of August 16, 2019.
This information is current as
Heteroprotein Complexes
Expression through Assembly of Homo- or
3
Φ
1 and IFN
Φ
Differentially Regulate IFN
Zhang, Ya-Wei Xiong, Min Wu and Jian-Fang Gui
Hui Feng, Qi-Min Zhang, Yi-Bing Zhang, Zhi Li, Jun
http://www.jimmunol.org/content/197/5/1893
doi: 10.4049/jimmunol.1600159
August 2016;
2016; 197:1893-1904; Prepublished online 5
J Immunol
References
http://www.jimmunol.org/content/197/5/1893.full#ref-list-1
, 23 of which you can access for free at:
cites 57 articles
This article
average
*
4 weeks from acceptance to publication
Fast Publication!
•
Every submission reviewed by practicing scientists
No Triage!
•
from submission to initial decision
Rapid Reviews! 30 days*
•
Submit online.
?
The JI
Why
Subscription
http://jimmunol.org/subscription
is online at:
The Journal of Immunology
Information about subscribing to
Permissions
http://www.aai.org/About/Publications/JI/copyright.html
Submit copyright permission requests at:
Email Alerts
http://jimmunol.org/alerts
Receive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606.
Immunologists, Inc. All rights reserved.
Copyright © 2016 by The American Association of
1451 Rockville Pike, Suite 650, Rockville, MD 20852
The American Association of Immunologists, Inc.,
is published twice each month by
The Journal of Immunology
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
Downloaded from
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
Hui Feng,
1,2Qi-Min Zhang,
1Yi-Bing Zhang, Zhi Li, Jun Zhang, Ya-Wei Xiong,
Min Wu, and Jian-Fang Gui
In mammals, IFN regulatory factor (IRF)1, IRF3, and IRF7 are three critical transcription factors that are pivotal for cooperative regulation of the type I IFN response. In this study, we explored the relative contribution of zebrafish (Danio rerio) IRF1 (DrIRF1), IRF3 (DrIRF3), and IRF7 (DrIRF7) (DrIRF1/3/7) to zebrafish IFNF1 (DrIFNF1) and IFNF3 (DrIFNF3) (DrIFNF1/3) activa-tion. Following spring viremia of carp virus infection, DrIFNF1/3 and DrIRF1/3/7 transcripts are significantly induced in zebra-fish tissues, which correlates with the replication of spring viremia of carp virus. DrIRF1/3/7 selectively bind to the IRF-binding element/IFN-stimulated regulatory element sites of DrIFNF1/3 promoters, with the exception that DrIRF3 has no preference for two IRF-binding element/IFN-stimulated regulatory element motifs within the DrIFNF3 promoter. Consistently, DrIRF3 alone activates DrIFNF1, but not DrIFNF3; DrIRF7 predominantly stimulates DrIFNF3; and DrIRF1 has similar potential to DrIFNF1 and DrIFNF3. Strikingly, DrIRF3 facilitates the binding of DrIRF1 and DrIRF7 to both zebrafish IFN promoters, and so does DrIRF7 for the binding of DrIRF1, particularly to the DrIFNF3 promoter. These binding properties correlate with differential responses of DrIFNF1 and DrIFNF3 to the combinatory stimulation of DrIRF1/3/7, depending on their relative amounts. Similar to the dual roles of human IRF3 in regulating IRF7-activated IFNa genes, DrIRF3 exerts dual effects on DrIRF1-mediated DrIFNF3 gene expression: an inhibitory effect at lower concentrations and a synergistic effect at higher concentrations. These data provide evidence that fish and mammals have evolved a similar IRF-dependent regulatory mechanism fine-tuning IFN gene activation. The Journal of Immunology, 2016, 197: 1893–1904.
