0095-1137/05/$08.00⫹0 doi:10.1128/JCM.43.7.3283–3289.2005
Characterization of Vancomycin-Resistant
Enterococcus faecium
Isolates from Broiler Poultry and Pig Farms in England
and Wales
L. Garcia-Migura, E. Pleydell, S. Barnes, R. H. Davies,
and E. Liebana*
Department of Food and Environmental Safety, Veterinary Laboratories Agency-Weybridge, Addlestone, Surrey KT15 3NB, United Kingdom
Received 29 July 2004/Returned for modification 29 November 2004/Accepted 24 February 2005
This study aimed to investigate the occurrence and molecular epidemiology of vancomycin-resistant
Entero-coccus faecium(VREF) isolates on poultry and pig farms in England and Wales. A total of 217 VREF isolates
were obtained from fresh feces and environmental swabs collected from conventional and organic farms. A predominant pulsed-field gel electrophoresis (PFGE) profile was found for each VREF-positive farm, together with less frequent types. All isolates presented thevanAgenotype and wereespnegative. Seventy-six percent of the VREF isolates were additionally resistant to nine or more antimicrobials, presenting a diverse range of resistance phenotypes. The multiresistance traits did not appear to be specific to individual farms or sample types (i.e., environmental or fecal), nor did they correlate with any specific PFGE type. Ninety-three percent of the isolates were resistant to penicillin, 89% were resistant to tetracycline, 87.5% were resistant to erythro-mycin, and 50% were resistant to quinupristin-dalfospristin (Synercid). The lack of clonality among these populations may suggest the horizontal transfer of resistance genes and/or a dynamic replacement of clonal lines rather than persistence.
Bacteria are capable of developing strategies to enhance their survival in adverse situations such as conditions of high salinity, changes in temperature, the presence of heavy metals, or the presence of antimicrobial activity.Enterococcus faecium, which is part of the normal intestinal flora in humans and animals (12), provides a good example of such adaptive bac-terial evolution. This species has emerged as an important opportunistic pathogen causing life-threatening infections in hospitals. The emergence of this pathogen is associated with a remarkable capacity to accumulate resistance to antimicrobials (13). Enterococci are intrinsically resistant to cephalosporins and aminoglycosides and were the first bacteria to acquire resistance to vancomycin (19), a glycopeptide antibiotic com-monly used for the clinical therapy of gram-positive nosoco-mial infections.
Multidrug-resistant enterococci, particularly those that are vancomycin resistant, are a major cause of concern for the medical community. The percentage of infections caused by vancomycin-resistant enterococci (VRE) has increased 20-fold over the last decade in the United States (13). Recently, it was shown that the genes responsible for this resistance have the potential to be transferred to other gram-positive pathogens such asStaphylococcus aureus(20), thus intensifying the public health threat, especially now that the first cases of vancomycin-and methicillin-resistantS. aureushave been reported in the United States (26, 30).
Antimicrobial growth promoters have been used for the past
five decades as an effective way of enhancing productivity and animal health during livestock production. Avoparcin is an example of a feed additive that has been used in intensive and integrated agricultural systems across Europe, especially within the pig and poultry industries. This growth promoter has not been used in the United States.
Avoparcin is a glycopeptide that induces cross-resistance to vancomycin (32) and has been considered an important factor in the emergence and spread of resistance to vancomycin in enterococcal populations (4). For this reason, the use of avoparcin was banned in Denmark in 1995 and throughout the European Union in 1997.
In Europe, VRE carriage occurs mainly among healthy in-dividuals in the community and in farm livestock (8). In con-trast, in the United States VRE are normally endemic to hos-pitals (8, 10, 15). VRE have also recently been found as a result of infections linked to foreign travel and to the consumption of imported food (17). It has been postulated that in Europe, the colonization of humans by VRE may occur predominantly via the food chain (5, 6). Moreover, similarvanAelements have been found in strains from animals and humans (9, 25, 29). Although the colonization of humans by animal isolates might be “transient” (7, 24), the risk of transfer of resistance genes during the colonization period has not yet been assessed and might be crucial.
Multiresistant organisms might enter the food chain via farm animals. The aim of the present study was to investigate the occurrence and molecular epidemiology of vancomycin-resis-tantE. faecium(VREF) on poultry and pig farms in England and Wales. The molecular characterization of isolates col-lected from farms (including genotyping and identification of specific resistance determinants) will help to develop a better
* Corresponding author. Mailing address: Department of Food and Environmental Safety, Veterinary Laboratories Agency-Weybridge, Addlestone, Surrey KT15 3NB, United Kingdom. Phone: 44 1932 357587. Fax: 44 1932 357595. E-mail: [email protected].
3283
on May 15, 2020 by guest
http://jcm.asm.org/
understanding of the persistence and dynamics of these organ-isms in farm environments.
