Original Article
microRNA-188 over-expression
inhibited cellular proliferation and
invasion of gastric cancer by targeting SOX4
Ping Zhang1, Baozhu Yu2, Zhiming Chen3
1Department of Surgical Nursing, Nursing School of Jilin University, China; 2Center for Disease Control and
Prevention of Jilin Province, China; 3Department of Forensic Medicine, College of Basic Medicine, Jilin University,
China
Received March 23, 2016; Accepted August 14, 2016; Epub September 15, 2016; Published September 30, 2016
Abstract: Previous studies reported that microRNA-188 (miR-188) was down-regulated and acted as a tumor sup-pressor in oral squamous cell carcinoma, hepatocellular carcinoma and prostate cancer. However, the expression, biological roles and molecular mechanism of miR-188 in gastric cancer remain unclear. In this study, we measured the expression levels of miR-188 in gastric cancer tissues and cell lines. The results showed that miR-188 was
significantly down-regulated in gastric cancer tissues and cell lines. Then following functional studies revealed that
miR-188 over-expression inhibited proliferation and invasion of gastric cancer cells. Using bioinformatic analysis,
luciferase report assay, qRT-PCR and western blot, SOX4 was identified as a novel direct target gene of miR-188 in
gastric cancer. Moreover, SOX4 under-expression had the similar functions with miR-188 over-expression in gastric cancer, further suggesting that miR-188 inhibited proliferation and invasion of GC cells though directly targeting
SOX4. These findings indicated that miR-188 was down-regulated and acted as a tumor suppressor in gastric can -cer through directly targeting SOX4. miR-188/SOX4 based targeted therapy could be a novel therapeutic application to treat patients with gastric cancer.
Keywords: microRNA-188, gastric cancer, proliferation, invasion, SOX4
Introduction
Gastric cancer (GC) is the fourth most common cancer and the second leading cause of cancer mortality around the world [1]. Based on the GLOBOCAN 2012 estimates, approximately 951,600 new GC cases and 723,100 deaths due to GC occurred in 2012 [2]. The highest morbidities are in Eastern Asia, Eastern Europe, and South America, especially in China whose patients with GC account for 42% of all cases in the world [3]. Environmental and behavioral habits have been demonstrated to involve in the tumorigenesis of GC, including Helicobacter pylori infec tion, high-salted diet and smoking [4]. Currently, surgery is the primary therapeu-tic strategy for the early stage of GC. However, most patients are diagnosed in the advanced stage with lymphatic, peritoneal or distal organ metastases [5]. In the last few decades, with great progress in surgery, chemotherapy, radio-therapy and molecular targeted radio-therapy, the
prognosis of GC was improved [6]. However, the long-term prognosis of GC patients remains poor, especially for advanced stage GC with a 5-year overall survival rate of 25% or less [7]. Therefore, it is urgent to investigate the molecu-lar mechanism underlying GC carcinogenesis and progression, and develop novel therapeutic targets to improve the survival rates of GC patients.
includ-ing cell proliferation, apoptosis, differentiation, carcinogenesis, metastasis, and drug resis-tance [11-13]. Increasing evidences have dem-onstrated miRNAs are abnormally expressed in various kinds of human cancers, also including GC [14-16]. The abnormal expression of
miR-NAs and their profiles in human cancers could
be applied not only in cancer prediction and
prog nosis, but also in tumor classification and
progression [17]. In the context of cancers, miRNAs could act as oncogene or tumor sup-pressor in the carcinogenesis and progression of cancers by directly targeting tumor suppres-sors or oncogenes, respectively [18, 19]. Therefore, miRNAs have considerable potential in early screening, diagnosis and treatments of human cancers.
In this study, we revealed that miR-188 expres-sion was lower in GC tissue and cell lines com-pared with corresponding non-tumor gastric tissues and human immortalized gastric epi-thelial, respectively. Restoration miR-188 expr- ession inhibited the proliferation and invasion
of GC cells. SOX4 was identified as a novel
direct target gene of miR-188 in GC.
