Before computers, animation was produced by drawing successive frames by hand or by taking snapshots of real models, which were then repositioned and their images again recorded. Modern computer animation may be divided accordingly into two-dimensional and three-dimensional techniques. The former is reminiscent of drawing flat frames on cell layers, whereas in three-dimensional animation virtual models are generated and manipulated, and the final images are produced by various rendering methods. For example, scan-line algorithms scan across the raster scan-lines and at each point check the polygons of the model, displaying the colour of the closest one or the background colour if no objects are present at that point. In ray tracing, virtual rays of light are traced through the scene, allowing true optical effects to occur.
Animation has come a long way, and not only because of the use of computers. ‘Computer animation is not done by the computer any more than clay animation is created by the clay.’ Craig Good, at Pixar Animation Studios, had in mind the talent and skills of the animators. Indeed, modern animation involves many fields of science: physics, robotics, biology, etc. In the present paper, I review work in which articulated or soft-bodied virtual models power themselves through their environment according to the laws of physics and are capable of evolving and behaving according to a mind of their own.
The degree of physical realism achieved depends on the degree to which the system is modelled. For example, visual simulations may occasionally show non-realistic movement, which is due to simplifications of the relevant biomechanics. Human perception is very accurate in finding discrepancies in details of movement and can be utilised in sorting out
oversimplifications. Simulations run on fast computers take place nearly or fully in real time, and the parameters can be changed interactively during the animation. The techniques of interactive animated simulation certainly have unused potential as research tools for several fields of biology.
Legged locomotion
The first section reviews the work on legged locomotion by Marc Raibert and Jessica Hodgins and their groups at the Massachusetts Institute of Technology and the Georgia Institute of Technology. Plenty of animations are available at their web sites (MIT Leg Laboratory, www.ai.mit.edu/projects/leglab/simulations/simulations.html; Georgia Tech Animation Laboratory, www.cc.gatech.edu/gvu/ animation/index.html). Raibert and Hodgins (1991) describe two robot-like creatures that trot, gallop, change gait and turn. These are modelled after real robots previously built by the MIT Leg Laboratory. The paper also presents a hopping kangaroo, modelled after Alexander and Vernon (1975). Hodgins and Wooten (1998) describe algorithms that allow a rigid-bodied model of a man or woman to stand, run and turn at a variety of speeds, to ride a bicycle on hills and around obstacles, and to perform gymnastic vaults. Hodgins and Pollard (1997) show how to adapt existing behaviours automatically to new characters.
Models
One of the virtual robots is a biped. It has two telescoping legs and ball-joint hips like his real counterpart. The other JEB2423
This review is about a field that does not traditionally belong to biological sciences. A branch of computer animation has its mission to create active self-powered objects living artificial lives in the theoretical biology zone. Selected work, of particular interest to biologists, is presented here. These works include animated simulations of legged locomotion, flexible-bodied animals swimming and crawling, artificial fish in virtual ecosystems, automated learning of swimming and the evolution of virtual creatures with respect to morphology, locomotion
and behaviour. The corresponding animations are available for downloading via the Internet. I hope that watching these intriguing pieces of visual simulation will stimulate digitally oriented biologists to seize the interactive methods made possible by ever-increasing computing power.
Key words: animation, artificial life, behaviour, evolution, locomotion, simulation.
Summary
Introduction
VISUAL SIMULATIONS, ARTIFICIAL ANIMALS AND VIRTUAL ECOSYSTEMS
J. E. I. HOKKANEN*
Institute of Biomedicine, PO Box 8, FIN-00014 University of Helsinki, Finland *e-mail: jyrki.hokkanen@helsinki.fi
robot is a gimbal-hipped quadruped. The kangaroo is planar; it has only one leg and one arm. The legs of the kangaroo have rotary joints. The ankle actuator consists of a spring-damper mechanism with an adjustable zero spring length, modelling a muscle acting in series with a springy Achilles tendon. A significant fraction of the energy stored in the spring during leg compression is returned during leg extension. Hopping kangaroos of different sizes are scaled according to dynamic scaling laws. The characters are displayed in Fig. 1.
The human models were constructed from rigid links connected by rotary joints. The mass and moment of inertia of each body part was calculated using data from cadavers. Each internal joint of the model has a very simple muscle model, a torque source, which allows the control algorithms to apply a torque between the two links that form the joint.