MATERIALS AND METHODS
Farms and sample collection.Samples were collected from a total of 47 farms. Twenty-five farms (six conventional and seven organic broiler farms and seven conventional and five organic pig farms) were sampled once in 2002 and then again within the same month in 2003. A small number of farms were visited more frequently in order to make preliminary assessments of short-term variability.
During each visit, 40 to 60 pooled fecal samples were collected, ensuring that all age groups or production classes present on each farm were represented. Pooled fecal samples were collected from the ground and consisted of approx-imately 5 g of fecal material from each of eight fresh fecal masses. Care was taken during the procedure to avoid environmental contamination of the fecal samples. Representative environmental swabs were also taken from internal building surfaces, feeders, and drinkers. For the remaining 22 farms, the farmers collected eight pooled fecal samples from three of their livestock houses on a single occasion and sent them by post to the laboratory for analysis.
Bacteriology. Fecal samples (40 g) were mixed with the same volume of phosphate-buffered saline and vortexed, and then 10l of each was streaked onto Slanetz and Bartley agar (Oxoid, Basingstoke, United Kingdom) containing vancomycin (6g/ml) and incubated aerobically for 48 h at 42°C. Environmental samples were enriched in buffered peptone water for 18 h at 37°C before plating onto the same selective medium. Presumptive VREF isolates were selected by their colony morphology and color (purple-pink colonies with a lighter halo), subcultured, and stored at⫺70°C. The identity of the presumptiveE. faecium
isolates was confirmed by real-time PCR. Amplification was performed in a LightCycler instrument (Roche Diagnostics UK Ltd., Lewes, United Kingdom) in glass capillaries containing 20-l reaction mixtures including 1⫻LightCycler FastStart DNA Master SYBR green I (Roche Molecular Biochemicals), 10 pmol each of theddlE. faecium-specific primers F1 and F2 (11), 4 mM MgCl2(final concentration), and 2l of a crude extract of bacterial DNA template. The PCR run comprised 30 cycles with denaturation at 95°C for 5 seconds, annealing at 50°C for 5 seconds, and extension at 72°C for 22 seconds. The nature of the amplicon was determined by a melting point analysis over a temperature range from 65 to 95°C, with a transition rate of 0.1°C/s and continuous detection of fluorescence in channel 1. The melting temperature (Tm) for the PCR product
was 84°C, and the size of the product was 550 bp.
The identification of the genes responsible for vancomycin resistance was investigated by a multiplex PCR as described previously (11). Subsequently, a duplex, SYBR green I-based PCR assay was developed for the simultaneous detection ofE. faeciumandvanAgenes by the use of the primers described above in a LightCycler assay. The PCR consisted of 30 cycles of 95°C for 5 seconds (denaturation), 54°C for 5 seconds (annealing), and 72°C for 25 seconds (exten-sion). A melting point analysis over a temperature range from 65 to 95°C with a transition rate of 0.1°C/s resulted in the identification of two distinct peaks representing the two targets, i.e., one at 84°C forddlE. faeciumand the other at
87°C forvanA.
The presence of theespgene in 137 randomly selected isolates was assessed by PCR. Primers were designed with DnaStar software from a published sequence of theespgene ofE. faecium(GenBank database accession number AX537383). PCRs were prepared as follows: 25-l volumes contained 10 pmol of each primer (esp1, 5⬘TTAGCGGGAACAGGTCACA; esp2, 5⬘TGTTGCATCATTTTCCA TAGC), 1.5 mM MgCl2, 1 U of AmpliTaq Gold (Applied Biosystems), 1⫻ GeneAmp PCR buffer, and 5l of template DNA. The PCR cycling parameters were as follows: denaturation at 94°C for 10 min, followed by 30 cycles consisting of 94°C for 30 seconds, 62°C for 1 min, and 72°C for 3 min and a final cycle of 10 min at 72°C. The expected size of the amplicon was 471 bp.
PFGE.DNAs were prepared as described previously (9a). A single colony of each isolate was streaked onto yeast extract agar and incubated overnight at 37°C. Using a cotton swab, we transferred a portion of the growth to 3 ml of TE buffer (10 mM Tris, 1 mM EDTA, pH 8.0) and adjusted the cell concentration to 0.59 with a Dade Microscan turbidity meter (Dade Behring). A total volume of 240l of the suspension was transferred to 1.5-ml microcentrifuge tubes, and 60l of lysozyme solution (10 mg/ml) was added. The tubes were incubated at 37°C for 10 min. Immediately after incubation, 300l of 1.2% Seakem Gold agarose (Cambrex, East Rutherford, N.J.)–1% sodium dodecyl sulfate–0.2 mg/ml proteinase K was mixed with the bacterial suspension and pipetted into dispos-able plug moulds. Three plugs per isolate were transferred to 50-ml polypro-pylene screw-top tubes with 5 ml of cell lysis buffer (50 mM Tris, 50 mM EDTA, pH 8.0, 1% sarcosyl, 0.15 mg/ml proteinase K) and then incubated at 54°C in a shaking water bath for 2 h. Thereafter, the plugs were washed twice with 15 ml
of sterile water and four times with TE buffer at 50°C for 15 min. Restriction digestion of chromosomal DNAs was carried out by using 25 units of SmaI (Promega, Southampton, United Kingdom) for 2 h at 25°C. Pulsed-field gel electrophoresis (PFGE) was performed on a CHEF DRIII system (Bio-Rad, Hercules, Calif.) in 0.5⫻TBE extended-range buffer (Bio-Rad) with recircula-tion at 14°C. DNA restricrecircula-tion fragments were resolved in 0.8% SeaKem Gold agarose in 0.5⫻TBE buffer. DNA fromSalmonellaBraenderup H9812 restricted with XbaI was used as a size marker. Restriction fragments were resolved under the running conditions described by Turabelidze et al. (28). Macrorestriction patterns were compared by the use of BioNumerics software (Applied Maths, Sint-Martens-Latem, Belgium).