Material and methods
Tissue samples
Fresh frozen human GC tissue samples and corresponding non-tumor gastric tissues were obtained from 46 patients with GC who
under-go radical gastrectomy at Affiliated Hospitals
of Jilin University. None of these patients had received chemotherapy, radiotherapy and mol- ecular targeted therapy prior to radical gastrec-tomy. This study was approved by Research Ethics Committee of Jilin University.
Cell culture
The human GC cell lines AGS, BGC-823, MKN-28, MKN-1, SGC-7901, and MKN-45 were pur-chased from American Type Culture Collection (ATCC, Rockville, MD, USA). The human immor-talized gastric epithelial (GES-1) and human embryonic kidney cells (HEK239T) were obt- ained from Beijing Institute for Cancer Research (Beijing, China). All these cells were grown in RPMI 1640 or DMEM culture medium (Gibco, Grand Island, NY) supplemented with 10% fetal
bovine serum (FBS; Gibco) in a humidified
atmosphere consisting with 5% CO2 and 95% air at 37°C.
Cell transfection
miR-188 mimics and miRNA mimics negative control (NC) were ordered from GenePharma (Shanghai, China). The small interfering RNA (siRNA) targeting SOX4 (siRNA-SOX4) and its negative control siRNA (siRNA-NC) were pur-chased from SunBio (SunBio Medical Biote- chnology, Guangzhou, China). For transfection, cells were seeded into 6-well plates at a density
of 70%~80% confluence. After incubation over -night, cells were transfected with mimics or siRNA at a concentration of 50 nM using Lipofectamine 2000 reagent (Invitrogen, Car- lsbad, CA, USA). Six hours after transfection, cells were continuously incubated with com-plete culture medium.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted using TRIzol agent (Invitrogen), according to the manufacturer’s instructions. For miR-188 expression, cDNA was synthesized from RNA using Taqman microRNA reverse transcription kit (Applied Biosystems, Foster City, CA, USA). Taqman microRNA assay kit (Applied Biosystems) was used to detect miR-188 and U6 expression. Quantitative (q)PCR was performed in triplicate on a 7900HT Real-Time PCR System (Applied Biosystems), and U6 was measured as an endogenous control for miR-188. For the detec-tion of SOX4 mRNA expression, a SYBR Green Real-Time PCR Master Mix Kit (Applied Bio- systems) was applied with GAPDH as an inter-nal control. The relative expression of miR-188 and SOX4 mRNA were analyzed by use of the 2-ΔΔCt method.
Cell proliferation assay
Cell proliferation assay was performed to deter-mine cell proliferation rate with Cell Counting Kit 8 (CCK8; Dojindo, Kumamoto, Japan) assay
kit. Briefly, GC cells were seeded into 96-well
plates at a density of 3000 cells per well. Following transfection, cells were incubated for
siRNA) using RIPA lysis buffer supplemented with proteinase/phosphatase inhibitors (The-
rmo Fisher Scientific, Cambridge, MA, USA).
Following 10% SDS polyacrylamide gel electro-phoresis, the separated proteins were
trans-ferred onto a polyvinylidene difluoride mem -branes (Millipore, Bedford, MA, USA). The mem-branes were then blocked in 5% nonfat dried milk in phosphate-buffered saline (PBS)-Tween for 3 h and incubated with primary antibodies, mouse anti-human monoclonal antibody SOX4 (1:10,00 dilution; sc-130633, Santa Cruz Bio-
technology, CA, USA) and β-actin (1:10,00 dilu -tion; sc-47778, Santa Cruz Biotechnology). After incubation at 4°C overnight, the mem-branes were washed three times and probed with goat anti-mouse horseradish peroxidase-conjugated mouse second antibody (1:10,000; Santa Cruz Biotechnology) for 1 h at room tem-perature. Membranes were washed again for three times and the protein signals were visual-ized using an enhanced chemiluminescence
solution (Pierce, Rockford, IL, USA). β-actin was
used as an endogenous control for SOX4 pro-tein expression.