Locomotion
For the robots and the kangaroo, six gates are analysed. The running algorithms for the gaits simulated are based on a control algorithm originally developed for one-legged hopping. For example, bipedal running is like one-legged hopping, except that there is an extra idle leg in addition to the active leg. The idle leg is kept short and out of the way while the active leg performs the functions to control forward speed, hopping height and balance. Control algorithms for the kangaroo are essentially the same as for the robot models, with additional provisions for coordinating the joints of the articulated leg and for moving the tail. The tail is made to counter-oscillate with the leg, keeping the angular momentum of the entire system near zero throughout the running cycle.
To keep the forward speed of a legged runner steady, the average point of support (the active foot in contact with the ground) is kept under the average location of the centre of mass of the system. The control system may thus accelerate or decelerate the runner by positioning the foot differently.
During running, tendons and ligaments in the legs and feet stretch during each collision with the ground, converting some of the system’s kinetic energy into elastic strain energy. The stored energy is returned during the next step, when the elastic structures rebound. For the virtual runners, control algorithms allow the mass of the body to rebound on the springy leg during ground collisions and to be drawn back to earth by gravity during the flight phase. The biped and quadruped legs were made springy using spring-damper actuator models for the telescoping joint. The kangaroo leg was made springy by modelling the ankle actuator as a torsional spring-damper with an adjustable resting length. Control of the resting length of the spring is used to inject or remove energy from the system to initiate the oscillation, to modulate it or to stop it.
Depending on the number of legs, the gait and whether there is a tail, the trunk may pitch and roll during running. The long-term attitude of the trunk must be stabilised if the system is to remain upright. The control system implemented regulates the orientation of the trunk by applying torques to the body during the support phase. In the biped and quadruped models, the hip actuators are used to apply the torques required for attitude
control. In the kangaroo model, the knee is used to perform this function.
The control algorithms for human running build on the robot and kangaroo control algorithms by extending them for a system with more controlled degrees of freedom and more stringent requirements on the style of the motion.
State machine
In addition to the control algorithms described above, each implementation uses a ‘state machine’ to track the ongoing behaviour of the model and to synchronise the control actions to the running behaviour. For each gait, the state machine cycles through the legs in correct order. In the case of the bipedal robot, the states are as follows.
Flight: the active leg leaves the ground (interchange active and idle legs, lengthen and position active leg for landing, shorten idle leg).
Loading: the active leg touches the ground (zero active hip torque, keep idle leg short).
Compression: the active leg spring shortens (servo pitch with active hip, keep idle leg short).
Thrust: the active leg spring lengthens (extend active leg, servo pitch with active hip, keep idle leg short).
Unloading: the active leg spring approaches full length (shorten active leg, zero active hip torque, keep idle leg short).
Results
Raibert and Hodgins (1991) compare the behaviour of each animated runner with the behaviour of the physical system it is designed to model. There are many similarities. For example, the extended flight phase (front and rear legs extended) is shorter than the gathered flight phase (legs gathered together under the body) for both the physical and the animated quadruped during bounding. The kangaroo’s body and tail oscillate as it runs, and the magnitudes of the oscillations are similar. A number of differences also exist. The duration of the gathered flight phase of the animated quadruped is twice that of the robot quadruped. The animated biped spends a great deal more time in flight than the robot. The real kangaroo accelerates its feet to the speed of the ground just before they touch the ground, so they do not scuff or have a tangential impact. The animated kangaroo does not do this.
The simulated human runner’s leg motion is similar to that of an athlete on a treadmill, although the simulated runner has a shorter stride and runs with a more upright body posture (Hodgins and Wooten, 1998). The simulated and real vaults differ in that the simulated vaulter keeps a straighter position during the vault and has less motion on landing. When comparing data from the handspring vault performed by 24 gymnasts in the 1987 Pan American Games with the simulated handspring vault, most of the simulated data fell within the range of the measured data.
Automatic adaptation
Hodgins and Pollard (1997) describe an algorithm for automatically adapting existing simulated behaviours to new
characters. Animating a new character is difficult because a control system tuned for one character will not, in general, work on a character with different limb lengths, masses or moments of inertia. The algorithm presented adapts the control system to a new character in two stages. First, the parameters of the control system are scaled on the basis of the sizes, masses and moments of inertia of the new and original characters. A subset of the parameters is then fine-tuned using a search process. To demonstrate the approach, the running motion of a woman, a child and an imaginary character are animated by modifying the control system for a man. In addition, an on-line adaptation with a physically realistic metamorphosis from a running man to a running woman over a period of 20 s is presented (Fig. 2).