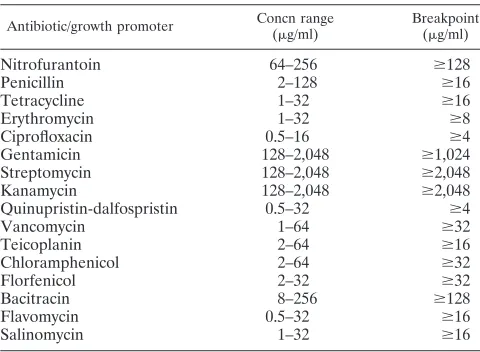
Antimicrobial susceptibility testing.Resistance patterns and MICs were as-certained for 16 different therapeutic and growth-promoting antimicrobials by a standard Sensititre protocol (Trek Diagnostic Systems Ltd., England). Table 1 gives the concentrations of antimicrobials tested and the breakpoint used for each drug (DANMAP 2002). Briefly, isolates were grown in yeast extract agar. Subsequently, a 0.5 McFarland cell suspension was prepared in demineralized water, and 10l was inoculated into 10 ml of cation-adjusted Mueller-Hinton broth (Trek Diagnostic Systems, Ohio) for a final inoculum of 105
CFU/ml. Aliquots of 50l of the inoculum were seeded in each well of a microtiter plate, which contained doubling dilutions of the antimicrobials. The plates were sealed and incubated aerobically at 37°C for 24 h. A growth control well was used as a reference for interpreting the growth patterns in each plate.Pseudomonas aerugi-nosa(ATCC 27853),Enterococcus faecalis(ATCC 29213),S. aureus(ATCC 29213), andEscherichia coli(ATCC 25922 and ATCC 35218) were used as quality control organisms. The MIC was recorded as the lowest concentration of antimicrobial that inhibited visible growth.
RESULTS
A total of 217 VREF isolates were obtained from fecal and environmental samples during the study period (January 2002 to February 2003).
When comparing the sensitivities of the two different sam-pling techniques (samsam-pling by farmers versus samsam-pling by the research team) for the detection of VREF by using the un-pairedttest, we found no evidence to reject the null hypothesis that there was no difference between the two collection meth-ods (P⫽0.457).
[image:2.585.301.541.88.264.2]Broiler farms. Twenty-seven of the 33 farms investigated were found to have at least one VREF-positive sample on at least one occasion over the course of the study. All 20 conven-tional broiler farms that submitted pooled fecal samples by post were found to be positive for VREF. The number of fecal samples from these farms which contained VREF ranged from
TABLE 1. Antimicrobials tested and breakpoints used for this study
Antibiotic/growth promoter Concn range (g/ml)
Breakpoint (g/ml)
Nitrofurantoin 64–256 ⱖ128
Penicillin 2–128 ⱖ16
Tetracycline 1–32 ⱖ16
Erythromycin 1–32 ⱖ8
Ciprofloxacin 0.5–16 ⱖ4
Gentamicin 128–2,048 ⱖ1,024
Streptomycin 128–2,048 ⱖ2,048
Kanamycin 128–2,048 ⱖ2,048
Quinupristin-dalfospristin 0.5–32 ⱖ4
Vancomycin 1–64 ⱖ32
Teicoplanin 2–64 ⱖ16
Chloramphenicol 2–64 ⱖ32
Florfenicol 2–32 ⱖ32
Bacitracin 8–256 ⱖ128
Flavomycin 0.5–32 ⱖ16
Salinomycin 1–32 ⱖ16
on May 15, 2020 by guest
http://jcm.asm.org/
one of eight to eight of eight. Of the further 13 farms that were visited in person, 35 to 100% of fecal samples from three of the six conventional broiler farms were found to be positive for VREF on every visit. Environmental swabs taken on these farms were also positive. On one conventional farm, VREF isolates were found in environmental swabs collected from the surfaces of two houses that had been cleaned and disinfected, although the organisms had not been found in fecal samples collected from the previous flock. Fecal VREF isolates were also found on three visits to two organic farms, but at a lower prevalence than that for the conventional farms (3 to 10% of fecal samples). On another organic farm, VREF isolates were found in environmental samples, but not in fecal samples, on one of five visits.