Luciferase reporter assay
PmirGLO-SOX4-3’UTR wild-type (WT) and mir-GLO-SOX4-3’UTR mutant (MUT) luciferase re- porter vectors were obtained from GenePh- arma. For luciferase reporter assay, HEK293T cells were co-transfected with PmirGLO-SOX4-3’UTR WT or PmirGLO-SOX4-PmirGLO-SOX4-3’UTR MUT, and miR-188 mimics or NC. At 48 h after transfec-tion, cells were collected and the relative lucif-erase activities were determined using Dual-Luciferase Reporter Assay System (Promega, Manheim, Germany) according to the manufac-turer’s instructions. Experiments were per-formed in triplicate and replicated 3 times.
Statistical analysis
Data were presented as mean ± S.D. Statistical analyses were performed with student’s t-test by using SPSS 15.0 (SPSS Inc.; Chicago, IL, USA). Double-tailed P value less than 0.05 was
considered to be statistically significant.
Results
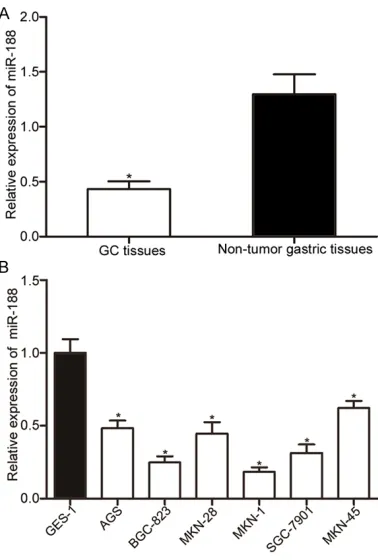
miR-188 expression was down-regulated in GC
We first examined miR-188 expression in GC
tissues and corresponding non-tumor gastric
Cell invasion assay
Cell invasion assay was performed to deter-mine cell invasion abilities by using Matrigel (BD Biosciences) coated transwell chambers (8-mm pores; BD Biosciences, San Jose, CA, USA). At 48 h after transfection, 4×104 trans-fected cells (mimics/siRNA) were seeded into the upper chambers in FBS-free culture media.
The lower chambers were filled with complete
culture medium supplemented with 20% FBS.
After incubation 48 h in a humidified atmo -sphere consisting with 5% CO2 and 95% air at 37°C, cells on the upper surface of the tran-swell insert chambers were removed using a cotton-tipped swab. The invaded cells were
fixed, stained with 0.5% crystal violet and then
visualized under an inverted microscope. Five
visual fields of each chamber were randomly
chosen and each experiment was repeated at least three times.
Western blot analysis
[image:3.612.93.282.70.350.2]significantly down-regulated in GC
tissues compared with that in cor-responding non-tumor gastric tis-sues (P<0.05). We then deter-mined miR-188 expression levels in GC cell lines (AGS, BGC-823, 28, 1, SGC-7901, MKN-45) and human immortalized gas-tric epithelial (GES-1). As shown in Figure 1B, all GC cell lines expressed lower miR-188 level than GES-1 (P<0.05). Therefore, we concluded that down-regulation of miR-188 might play important roles in GC.
miR-188 acted as a tumor sup-pressor in GC cells
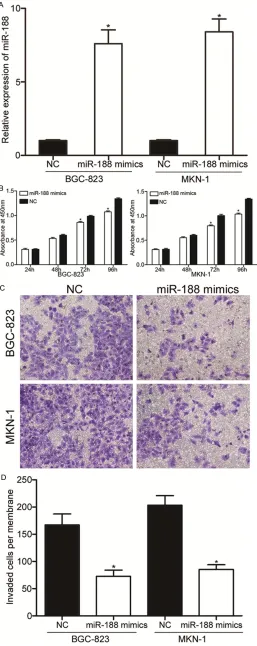
As miR-188 was expressed at lower levels in GC tissues and cell lines, we hypothesized that miR-188 might act as a tumor suppres-sor in GC. Therefore, miR-188 mim-ics or NC was transfected into BGC-823 and MKN-1 cells which showed lower miR-188 expression. As shown in Figure 2A, miR-188 mimics could markedly increa- sed miR-188 expression levels in both BGC-823 and MKN-1 cells (P<0.05).