Artificial fish, virtual ecosystems
The previous section dealt with legged locomotion. In this
section, virtual soft-bodied animals are studied. The research of Demetri Terzopoulos and his group at the University of Toronto has resulted in a physics-based, virtual marine world inhabited by lifelike artificial life forms that emulate the appearance, motion and behaviour of fish in their natural habitats. Each artificial fish is an autonomous agent with a deformable body actuated by internal muscles, eyes and a mind that includes learning, behaviour, perception and motor centres. Animations are available via the Internet (Terzopoulos, www.cs.toronto.edu/~dt/; Tu, www.dgp.toronto.edu/people/tu/tu.html).
Soft-bodied locomotion
Animals such as snakes, worms, fish and marine mammals, with highly flexible bodies, have been mechanically modelled using spring-mass systems (Miller, 1988; Tu and Terzopoulos, 1994; Grzeszczuk and Terzopoulos, 1995). As an example, let us consider fish. The dynamic model of a fish includes its
Fig. 1. Characters from the animation ‘On the Run’. The ostrich model is described in Borvansky (1991). Image courtesy of Marc Raibert, MIT Leg Laboratory, Massachusetts Institute of Technology.
Fig. 2. Images of the metamorphosis of a man into a woman while running. The metamorphosis occurred over 20 s. Image courtesy of Jessica Hodgins, Georgia Institute of Technology, and Nancy Pollard, Brown University.
muscle actuators and a set of motor controllers. Tu and Terzopoulos (1994) used a biomechanical body model that produces realistic swimming motion using 91 virtual springs. The spring arrangement maintains the structural stability of the body while allowing it to flex. Twelve of the springs running the length of the body also serve as simple muscles. The artificial fish is neutrally buoyant in the virtual water and has a pair of pectoral fins. Additional motor controllers coordinate the actions of the fins. Artificial fish swim, like real fish, by autonomously contracting their muscles. As the body flexes, it displaces virtual fluid, which induces local reaction forces normal to the body. These hydrodynamic forces generate thrust, which propels the fish forward. The model mechanics are governed by Lagrange equations of motion.
The motor controllers coordinate muscle actions to carry out specific motor functions, such as swimming forward, turning left or turning right. They translate natural control parameters, such as the forward speed or angle of the turn, into detailed muscle actions that execute the function.
Automated learning
Grzeszczuk and Terzopoulos (1995) present algorithms that allow a physics-based, muscle-actuated model of an animal to learn from first principles how to control its muscles in order to move in a natural fashion. The approach is based on the notion that natural locomotion patterns are energetically efficient. This allows the problem of learning realistic locomotion to be formalised as one of optimising a class of objective functions, for which there are various solution techniques.
The learning algorithms discover muscle controllers that produce efficient locomotion through optimisation. Muscle contractions that produce forward movements are ‘remembered’. These partial successes then form the basis for the fish’s subsequent improvement in its swimming technique. Their brain’s learning centres also enable these artificial
animals to train themselves to accomplish higher-level tasks, such as manoeuvring to reach a visible target or leaping out of the water (Fig. 3). To this end, abstracted controllers are concatenated in sequence, with each controller slightly overlapping the next (Fig. 4).
Perception
Terzopoulos et al. (1996) present perception strategies that enable artificial marine animals to navigate purposefully through their world by using computer vision algorithms to analyse the image streams acquired by their eyes.
The basic functionality of the active vision system starts with a binocular perspective projection of the coloured three-dimensional world onto the two-three-dimensional retinas of the artificial fish. This projection respects occlusion relationships among objects. It forms spatially non-uniform visual fields with high-resolution foveae and low-resolution peripheries. On the basis of an analysis of the incoming colour retinal image
Fig. 3. SeaWorld tricks learned by the artificial dolphin. Image courtesy of Radek Grzeszczuk, University of Toronto.