Pig farms. Fewer VREF isolates were isolated from pig farms in this study, although 4 of the 14 farms investigated did yield at least one positive sample. The two conventional fin-isher units that sent pooled fecal samples to the laboratory were found to have one of eight and five of eight positive samples. Fecal samples from 2 of the 12 farms that were sam-pled more intensively were positive for VREF at low levels (2 to 5%) on a single visit. One of these farms was conventional and the other was organic. We failed to find any VREF on a second visit to every one of those 12 farms.
Genotyping. All VREF isolates harbored the vanA resis-tance gene and all selected isolates wereespnegative. They all exhibited MICs of⬎64g/ml of vancomycin and between 8
g/ml andⱖ64g/ml of teicoplanin. One hundred nineteen PFGE restriction profiles were identified among the 217 iso-lates by SmaI-PFGE (Fig. 1). Each profile was given a unique identification number. The number of fragments generated ranged from 16 to 24, and their sizes varied from 20 to 350 kb. Our analysis revealed a predominant PFGE profile for each farm together with less represented types. The most prevalent type from one visit was never detected again in isolates from a second visit.
Two isolates from farms 3 and 8 were highly related accord-ing to PFGE. In order to check the stability of the PFGE types, we repeatedly passaged one colony of each of two presump-tively related isolates in the laboratory. PFGE was then re-peated with the new subcultures. During the third passage, a new band appeared in the PFGE profile of one of the isolates. This suggests that small changes in PFGE profiles may occur in short periods of time.
Antimicrobial resistance.Ninety-three percent of the VREF isolates were resistant to penicillin (MIC, ⱖ16g/ml), 89% were resistant to tetracycline (MIC, ⱖ16g/ml), 88% were resistant to erythromycin (MIC,ⱖ8g/ml), 79% were resistant to streptomycin (MIC,ⱖ2,048g/ml), and 50% were resistant to quinupristin-dalfospristin (Synercid) (MIC, 4 to 32g/ml). One isolate displayed resistance to chloramphenicol, with an MIC of 32g/ml, and 8% of the isolates were found to be of intermediate resistance (MIC, 16g/ml). None of the strains exhibited high-level resistance to gentamicin, and all were sen-sitive to florfenicol (MIC,ⱖ32g/ml). With the exception of vancomycin and teicoplanin, no correlation among resistances to several antimicrobials was found in this study.
An analysis of the resistance profiles indicated that 166 (76%) of the 217 VREF isolates tested were resistant to nine or more antimicrobials, comprising a diverse range of
pheno-types (n⫽70). Eleven percent of the isolates were resistant to the same 12 antimicrobials, as follows: bacitracin (MIC,⬎256
g/ml), ciprofloxacin (MIC, 4g/ml), erythromycin (MIC,⬎32
g/ml), flavomycin (MIC, ⱖ32 g/ml), kanamycin (MIC,
⬎2,048g/ml), nitrofurantoin (MIC, 128 to 256g/ml), pen-icillin (MIC, 16 to 128 g/ml), streptomycin (MIC, ⱖ2,048
g/ml), quinupristin-dalfospristin (MIC, 4 to 16g/ml), tetra-cycline (MIC,⬎32g/ml), vancomycin (MIC,⬎64g/ml), and teicoplanin (MIC,ⱖ64g/ml). These 24 isolates were all ob-tained from different conventional poultry farms. Finally, 23% of the isolates were resistant to 11 antimicrobials, 25% were resistant to 10, 17% were resistant to 9, and 11.4% were resis-tant to 8.
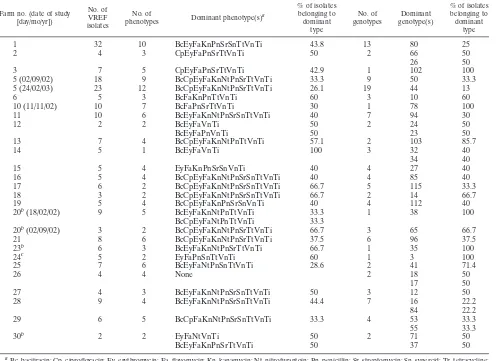
Table 2 shows the distribution of genotypes and phenotypes on farms with more than one VREF isolate.