[image:4.612.95.352.73.719.2]To investigate the effect of miR-188 on proliferation of GC, cell pro-liferation assay was performed. As shown in Figure 2B, the prolifera-tion of BGC-823 and MKN-1 cells transfected with miR-188 mimics was reduced compared with that of BGC-823 and MKN-1 cells trans-fected with NC (P<0.05).
Figure 2. miR-188 over-expression in-hibited proliferation and invasion of GC cells. A. qRT-PCR analysis of relative miR188 expression in BGC-823 and MKN-1 cells transfected with miR-188 mimics or NC. B. Cell proliferation assay was used to determine the cellular pro-liferation in BGC-823 and MKN-1 cells following transfection with miR-188 mimics or NC. C, D. The invaded cells per membrane in the miR-188-mimic
groups were significantly less than that
significantly lower in the miR-188
groups compared with NC groups
(P<0.05). All these findings suggest -ed that miR-188 act-ed as a tumor suppressor in GC.
SOX4 was a direct target of miR-188 in GC.
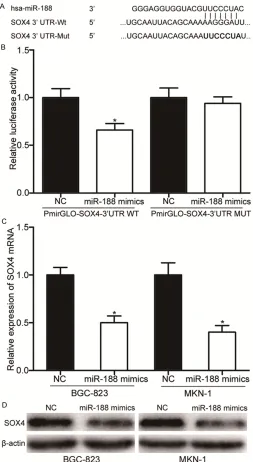
Due to the important functions of miR-188 in GC, we explored the underlying mechanisms. Using the publicly available databases miRan-da (www.microrna.org) and TargetS-can (www.targetsTargetS-can.org), we found that SOX4 was a putative target gene of miR-188 (shown in Figure 3A).
To investigate whether miR-188 directly recognizes the 3’UTR of SOX4, PmirGLO-SOX4-3’UTR WT or PmirGLO-SOX4-3’UTR MUT was tra- nsfected into HEK293T cells in com-bination with miR-188 mimics or NC. The results showed that miR-188 mimics decreased the luciferase activities of PmirGLO-SOX4-3’UTR WT compared with NC groups, while the luciferase activities of PmirGLO-SOX4-3’UTR MUT was restored (sh- own in Figure 3B). It indicated that
miR-188 could specifically bind to
the 3’UTR of SOX4 and decrease its luciferase activities, while MUT vec-tor could not be combined with miR-188 to decrease its luciferase activi-ties (P<0.05).
We then used qRT-PCR and western blot to detect SOX4 expression lev-els in BGC-823 and MKN-1 cells transfected with miR-188 mimics or NC. As shown in Figure 3C and 3D,
miR-188 over-expression
significa-ntly decreased SOX4 expression at both the mRNA and protein levels in BGC-823 and MKN-1 cells (P<0.05). Taken together, SOX4 was a direct target of miR-188 in GC.
[image:5.612.93.346.71.533.2]We also performed cell invasion assay to evalu-ate the function of miR-188 in regulating the invasion ability of GC cells. As shown in Figure 2C and 2D, the average number of invaded cells penetrating the transwell membrane was
Figure 3. SOX4 was a novel target of miR-188. A. Predicted conse-quential pairing of target 3’ UTR region of SOX4 (WT and MUT) and miR-188 sequence. B. HEK293T cells were co-transfected with PmirGLO-SOX4-3’UTR WT or PmirGLO-SOX4-3’UTR MUT, and either miR-188 mimics or NC. Luciferase activities was normalized to the activities of Renilla luciferase. C. qRT-PCR analysis for SOX4 mRNA of BGC-823 and MKN-1 cells transfected with miR-188 mimics or NC. D. Western blot analysis for SOX4 protein of BGC-823 and MKN-1 cells transfected with miR-188 mimics or NC. *P<0.05.