CONTROLLERS BASIC ABSTRACTED
Turn up controller Move forward controller Turn down controller
Turn left controller Turn right controller
Turn down controller Move forward controller Turn up controller Turn right controller
HIGHER-ORDER CONTROLLER USED FOR JUMPING OUT OF WATER
Fig. 4. The higher-level controller for jumping out of water is constructed from a set of abstracted basic controllers. Image courtesy of Radek Grzeszczuk, University of Toronto.
stream, the perception centre of the artificial fish’s brain supplies saccade control signals to its eyes to stabilise the visual fields during locomotion, to attend to interesting targets on the basis of colour and to keep targets fixated.
Each eye is controlled independently during stabilisation of a target. Hence, the two retinal images must be correlated to keep them aimed accurately at the target. Once the eyes are aimed at a target, it is straightforward for the fish to estimate the range to the target by triangulation using the gaze angles. The artificial fish is thus able to track and approach other artificial fish.
Group behaviour
The algorithms developed by Tu and Terzopoulos (1994) and Terzopoulos et al. (1994) also emulate the complex group behaviours evident in many aquatic ecosystems.
The innate character of the fish is determined by a set of habit parameters that determine whether or not it likes brightness, darkness, cold, warmth, schooling or is a male/female, etc. The artificial fish has three mental state variables: hunger, libido and fear.
An intention generator first checks the sensory information stream to see whether there is any immediate danger of collision. If any object penetrates the fish’s collision sensitivity region, then the intention generated is to avoid collision. A large sensitivity region results in a ‘timid’ fish that takes evasive action to avoid a potential collision well in advance, while a tight sensitivity region yields a ‘courageous’ fish that takes evasive action only at the last moment.
If there is no immediate danger of collision, the neighbourhood is searched for predators, and the fear state variable for the most dangerous predator is calculated.
If fear is below threshold, the hunger and libido mental state variables are calculated. If the greater of the two exceeds a threshold, the intention generated will be to eat or mate accordingly.
If the above test fails, the intention generator accesses the ambient light and temperature information from the perception system. If the fish’s habits dictate contentment with the ambient conditions, the intention generated will be to wander about; otherwise, it will be to leave the vicinity.
The artificial fish should have some persistence in its intentions, otherwise it will tend to dither, perpetually switching goals. If the current behaviour is interrupted by a high-priority event, the intention generator stores the current intention in a single-item short-term memory.
When the intention generator has selected an intention, it passes control to a behaviour routine. The artificial fish includes eight behaviour routines: avoiding static obstacle, avoiding fish, eating food, mating, leaving, wandering, escaping and schooling. The behaviour routine uses the perceptual data to select a motor controller and to provide it with the appropriate parameters.
For example, the eating-food routine tests the distance from the fish’s mouth to the food. If the distance is greater than some threshold value, the subroutine chasing-target is invoked. The chasing-target subroutine guides a fish as it swims towards a
goal. When the distance is less than the threshold value, the subroutine suck-in is activated in which a ‘vacuum’ force is calculated and then exerted on the food.
At each time step, the intention generator issues an intention based on the fish’s habits, mental state and incoming sensory information. It then chooses and executes a behaviour routine, which in turn runs the appropriate motor controllers.
Evolving moving and competing creatures
Karl Sims (1994a,b) studies the evolution of virtual creatures that move and compete in physically simulated three-dimensional worlds. The morphology of these creatures and the neural systems for controlling their muscle forces are both genetically determined, and the morphology and behaviour can adapt to each other as they evolve simultaneously. When simulated evolutions are performed, a variety of locomotion and competition strategies emerge. The corresponding animation is available for downloading (Sims, www.genarts.com/karl/index.html).
Morphology
In artificial evolution, a genotype is a coded representation of a possible individual. Populations of genotypes are read to produce phenotypes that are then evaluated according to some fitness criteria and selectively reproduced. Copying, mutating and/or combining the genotypes of the fittest individuals generate new genotypes and, as the cycle repeats, the population should ascend to progressively higher levels of fitness.