DISCUSSION
VREF was found in samples from 31 of 47 farms (66%). However, we identified differences in the percentages of pos-itive pooled fecal samples between pig and poultry farms. Fur-thermore, VREF isolates were repeatedly isolated with a rel-atively high sample prevalence from conventional broiler farms. Far fewer samples from organic poultry yielded VREF, and VREF was never isolated from any organic broiler farm on more than one occasion. In the United Kingdom, certified organic farms must agree to relatively restrictive policies in terms of antimicrobial usage (http://www.defra.gov.uk/farm/or-ganic/legislation-standards). For instance, organic farms are not permitted to use antimicrobial growth-promoting agents, but they may administer therapeutic treatment if this is judged to be clinically necessary by a veterinary surgeon. None of the participating organic poultry farms used any antimicrobials during this study, and most of them (6/7) had not used such treatment within the 12 months preceding the study.
On one conventional farm, VREF was detected in samples originating from floors, walls, wooden roof supports, and dust taken after cleaning and disinfection. This indicates that insuf-ficient cleaning and disinfection may play a role in the persis-tence of these organisms. Interestingly, all isolates from this farm (which used no therapeutic antimicrobials but did use in-feed avilamycin) were of a single clone. The same pattern was also detected in a positive nondomestic bird sample col-lected from the concrete concourse outside the houses and in dust samples from an occupied house collected during the same visit. Suitable routine hygiene protocols should be per-formed thoroughly to prevent the persistence of these organ-isms on surfaces and to minimize colonization by resistant strains when new flocks enter the premises.
Multidrug-resistant VREF has been described previously (1, 2, 14), but the level of multiresistance among isolates found in the present study appears to be unprecedented. To date, such multiresistant profiles have not been reported elsewhere in Europe for animal sources. It is difficult to evaluate the signif-icance of these differences since the use of antimicrobials is a common practice in most European countries. It is also diffi-cult to assess the significance of potential horizontal gene transfer between other bacteria andE. faeciumclones in our study, and this would require a detailed characterization of the different genetic elements involved.
on May 15, 2020 by guest
http://jcm.asm.org/
on May 15, 2020 by guest
http://jcm.asm.org/
From the medical point of view, the emergence of multire-sistance among VREF isolates is a great cause for concern, since some of the antimicrobials involved are commonly used for the treatment of human VRE infections. This is well illus-trated by the examples of gentamicin and streptomycin, which have a synergistic effect when administered with cell wall in-hibitors such as vancomycin (18). A high-level resistance to aminoglycosides might pose a serious risk in hospitals, as an-timicrobial therapy could be limited. In this study, while no isolates were resistant to high levels of gentamicin, 70% were resistant to high levels of streptomycin. A combination of streptogramins (quinupristin-dalfospristin [Synercid]) has been successfully used for the treatment of VREF infections (27). However, someE. faeciumstrains have already acquired resistance to these antibiotics. This resistance may be related in
part to the use of virginiamycin, a growth-promoting feed ad-ditive incorporated in agriculture for poultry and pig produc-tion (32). The use of virginiamycin was banned in Denmark in 1998 and in the rest of the European Union in 1999. In the present study, 50% of the isolates were resistant to quinupris-tin-dalfospristin. Erythromycin resistance was also found at very high levels on every farm. Macrolide resistance encoded byermB-type genes has been linked to the same conjugative plasmid harboringvanAgenes (3). Further studies are being carried out to ascertain the possible role of gene linkage and the coselection of VREF on farms in the United Kingdom as a result of the use of macrolides.
The multiresistance profiles found in this study do not seem to be specific to a particular farm or sample type, and they do not correlate with any specific PFGE banding pattern.
Al-TABLE 2. Distribution of dominant phenotypes and genotypes on farms
Farm no. (date of study [day/mo/yr])
No. of VREF isolates
No. of
phenotypes Dominant phenotype(s)
a
% of isolates belonging to dominant
type
No. of genotypes
Dominant genotype(s)
% of isolates belonging to dominant
type
1 32 10 BcEyFaKnPnSrSnTtVnTi 43.8 13 80 25
2 4 3 CpEyFaPnSrTtVnTi 50 2 66 50
26 50
3 7 5 CpEyFaPnSrTtVnTi 42.9 1 102 100
5 (02/09/02) 18 9 BcCpEyFaKnNtPnSrTtVnTi 33.3 9 50 33.3
5 (24/02/03) 23 12 BcCpEyFaKnNtPnSrTtVnTi 26.1 19 44 13
6 5 3 BcFaKnPnTtVnTi 60 3 10 60
10 (11/11/02) 10 7 BcFaPnSrTtVnTi 30 1 78 100
11 10 6 BcEyFaKnNtPnSrSnTtVnTi 40 7 94 30
12 2 2 BcEyFaVnTi 50 2 24 50
BcEyFaPnVnTi 50 23 50
13 7 4 BcCpEyFaKnNtPnTtVnTi 57.1 2 103 85.7