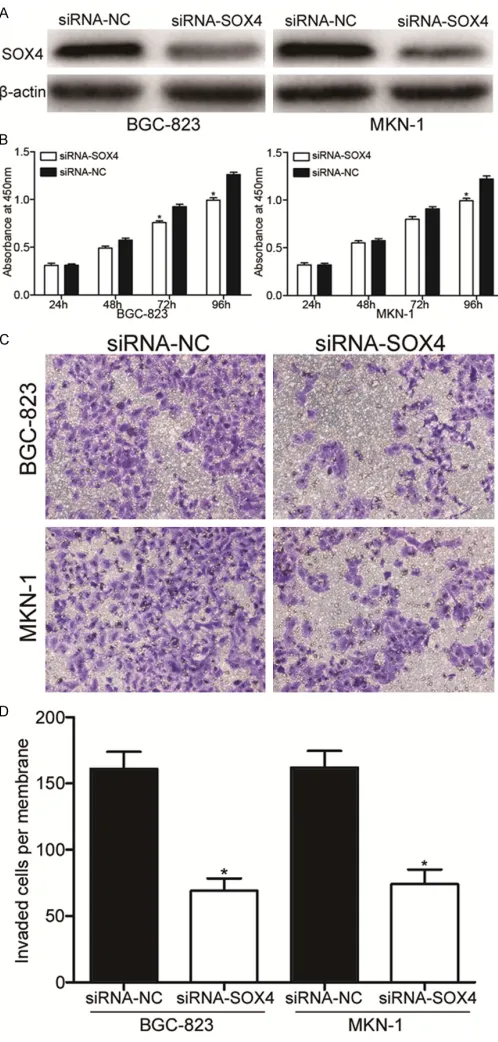
Effects of SOX4 knockdown on the prolifera-tion and invasion of GC cells
Figure 4. SOX4 under-expression inhibited proliferation and invasion of GC cells. A. Western blot analysis for SOX4 protein of BGC-823 and MKN-1 cells transfected with siRNA-SOX4 or siRNA-NC. B. Cell prolif-eration assay was used to determine the cellular prolifprolif-eration in BGC-823 and MKN-1 cells following transfection with SOX4 or siRNA-NC. C, D. The invaded cells per membrane in the siRNA-SOX4 groups
were significantly less than that of the siRNA-NC groups. *P<0.05.
BGC-823 and MKN-1 cells. As shown in Figure 4A, SOX4 protein levels in siRNA-SOX4
trans-fection BGC-823 and MKN-1 cells
were significantly lower compared
to that in siRNA-NC-transfected BGC-823 and MKN-1cells. After siR-NA-SOX4 had been transfected into GC cells, we investigated the effects of SOX4 on proliferation and inva-sion of GC cells. As shown in Figure 4B, cell proliferation was significant -ly reduced in BGC-823 and MKN-1 cells transfected with siRNA-SOX4 compared with that in those trans-fected with siRNA-NC (P<0.05). As shown in Figure 4C and 4D, the average number of invaded cells penetrating the transwell
mem-brane was significantly lower in the
siRNA-SOX4 groups compared with siRNA-NC groups (P<0.05). These results may partly elucidated the miR-188 inhibited proliferation and invasion of GC cells though down-regulation of SOX4.