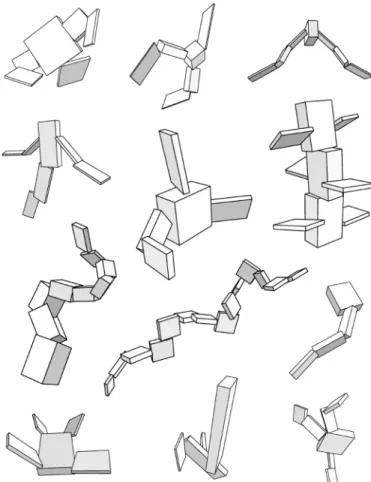
The phenotype embodiments of Karl Sims’ virtual creatures are hierarchies of articulated three-dimensional rigid parts. The genetic representation of this morphology is a directed graph of nodes and connections. Each node in the graph contains information describing a rigid part. A set of local neurons is also included in each node and will be explained below. Finally, a node contains a set of connections to other nodes. Fig. 5 shows some simple hand-designed graph topologies and resulting phenotype morphologies. Note that the parameters in the nodes and connections are not shown for the genotype even though they affect the morphology of the phenotype.
Control
Each sensor is contained within a specific part of the body. Three different types of sensor were used: joint angle sensors, contact sensors and photosensors.
Internal neural nodes allow a creature to have an internal state beyond its sensor values and to be affected by its history. Different neural nodes can perform diverse functions on their inputs to generate their output signals. Each input contains a connection to another neuron or a sensor from which to receive a value.
Each effector contains a connection from a neuron or a sensor from which to receive a value. This input value is exerted as a joint force. Each effector is given a maximum
strength proportional to the cross-sectional area of the two parts it joins.
Selection
Virtual creatures are evolved by optimising for a specific task. A creature is grown from its genetic description and is then placed in a dynamically simulated virtual world. After a certain duration of virtual time, a fitness value is assigned that corresponds to the success level of that behaviour. If a creature has a high fitness relative to the rest of the population, it will be selected for survival and reproduction. Sims (1994a) used four different types of fitness measures.
Swimming speed is measured by the distance travelled by the creature’s centre of mass per unit time. Straight swimming is rewarded over circling by using the maximum distance from the initial centre of mass.
The term ‘walking’ is used loosely here to indicate any form of land locomotion. Horizontal speed is used as the selection criterion.
Jumping behaviour can be selected for by measuring the maximum height above the ground of the lowest part of the creature. An alternative method is to use the average height of the lowest part of the creature during the duration of simulation.
The fourth evaluation method is to select for creatures that can follow a light source. Several trials are run with the light source in different locations, and the speeds at which a creature moves towards it are averaged to give the fitness value.
In natural evolutionary systems, the measure of fitness is not constant. The fitness of an individual is highly dependent on the specific behaviours of other individuals currently in the population. Sims (1994b) studied a simulated contest.
A single cube is placed in the centre of the world, and the creatures start on opposite sides of the cube. The winner is the creature that has the most control over the cube after a certain duration of simulated time. A creature achieves a higher fitness score by being closer to the cube, but also gets a higher score when its opponent is further away. The most interesting results occurred when each individual was chosen to compete, once per generation, against the same opponent. The individual with the highest fitness from the previous generation was chosen as the one to beat.
Evolution
The evolution of virtual creatures is begun by first creating an initial population of genotypes by random generation. A survival ratio determines the percentage of the population that will survive each generation. Population sizes were typically 300, and the survival ratio was 1/5. For each generation, creatures are grown from their genotypes, and their fitness values are measured. The individuals whose fitness falls within the survival percentile are then reproduced, and their offspring fill the slots of those individuals that did not survive. The number of offspring that each surviving individual generates is proportional to its fitness. The survivors are kept in the population for the next generation, and the total size of the population is maintained. Offspring are generated from the surviving creatures by copying and combining their directed graph genotypes. When these graphs are reproduced, they are subjected to probabilistic variation or mutation. The evolution of a three-segmented swimmer is shown in Fig. 6.
Sexual reproduction allows components from more than one parent to be combined into new offspring. This permits features to evolve independently and later to be merged into a single individual. Two different methods for mating-directed graphs are used. The reproduction method is chosen at random for each offspring.
Results
The swimming fitness measure produced a large number of simple paddling and tail-wagging creatures. A few creatures pulled themselves through the water using specialised sculling appendages. Some used two symmetrical flippers or even large numbers of similar flippers to propel themselves, and several multi-segmented watersnake creatures evolved that wound through the water with sinusoidal motions. Some of the evolved swimmers are shown in Fig. 7.
The walking fitness measure also produced a surprising number of simple creatures that could shuffle or hobble along at fairly high speeds. Some walk with lizard-like gaits using
Fig. 5. Designed examples of genotype graphs and corresponding creature morphologies. Image courtesy of Karl Sims.