14 5 1 BcEyFaVnTi 100 3 32 40
34 40
15 5 4 EyFaKnPnSrSnVnTi 40 4 27 40
16 5 4 BcCpEyFaKnNtPnSrSnTtVnTi 40 4 85 40
17 6 2 BcCpEyFaKnNtPnSrSnTtVnTi 66.7 5 115 33.3
18 3 2 BcCpEyFaKnNtPnSrSnTtVnTi 66.7 2 14 66.7
19 5 4 BcCpEyFaKnPnSrSnVnTi 40 4 112 40
20b(18/02/02) 9 5 BcEyFaKnNtPnTtVnTi 33.3 1 38 100
BcCpEyFaNtPnTtVnTi 33.3
20b(02/09/02) 3 2 BcCpEyFaKnNtPnSrTtVnTi 66.7 3 65 66.7
21 8 6 BcCpEyFaKnNtPnSrTtVnTi 37.5 6 96 37.5
23b 6 3 BcEyFaKnNtPnSrTtVnTi 66.7 1 35 100
24c 5 2 EyFaPnSnTtVnTi 60 1 3 100
25 7 6 BcEyFaNtPnSnTtVnTi 28.6 2 41 71.4
26 4 4 None 2 18
17
50 50
27 4 3 BcEyFaKnNtPnSrSnTtVnTi 50 3 12 50
28 9 4 BcEyFaKnNtPnSrSnTtVnTi 44.4 7 16 22.2
84 22.2
29 6 5 BcCpFaKnNtPnSrSnTtVnTi 33.3 4 53
55
33.3 33.3
30b 2 2 EyFaNtVnTi 50 2 71 50
BcEyFaKnPnSrTtVnTi 50 37 50
a
Bc, bacitracin; Cp, ciprofloxacin; Ey, erythromycin; Fa, flavomycin; Kn, kanamycin; Nt, nitrofurantoin; Pn, penicillin; Sr, streptomycin; Sn, synercid; Tr, tetracycline; Vn, vancomycin; Ti, teicoplanin.
b
Organic farm.
c
Pig farm.
FIG. 1. Dendrogram generated by Gel Compar II software showing the relationships of 119 representative fingerprints (SmaI-PFGE types) for 217 VREF isolates. The analysis of the generated bands was performed by using the Dice coefficient and the unweighted-pair group method with arithmetic averages (optimization of 0.00% and position tolerance of 1.00%).
on May 15, 2020 by guest
http://jcm.asm.org/
[image:5.585.44.540.81.442.2]though PFGE is still considered the standard typing method for enterococci, there are no standardized criteria for analyz-ing PFGE patterns (16). Therefore, the interpretation of the results can lead to different conclusions. In addition, the lack of standardization in terms of PFGE running conditions makes interlaboratory comparisons rather limited. The issue of how many band differences account for the description of a new clone has not yet been resolved. This is especially true for long-term studies. In the present study, there was only one case of related PFGE types being isolated on two different premises that were spatially separated and managed by different farm-ers. Farm 8 was a conventional poultry farm which supplied stock to farm 3 (organic poultry farm). Interestingly, similar PFGE profiles were found on each of these farms, differing in only one band. The link between farms was curtailed during the course of the study, and subsequent visits to farm 3 did not recover further VREF isolates. This event might suggest a transfer of resistant strains from farm to farm.
PFGE has provided evidence of the high level of genetic diversity amongE. faeciumpopulations in farm environments in the United Kingdom. The PFGE results of our study suggest that the introduction of new clones to the farm by deliveries of new stock or by other reservoirs (e.g., other domestic and wild animals, feed, litter, and water), rather than the persistence of resistant clones, may be the cause for the persistence of van-comycin resistance on these farms. Alternatively, we may have observed the effects of the dynamic interaction of bacterial populations, by which the previously detected clone called “the dominant type” may still be present on the farm on subsequent visits, but at levels below the limit of detection, whereas “new” clones that previously survived in small numbers may increase to detectable levels for unknown reasons. Management prac-tices such as the use of disinfectants, sources of replacement stock, or even interactions with other enteric flora might have an important impact on the selection of the new dominant bacterial population.
VREF isolates from different habitats are very polyclonal, suggesting the horizontal gene transfer of the vancomycin re-sistance genes rather than the spread of a single clone. There-fore, if we cannot find the same clones in different environ-ments, but we are able to find the same resistance genes, some clear questions arise. Where is the transfer ofvanand other genes occurring? Are farm animals a significant long-term res-ervoir of these resistance genes, and if so, since avoparcin has not been administered to livestock since 1999, are we coselect-ing for vancomycin resistance by the use of other compounds? There may also be other undetermined factors indirectly se-lecting for the persistence of the vancomycin resistance genes if these organisms are more suited for environmental survival. Theespgene encodes an enterococcal surface protein (Esp), which contributes to the colonization and infection of the uri-nary tract by increasing attachment to epithelial surfaces and biofilm production (22). This gene appears to be an entero-coccus-specific virulence factor which is highly conserved inE. faecium subpopulations involved in hospital outbreaks (31), independent of the vancomycin susceptibility (23). The isolates tested in our study were negative for this element, which sug-gests that farm animals may not be a significant source of these genes. The absence ofespgenes in our isolates indicates that it is unlikely that they could cause infections in humans, although
“in vitro” conjugative transfer of theespgene has been dem-onstrated (21).