Discussion
The tumorigeneis and tumor devel-opment of GC is controlled by multi-ple factors, and a great deal of diverse molecules act as important factors in GC [20]. Previous studies have indicated that miRNAs are important in numerous aspects of human cancers and play important roles in carcinogenesis and cancer development [21-23]. miRNAs have been reported to act as tumor sup-pressors or oncogenes in human cancer, providing a new perspective on the treatment of cancer [18, 19]. However, possible roles of miRNAs in carcinogenesis and progression of GC remain largely unexplored. In this study, we found that miR-188
was significantly down-regulated in
GC tissues and cell lines (AGS, BGC-823, MKN-28, MKN-1, SGC-7901, MKN-45) compared with in corre-sponding non-tumor gastric tissues and GES-1 which is a human immor-talized gastric epithelial, respective-ly. In addition, restoration of miR-188 expression inhibited prolifera-tion and invasion of GC cells. Furthermore,
miR-188 in GC. SOX4 under-expression also inhibit-ed proliferation and invasion of GC cells. These results suggested that miR-188 might act as a tumor suppressor in GC through directly target-ing SOX4.
miR-188 has been mainly studied in several human cancers, including oral squamous cell carcinoma [24], heaptocellular carcinoma [25], prostate cancer [26] and acute myeloid leuke-mia [27]. Recent study demonstrated that miR-188 was down-regulated in oral squamous cell carcinoma, and miR-188 overexpression inhib-ited cell proliferation, cell cycle progression and invasion [24]. miR-188 was also down-regulat-ed in hepatocellular carcinoma, and its expres-sion levels were highly correlated with multiple nodules, microvascular invasion, overall and disease-free survival. Functionally, miR-188 suppressed proliferation and metastasis of hepatocellular carcinoma cells in vitro and in vivo [25]. Zhang et. al reported that miR-188
was significantly down-regulated in prostate
cancer with metastasis. Low expression levels of miR-188 were an independent prognostic factor for poor overall and biochemical recur-rence-free survival. Ectopic of miR-188 expres-sion inhibited prostate cancer cells prolifera-tion, migration and invasion in vitro and sup-pressed tumour growth and metastasis in vivo [26]. These studies suggested that miR-188 may be important in these types of cancer and may serve as a potential therapeutic target for their treatments.
Since miRNAs function through targeting differ-ent mRNAs, we deduced that miR-188 might directly target mRNAs to inhibit proliferation and invasion of GC cells. Studies revealed that it may regulate oncogenic transcripts in human cells, including SIX1 [24], FGF5 [25], and LAPTM4B [26]. In this study, SOX4 was validat-ed to be a novel direct target gene of miR-188 in GC. Firstly, two publicly available databases (miRanda and TargetScan) predicated that SOX4 was a putative target of miR-188. Sec- ondly, luciferase report assays demonstrated
that miR-188 could specifically bind to the
3’UTR of SOX4. Thirdly, qRT-PCR and western blot revealed that miR-188 decreased SOX4 expression at both mRNA and protein levels in GC cells. Finally, SOX4 under-expression has the similar effects with miR-188
over-expres-sion in GC. Identification of miR-188 target
genes is critical for understanding its role in tumorigenesis and progression of GC.
The sex-determining region Y (SRY) box family, also referred to as the SOX family, comprises 20 highly conserved transcription factors [28]. SOX4, a member of the SOX family, is a 47-kDa protein that is encoded by a single exon gene [29]. Increasing evidences suggested that
SOX4 was significantly up-regulated in many
kinds of human cancers, such as breast cancer [30], prostate cancer [31], lung cancer [32], hepatocellular carcinoma [33] and GC [28]. In GC, expression levels of SOX4 were correlated with depth of invasion, nodal status, distant metastasis, stage, and vascular invasion. Patients with high expression levels of SOX4
achieved a significantly poorer disease-free
survival rate, compared with patients with low SOX4 expression levels. Univariate Cox regres-sion analysis showed that SOX4 over-expres-sion was a clear prognostic marker for GC [28]. These results indicated that SOX4 was a
suit-able target for GC. Combination with findings in
the present study, miR-188/SOX4 based tar-geted therapy may be a novel therapeutic strat-egy for patients with GC.