(Head)
(Body) segment)(Limb
Genotype: directed graph Phenotype: hierarchy of three-dimensional parts (Leg segment) (Body segment) (Segment)
the corners of their parts. Some simply wag an appendage in the air to rock back and forth in just the right manner to move forward. A number of more complex creatures push or pull themselves along, inchworm-style. Others use one or more leg-like appendages to crawl or walk successfully. Some hopping creatures even emerged that raise and lower arm-like structures to bound along at fairly high speeds.
The jumping fitness measure did not seem to produce as many different strategies as the swimming and walking optimisations, but a number of simple jumping creatures did emerge.
The light-following fitness measure produced a wide variety of creatures that can swim or walk towards a light source. In
the water environment, some developed steering fins that turn them towards the light using photosensor inputs. Others adjust the angle of their paddles appropriately as they oscillate along. Creatures that evolved in one physical world can be placed in another and evolved further. An evolved watersnake, for example, was placed on land and then evolved to crawl instead of swim.
Many independent evolutions were performed using the competition pattern. Some creatures extended arms out onto the cube (Fig. 8). Others could crawl inchworm-style or roll towards the cube, and a few even developed leg-like appendages that they used to walk towards it. Some creatures pushed their opponent away from the cube, and some moved the cube away from its initial location and then followed it. For example, an arm-like creature pushed the cube aside and then used photosensors adaptively to follow it. If its opponent moved the cube in a different direction, it would successfully grope towards the new location. Several two-armed creatures used the strategy of batting the cube to the side with one arm and catching it with the other arm. A larger creature won by a large margin against some opponents because it could literally walk away with the cube, but it did not initially reach the cube very quickly and tended to lose against faster opponents.
Fig. 7. Creatures evolved for swimming. Image courtesy of Karl Sims.
Fig. 8. Contestants going for the green cube. Image courtesy of Karl Sims.
Fig. 6. Ancestors of the three-segmented flipper. Image courtesy of Karl Sims.
I would like to thank Radek Grzeszczuk, Jessica Hodgins, Marc Raibert and Karl Sims for correspondence and for providing the illustrations.
References
Alexander, R. McN. and Vernon, A. (1975). The mechanics of hopping by kangaroos (Macropodidas). J. Zool., Lond. 177, 265–303.
Borvansky, L. (1991). Dynamic simulation of the running motion of an ostrich. MS thesis, Department of Electrical Engineering and Computer Science, Massachusetts Institute of Technology, Cambridge, Massachusetts, USA.
Grzeszczuk, R. and Terzopoulos, D. (1995). Automated learning of muscle-actuated locomotion through control abstraction. Proc.
ACM SIGGRAPH, pp. 63–70.
Hodgins, J. K. and Pollard, N. S. (1997). Adapting simulated behaviors for new characters. Proc. ACM SIGGRAPH, pp. 153–162.
Hodgins, J. K. and Wooten, W. L. (1998). Animating human athletes. In Robotics Research: The Eighth International
Symposium (ed. Y. Shirai and S. Hirose), pp. 356–367. Berlin:
Springer-Verlag.
Miller, G. S. P. (1988). The motion dynamics of snakes and worms.
Computer Graphics 22, 169–177.
Raibert, M. H. and Hodgins, J. K. (1991). Animation of dynamic legged locomotion. Computer Graphics 25, 349–358.
Sims, K. (1994a). Evolving virtual creatures. Proc. ACM SIGGRAPH, pp. 15–22.
Sims, K. (1994b). Evolving three-dimensional morphology and behavior by competition. Artificial Life IV Proceedings (ed. R. A. Brooks and P. Maes), pp. 28–39. Cambridge, MA: The MIT Press. Terzopoulos, D., Rabie, T. and Grzeszczuk, R. (1996). Perception and learning in artificial animals. Artificial Life V Proceedings (ed. C. G. Langton and K. Shimohara), pp. 313–320. Cambridge, MA: The MIT Press.
Terzopoulos, D., Tu, X. and Grzeszczuk, R. (1994). Artificial fishes: Autonomous locomotion, perception, behavior and learning in a simulated physical world. Artificial Life 1, 327–351.
Tu, X. and Terzopoulos, D. (1994). Artificial fishes: Physics, locomotion, perception, behavior. Proc. ACM SIGGRAPH, pp. 43–50.