In our study, we detected antimicrobial resistance even on farms where antimicrobials had not been used for many years, if at all. The factors promoting the persistence of resistant bacteria or resistance genes are not clear. Ideally, farm anti-microbial usage and hospital policies should be implemented to minimize the further development, spread, and persistence of resistant organisms. Statistically representative surveys should be carried out to detect and quantify specific genes in agricultural systems as well as in hospital environments. In addition, epidemiological studies to help to unravel the mech-anisms underlying the observed heterogeneity of VREF iso-lates should be attempted. This would provide useful informa-tion to help to prevent and ultimately control the spread of antimicrobial resistance among bacteria.
ACKNOWLEDGMENTS
We gratefully acknowledge Defra for funding project OD2006. Lourdes Garcia-Migura is a Ph.D. student registered with the Univer-sity of Liverpool.
We also thank Felicity Clifton-Hadley for her corrections and sug-gestions and the participating farmers for letting us undertake this research on their farms.
REFERENCES
1.Aarestrup, F. M., H. Hasman, L. B. Jensen, M. Moreno, I. A. Herrero, L. Dominguez, M. Finn, and A. Franklin.2002. Antimicrobial resistance among enterococci from pigs in three European countries. Appl. Environ. Micro-biol.68:4127–4129.
2.Aarestrup, F. M., H. Kruse, E. Tast, A. M. Hammerum, and L. B. Jensen. 2000. Associations between the use of antimicrobial agents for growth pro-motion and the occurrence of resistance among Enterococcus faecium from broilers and pigs in Denmark, Finland, and Norway. Microb. Drug Resist. 6:63–70.
3.Aarestrup, F. M., A. M. Seyfarth, H. D. Emborg, K. Pedersen, R. S. Hen-driksen, and F. Bager.2001. Effect of abolishment of the use of antimicrobial agents for growth promotion on occurrence of antimicrobial resistance in fecal enterococci from food animals in Denmark. Antimicrob. Agents Che-mother.45:2054–2059.
4.Bager, F., M. Madsen, J. Christensen, and F. M. Aarestrup.1997. Avoparcin used as a growth promoter is associated with the occurrence of vancomycin-resistant Enterococcus faecium on Danish poultry and pig farms. Prev. Vet. Med.31:95–112.
5.Bates, J.1997. Epidemiology of vancomycin-resistant enterococci in the community and the relevance of farm animals to human infection. J. Hosp. Infect.37:89–101.
6.Bates, J., J. Z. Jordens, and D. T. Griffiths.1994. Farm animals as a putative reservoir for vancomycin-resistant enterococcal infection in man. J. Antimi-crob. Chemother.34:507–514.
7.Berchieri, A.1999. Intestinal colonization of a human subject by vancomycin-resistant Enterococcus faecium. Clin. Microbiol. Infect.5:97–100. 8.Bonten, M. J., R. Willems, and R. A. Weinstein.2001. Vancomycin-resistant
enterococci: why are they here, and where do they come from? Lancet Infect. Dis.1:314–325.
9.Borgen, K., Y. Wasteson, H. Kruse, and R. J. Willems.2002. Vancomycin-resistantEnterococcus faecium(VREF) from Norwegian poultry cluster with VREF from poultry from the United Kingdom and The Netherlands in an amplified fragment length polymorphism genogroup. Appl. Environ. Micro-biol.68:3133–3137.
9a.Centers for Disease Control and Prevention.Standardized protocol for mo-lecular subtyping ofListeria monocytogenesby PFGE. CDC, Atlanta, Ga. [Online.] http://www.cdc.gov/pulsenet/.
10.Coque, T. M., J. F. Tomayko, S. C. Ricke, P. C. Okhyusen, and B. E. Murray. 1996. Vancomycin-resistant enterococci from nosocomial, community, and animal sources in the United States. Antimicrob. Agents Chemother.40: 2605–2609.
11.Dutka-Malen, S., S. Evers, and P. Courvalin.1995. Detection of glycopep-tide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol.33:24–27.
12.Giraffa, G.2002. Enterococci from foods. FEMS Microbiol. Rev.26:163– 171.
13.Huycke, M. M., D. F. Sahm, and M. S. Gilmore.1998. Multiple-drug
on May 15, 2020 by guest
http://jcm.asm.org/
tant enterococci: the nature of the problem and an agenda for the future. Emerg. Infect. Dis.4:239–249.
14.Iversen, A., I. Kuhn, A. Franklin, and R. Mollby.2002. High prevalence of vancomycin-resistant enterococci in Swedish sewage. Appl. Environ. Micro-biol.68:2838–2842.