In conclusion, we have four major findings in this study: 1) miR-188 was significantly
down-regulated in GC tissues and cell lines. 2) Ectopic expression of miR-188 inhibited proliferation and invasion of GC cells. 3) miR-188 could directly target SOX4, which was a novel target gene of miR-188. 4) SOX4 under-expression had the similar functions with miR-188 over-expression in GC.
Disclosure of conflict of interest
None.
Address correspondence to: Drs. Ping Zhang and Zhiming Chen, Department of Surgical Nursing, Nursing School of Jilin University, 965 Xinjiang Street, Changchun 130021, Jilin Province, China. E-mail: [email protected] (PZ); czmzhiming@ hotmail.com (ZMC)
References
[1] Jemal A, Bray F, Center MM, Ferlay J, Ward E and Forman D. Global cancer statistics. CA Cancer J Clin 2011; 61: 69-90.
[2] Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieu-lent J and Jemal A. Global cancer statistics, 2012. CA Cancer J Clin 2015; 65: 87-108. [3] Leung WK, Wu MS, Kakugawa Y, Kim JJ, Yeoh
KG, Goh KL, Wu KC, Wu DC, Sollano J, Kachin-torn U, Gotoda T, Lin JT, You WC, Ng EK, Sung
Screening for gastric cancer in Asia: current evidence and practice. Lancet Oncol 2008; 9: 279-287.
[4] Kaneko S and Yoshimura T. Time trend analy-sis of gastric cancer incidence in Japan by his-tological types, 1975-1989. Br J Cancer 2001; 84: 400-405.
[5] Yuasa Y. Control of gut differentiation and in-testinal-type gastric carcinogenesis. Nat Rev Cancer 2003; 3: 592-600.
[6] Lee JH, Kim KM, Cheong JH and Noh SH. Cur-rent management and future strategies of gas-tric cancer. Yonsei Med J 2012; 53: 248-257. [7] Hartgrink HH, Jansen EP, van Grieken NC and
van de Velde CJ. Gastric cancer. Lancet 2009; 374: 477-490.
[8] Ma L, Teruya-Feldstein J and Weinberg RA. Tu-mour invasion and metastasis initiated by mi-croRNA-10b in breast cancer. Nature 2007; 449: 682-688.
[9] Wang Z, Wang J, Yang Y, Hao B, Wang R, Li Y and Wu Q. Loss of has-miR-337-3p expression is associated with lymph node metastasis of human gastric cancer. J Exp Clin Cancer Res 2013; 32: 76.
[10] Carrington JC and Ambros V. Role of microR-NAs in plant and animal development. Science 2003; 301: 336-338.
[11] Phuah NH and Nagoor NH. Regulation of mi-croRNAs by natural agents: new strategies in cancer therapies. Biomed Res Int 2014; 2014: 804510.
[12] Ling H, Fabbri M and Calin GA. MicroRNAs and other non-coding RNAs as targets for antican-cer drug development. Nat Rev Drug Discov 2013; 12: 847-865.
[13] Ambros V. The functions of animal microRNAs. Nature 2004; 431: 350-355.
[14] Wu XL, Cheng B, Li PY, Huang HJ, Zhao Q, Dan ZL, Tian DA and Zhang P. MicroRNA-143 sup-presses gastric cancer cell growth and induces apoptosis by targeting COX-2. World J Gastro-enterol 2013; 19: 7758-7765.
[15] Naito Y, Yasuno K, Tagawa H, Sakamoto N, Oue N, Yashiro M, Sentani K, Goto K, Shinmei S, Oo HZ, Yanagihara K, Hirakawa K and Yasui W. Mi-croRNA-145 is a potential prognostic factor of scirrhous type gastric cancer. Oncol Rep 2014; 32: 1720-1726.
[16] Zhang XL, Shi HJ, Wang JP, Tang HS, Wu YB, Fang ZY, Cui SZ and Wang LT. MicroRNA-218 is upregulated in gastric cancer after cytoreduc-tive surgery and hyperthermic intraperitoneal chemotherapy and increases chemosensitivity to cisplatin. World J Gastroenterol 2014; 20: 11347-11355.