I
n mammals, type I IFNs (primarily IFN-a/b) are induced to boost the immune response, protecting hosts from viral and nonviral pathogens (1). Transcriptional initiation of type I IFN genes depends on timely recognition of pathogenic signals through host pattern recognition receptors within endosomes (such as TLR2/3/4/7/8/9) or in the cytosol (such as cytosolic RNA sensors, RNA helicases, and cytosolic DNA sensors) (2). Such recognition events trigger distinct signaling pathways that finallyconverge on the activation of a family of transcriptional factors called IFN regulatory factors (IRFs) (1). The IRF family includes 9 members in mammals, 10 members in birds, and 11 members in fish (3). All IRFs contain a conserved N-terminal DNA-binding domain (DBD) characterized by five tryptophan repeat elements. By folding to a helix-turn-helix motif, DBD specifically recog-nizes and binds to a similar DNA sequence termed an IRF-binding element (IRF-E)/IFN-stimulated response element (ISRE) within the promoters of type I IFN genes and IFN-stimulated genes (4–7). In addition to DBD, each family member contains a unique C-terminal IRF-associated domain that accounts for interactions with other IRFs and transcription factors (1).
Among the IRF family, IRF1, IRF3, and IRF7 are characterized early in regulating type I IFN expression (1). In most cell types, the IRF3/7-dependent IFN response is activated through the cytosolic RNA-mediated mitochondrial antiviral signaling protein path-way and the cytosolic DNA-mediated mediator of IRF3 activator pathway (1, 2). IRF3 and IRF7 also contribute to TLR3-dependent IFN expression, and IRF3 is primarily responsible for TLR4-IFN signaling through the TIR domain-containing adaptor protein pathway (8). However, TLR2/7/8/9-mediated induction of type I IFN appears to use distinct IRFs in a cell type–specific fashion but through a common adaptor called MyD88 (9–12). For example, the TLR9-triggered type I IFN response exclusively requires IRF7 in plasmacytoid dendritic cells (DCs) (9), IRF1 in conventional DCs (cDCs) (13, 14), and IRF3 in B lymphocytes (15); IRF1 and IRF7 are necessary for the TLR2-driven type I IFN response in immune cells, including macrophages and cDCs (12), and for the TLR7/9-triggered IFN response in cDCs (11). Therefore, like IRF3 and IRF7, IRF1 is another critical regulator of the type I IFN response (16, 17).
State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, University of Chinese Academy of Sciences, Wuhan 430072, China
1
H.F. and Q.-M.Z. contributed equally to this work.
2Current address: Lineberger Comprehensive Cancer Center, University of North Carolina at Chapel Hill, Chapel Hill, NC.
Received for publication January 28, 2016. Accepted for publication July 4, 2016.
This work was supported by Grants 31272690 and 31572646 from the National Natural Science Foundation and Grant XDA08010207 from the Strategic Priority Research Program of the Chinese Academy of Sciences.
Address correspondence and reprint requests to Dr. Yi-Bing Zhang, State Key Lab-oratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Uni-versity of Chinese Academy of Sciences, Wuhan 430072, China. E-mail address: [email protected]
Abbreviations used in this article: cDC, conventional DC; co-IP, coimmunoprecipita-tion; DBD, DNA-binding domain; DC, dendritic cell; DrIFNF1, zebrafish IFNF1; DrIFNF1/3, DrIFNF1 and DrIFNF3; DrIFNF3, zebrafish IFNF3; DrIRF1, zebrafish (D. rerio) IRF1; DrIRF1/3/7, zebrafish IRF1, IRF3, and IRF7; DrIRF3, zebrafish IRF3; DrIRF7, zebrafish IRF7; DrMyD88, zebrafish myeloid differentiation factor 88; HA, hemagglutinin; IRF, IFN regulatory factor; IRF-E, IRF-binding element; IRF1-HA, HA-tagged DrIRF1; IRF3-HA, HA-tagged DrIRF3; IRF7-HA, HA-tagged DrIRF7; ISRE, IFN-stimulated response element; lambda PPase, lambda protein phosphatase; ORF, open reading frame; poly(I:C), polyinosinic-polycytidylic acid; qRT-PCR, quan-titative RT-PCR; rIFN, recombinant crucian carp IFN; RLR, retinoic acid–inducible gene I–like receptor; SVCV, spring viremia of carp virus; ZFL, zebrafish liver cell.