15.Martone, W. J.1998. Spread of vancomycin-resistant enterococci: why did it happen in the United States? Infect. Control Hosp. Epidemiol.19:539–545. 16.Morrison, D., N. Woodford, S. P. Barrett, P. Sisson, and B. D. Cookson. 1999. DNA banding pattern polymorphism in vancomycin-resistant Entero-coccus faeciumand criteria for defining strains. J. Clin. Microbiol.37:1084– 1091.
17.Moubareck, C., N. Bourgeois, P. Courvalin, and F. Doucet-Populaire.2003. Multiple antibiotic resistance gene transfer from animal to human entero-cocci in the digestive tracts of gnotobiotic mice. Antimicrob. Agents Che-mother.47:2993–2996.
18.Murray, B. E.1990. The life and times of theEnterococcus. Clin. Microbiol. Rev.3:46–65.
19.Murray, B. E. 1998. Diversity among multidrug-resistant enterococci. Emerg. Infect. Dis.4:37–47.
20.Noble, W. C., Z. Virani, and R. G. Cree.1992. Co-transfer of vancomycin and other resistance genes from Enterococcus faecalis NCTC 12201 to Staphy-lococcus aureus. FEMS Microbiol. Lett.72:195–198.
21.Oancea, C., I. Klare, W. Witte, and G. Werner.2004. Conjugative transfer of the virulence gene,esp, among isolates of Enterococcus faecium and En-terococcus faecalis. J. Antimicrob. Chemother.54:232–235.
22.Shankar, N., C. V. Lockatell, A. S. Baghdayan, C. Drachenberg, M. S. Gilmore, and D. E. Johnson.2001. Role ofEnterococcus faecalissurface protein Esp in the pathogenesis of ascending urinary tract infection. Infect. Immun.69:4366–4372.
23.Shankar, V., A. S. Baghdayan, M. M. Huycke, G. Lindahl, and M. S. Gil-more.1999. Infection-derivedEnterococcus faecalisstrains are enriched in
esp, a gene encoding a novel surface protein. Infect. Immun.67:193–200. 24.Sorensen, T. L., M. Blom, D. L. Monnet, N. Frimodt-Moller, R. L. Poulsen,
and F. Espersen.2001. Transient intestinal carriage after ingestion of anti-biotic-resistant Enterococcus faecium from chicken and pork. N. Engl. J. Med.345:1161–1166.
25.Stobberingh, E., A. van den Bogaard, N. London, C. Driessen, J. Top, and R. Willems.1999. Enterococci with glycopeptide resistance in turkeys, turkey farmers, turkey slaughterers, and (sub)urban residents in the south of The Netherlands: evidence for transmission of vancomycin resistance from ani-mals to humans? Antimicrob. Agents Chemother.43:2215–2221. 26.Tenover, F. C., L. M. Weigel, P. C. Appelbaum, L. K. McDougal, J.
Chait-ram, S. McAllister, N. Clark, G. Killgore, C. M. O’Hara, L. Jevitt, J. B. Patel, and B. Bozdogan.2004. Vancomycin-resistantStaphylococcus aureusisolate from a patient in Pennsylvania. Antimicrob. Agents Chemother.48:275–280. 27.Thal, L. A., and M. J. Zervos.1999. Occurrence and epidemiology of resis-tance to virginiamycin and streptogramins. J. Antimicrob. Chemother.43: 171–176.
28.Turabelidze, D., M. Kotetishvili, A. Kreger, J. G. Morris, Jr., and A. Su-lakvelidze.2000. Improved pulsed-field gel electrophoresis for typing van-comycin-resistant enterococci. J. Clin. Microbiol.38:4242–4245.
29.van Den Bogaard, A. E., R. Willems, N. London, J. Top, and E. E. Stobber-ingh.2002. Antibiotic resistance of faecal enterococci in poultry, poultry farmers and poultry slaughterers. J. Antimicrob. Chemother.49:497–505. 30.Weigel, L. M., D. B. Clewell, S. R. Gill, N. C. Clark, L. K. McDougal, S. E.
Flannagan, J. F. Kolonay, J. Shetty, G. E. Killgore, and F. C. Tenover.2003. Genetic analysis of a high-level vancomycin-resistant isolate of Staphylococ-cus aureus. Science302:1569–1571.
31.Willems, R. J., W. Homan, J. Top, M. van Santen-Verheuvel, D. Tribe, X. Manzioros, C. Gaillard, C. M. Vandenbroucke-Grauls, E. M. Mascini, E. van Kregten, J. D. van Embden, and M. J. Bonten.2001. Variant esp gene as a marker of a distinct genetic lineage of vancomycin-resistant Enterococ-cus faecium spreading in hospitals. Lancet357:853–855.
32.Witte, W.1997. Impact of antibiotic use in animal feeding on resistance of bacterial pathogens in humans. Ciba Found. Symp.207:61–71.