[17] Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell 2009; 136: 215-233.
[18] Shenouda SK and Alahari SK. MicroRNA func-tion in cancer: oncogene or a tumor suppres-sor? Cancer Metastasis Rev 2009; 28: 369-378.
[19] Chen CZ. MicroRNAs as oncogenes and tumor suppressors. N Engl J Med 2005; 353: 1768-1771.
[20] Zhang SJ, Feng JF, Wang L, Guo W, Du YW, Ming L and Zhao GQ. miR-1303 targets clau-d18 gene to modulate proliferation and in-vasion of gastric cancer cells. Dig Dis Sci 2014; 59: 1754-1763.
[21] Croce CM and Calin GA. miRNAs, cancer, and stem cell division. Cell 2005; 122: 6-7. [22] Tao J, Wu D, Xu B, Qian W, Li P, Lu Q, Yin C and
Zhang W. microRNA-133 inhibits cell prolifera-tion, migration and invasion in prostate cancer cells by targeting the epidermal growth factor receptor. Oncol Rep 2012; 27: 1967-1975. [23] Gao P, Xing AY, Zhou GY, Zhang TG, Zhang JP,
Gao C, Li H and Shi DB. The molecular mecha-nism of microRNA-145 to suppress invasion-metastasis cascade in gastric cancer. Onco-gene 2013; 32: 491-501.
[24] Wang L and Liu H. microRNA-188 is downregu-lated in oral squamous cell carcinoma and in-hibits proliferation and invasion by targeting SIX1. Tumour Biol 2015; 37: 4105-13.
[25] Fang F, Chang RM, Yu L, Lei X, Xiao S, Yang H and Yang LY. MicroRNA-188-5p suppresses tu-mor cell proliferation and metastasis by direct-ly targeting FGF5 in hepatocellular carcinoma. J Hepatol 2015; 63: 874-885.
[26] Zhang H, Qi S, Zhang T, Wang A, Liu R, Guo J, Wang Y and Xu Y. miR-188-5p inhibits tumour growth and metastasis in prostate cancer by repressing LAPTM4B expression. Oncotarget 2015; 6: 6092-6104.
[27] Jinlong S, Lin F, Yonghui L, Li Y and Weidong W.
Identification of let-7a-2-3p or/and
miR-188-5p as prognostic biomarkers in cytogenetically normal acute myeloid leukemia. PLoS One 2015; 10: e0118099.
[28] Fang CL, Hseu YC, Lin YF, Hung ST, Tai C, Uen YH and Lin KY. Clinical and prognostic associa-tion of transcripassocia-tion factor SOX4 in gastric can-cer. PLoS One 2012; 7: e52804.
[29] Li Y, Zu L, Wang Y, Wang M, Chen P and Zhou Q. miR-132 inhibits lung cancer cell migration and invasion by targeting SOX4. J Thorac Dis 2015; 7: 1563-1569.
[30] Zhang J, Liang Q, Lei Y, Yao M, Li L, Gao X, Feng J, Zhang Y, Gao H, Liu DX, Lu J and Huang B. SOX4 induces epithelial-mesenchymal transi-tion and contributes to breast cancer progres-sion. Cancer Res 2012; 72: 4597-4608. [31] Wang L, Li Y, Yang X, Yuan H, Li X, Qi M, Chang
epithelial-mesenchymal transition in prostate cancer cells. Prostate 2014; 74: 647-658.
[32] Castillo SD, Matheu A, Mariani N, Carretero J, Lopez-Rios F, Lovell-Badge R and Sanchez-Cespedes M. Novel transcriptional targets of the SRY-HMG box transcription factor SOX4 link its expression to the development of small cell lung cancer. Cancer Res 2012; 72: 176-186.
[33] Zheng JH, Jian ZX, Jin HS, Chen SC and Wang GY. [Expression of SOX4 gene and early recur-rence of hepatocellular carcinoma: their
rela-tionship and the clinical significance]. Nan