CopyrightÓ2016 by The American Association of Immunologists, Inc. 0022-1767/16/$30.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1600159
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
Interestingly, accumulating data showed that other IRF members also participate in retinoic acid–inducible gene I–like receptor (RLR)-IFN or TLR-IFN signaling. IRF8 directs the IFN response, together with IRF3/7, in DCs (18) and with IRF3 in human blood monocytes (19). IRF3, IRF5, and IRF7 coordinately act in mye-loid DCs downstream of mitochondrial antiviral signaling protein signaling (20). Following Lactobacillus acidophilus infection, MyD88 triggers IFN-b responses in DCs through a mechanism involving IRF1, IRF3, and IRF7 (21). These results suggest that multiple IRF members often work together to coordinately reg-ulate type I IFN gene activation, although the molecular mecha-nisms involved remain unclear.
Studies during the past decade have led to tremendous ad-vances in the understanding of fish IFN responses (22–24). Despite functional similarity, fish IFNs are classified into group I and group II IFNs (25–27), and they signal through distinct receptors that are different from mammalian type I IFNRs (26). Neverthe-less, fish IRF family members exhibit an orthologous relationship with mammalian homologs (3, 28) and play conserved roles in the host antiviral response (28–34). Notably, IRF3 and IRF7 are necessary for the RLR-mediated IFN response in zebrafish (Danio rerio) (29, 32), as well as for the MyD88-mediated IFN response in Atlantic salmon (Salmo salar) (35); zebrafish (D. rerio) IRF1 (DrIRF1) synergizes with MyD88 to regulate the expression of zebrafish IFNF3 (DrIFNF3) (36). Furthermore, fish IRF3 is an IFN-inducible protein and is translocated to the nucleus following virus infection or IFN treatment (28). These unique findings prompted us to explore the coordination of fish IRF3 with other IRF members, such as IRF1 and IRF7, on the IFN antiviral re-sponse.
In the current study, we determined the relative contribution of zebrafish IRF1, IRF3, and IRF7 (DrIRF1/3/7) to distinct zebra-fish IFN activation. RT-PCR revealed the expression of zebrazebra-fish IFNF1 (DrIFNF1) and DrIFNF3 (DrIFNF1/3), together with DrIRF1/3/7, in some zebrafish tissues, and coimmunoprecipitation (co-IP) showed the interaction of DrIRF1/3/7 and MyD88, implying that DrIRF1/3/7 might coregulate DrIFNF1 and DrIFNF3 gene activation. Based on in vitro pull-down assays and promoter acti-vation analyses, we found that DrIRF1/3/7 coordinately bound to IRF-E/ISRE motifs within DrIFN promoters as homo- or hetero-protein complexes to synergistically stimulate or downregulate the expression of DrIFNF1 and DrIFNF3, depending on their combi-natory binding and the relative amounts.
Materials and Methods
Cells, virus, and zebrafishCrucian carp (Carassius auratusL.) blastula embryonic cells, epithe-lioma papulosum cyprini cells, and zebrafish liver cells (ZFLs) were cultured as described previously (37, 38). Spring viremia of carp virus (SVCV) was propagated and titered in epithelioma papulosum cyprini cells. Zebrafish (D. rerio) was maintained and infected i.p. with 50ml of SVCV (108TCID50/ml)/fish, as described (36).
Plasmids
For in vitro translation and co-IP assays, the expression plasmids were generated by inserting the open reading frames (ORFs) of DrIRF1, zebrafish IRF3 (DrIRF3), and zebrafish IRF7 (DrIRF7) into a pCS2(+) vector that had precloned the HA/Myc coding sequence into the XbaI restriction site. For luciferase assays, the ORFs of DrIRF1, DrIRF3, and DrIRF7 were cloned into the EcoRI and NotI sites of the pcDNA3.1(+)
vector. For prokaryotic expression, the ORF of DrIRF3 was cloned into EcoRI and NotI sites of pGEX4T-1. All constructs were confirmed by sequencing. Other plasmids, including zebrafish IFN promoter-driven luciferase plasmids DrIFNF1pro-luc and DrIFNF3pro-luc, zebrafish myeloid differentiation factor 88 (DrMyD88)-Flag were described pre-viously (29, 36).
