figure


Related documents
This study used a self-reported questionnaire from all participants working in the six selected PHC centres; however, semi-structured interviews were only conducted with nurses who
Additional data file 2 contains data showing that double RNAi by soaking worms in dsRNA for two genes at once is an inefficient means of synthetic genetic analysis. Additional data
ACF: Ambuja Cement Foundation; ASHA: Accredited social health activist; CHW: Community health workers; CSR: Corporate social responsibility; FGD: Focused group discussion;
It may be stated that a very old serum containing an anti-E agglutinin has been tested which gave specific agglutination of homologous cells by the ordinary test, aud yet our
The objectives of the study were to identify the more-effective and less-effective schools; to find out the differences between more-effective and less-effective
Using a literature search, we examined (1) the frequency of SBCs resulting in fatal injuries; (2) the frequency of SBCs resulting in severe non-fatal injuries (here de fi ned
In the latter, an external electron beam from a high quality source, such as a con- ventional accelerator, is injected into another plasma stage where it is further accelerated.. In
1.1 The Task Team on Forecaster Competencies & Training (TT-TRG) was formed as part of the Working Group on Weather Services (WG-WXS) in April 2011, following the 15th session