Luciferase activity assays
Stimulation, transfection, and subsequent luciferase activity assays were performed as previously described (28, 29, 37).
DNA pull-down assays
DNA pull-down assays were performed as described previously (36, 39), with in vitro–translated IRF proteins, prokaryotically expressed proteins, or cell lysates of HEK293T transfected or not with IRF expression plasmids. Zebrafish IRF proteins were in vitro–translated in the TNT SP6 Quick Coupled Transcription/Translation System supplemented with 40 mM KCl (Promega) or expressed as recombinant proteins by a prokaryotic expression system, according to previous studies (27, 28), or they were overexpressed as tag-fused proteins by transfection of HEK293T cells for 24 h. Briefly, 25ml of individual translated products, 4mg of recombinant products, or an adequate amount of IRF-overexpressing HEK293T cell lysates (one fifth of a 10 cm-dish, unless otherwise indicated), together with 1mg of biotinylated promoter dsDNA, was incubated in HKMG buffer at 4˚C for 24 h. The bead-bound DNA-protein complex was captured with M-280 streptavidin-coupled Dynabeads (Invitrogen) and washed with incubation buffer five times, fol-lowed by further Western blotting analysis.
Quantitative RT-PCR
Tissues were sampled from healthy zebrafish or ones that had been infected for 48 h with SVCV, and total RNA was extracted for further expression analysis of zebrafish IRFs, zebrafish IFNs, and SVCV genes, including L, N, and G, by quantitative RT-PCR (qRT-PCR). qRT-PCR was also used to detect zebrafish IRF and IFN expression in ZFLs, which were seeded in six-well plates and transfected with polyinosinic-polycytidylic acid [poly(I:C); 2 mg/ml] or treated with SVCV (1000 TCID50/ml) and recombinant crucian
carp IFN (rIFN) protein (5 ng/ml). rIFN protein was generated by a pro-karyotic expression system (27). qRT-PCR was performed in a DNA En-gine Chromo4 Real-Time PCR System with SYBR Green Real-Time PCR Master Mix (both from Bio-Rad) (27). All samples were analyzed in trip-licates, and the expression values were normalized tob-actin. The following primer sequences were used: DrIRF1 (F: 59 -CCGGCGGATGAAGGGTC-TGT-39, R: 59-CGTTGCATGTGCTGTCAGGG-39); DrIRF3 (F: 59 -CAA-AACCGCTGTTCGTGCC-39, R: 59-CATCGTCGCTGTTGGAGTCCT-39); DrIRF7 (F: 59-AGGCAGTTCAACGTCAGCTACCAT-39, R: 59 -TTCCAC-CAAGTTGAGCAATTCCAG-39); DrIFNF1 (universal primers F: 59 -ACG-ACAGAATCTCTGAACCT-39, R: 59-GTCAGGACTAAAAACTTCAC-39); the secreted DrIFNF1 isoform (F: 59-TGAGAACTCAAATGTGGACCT-39, R: 59-GTCCTCCACCTTTGACTTGT-39); the intracellular DrIFNF1 iso-form (F: 59-ACGGCAGCCTGAAATACGTT-39, R: 59 -GTCCTCCACCT-TTGACTTGT-39); DrIFNF3 (F: 59-TTCTGCTTTGTGCAGGTTTG-39, R: 59-GGTATAGAAACGCGGTCGTC-39); SVCV N gene (F: 59 -GGTGCGA-GTAGAAGACATCCCCG-39, R: 59 -GTAATTCCCATCATTGCCCCAGA-C-39); SVCV L gene (F: 59-CAAGTTCACAATCGGGAAGACGC-39, R: 59-CCAGTTGCTTGTTGGCTTATCCG-39); and SVCV G gene (F: 59 -CC-ATTCTGTTCATTTGGAGCCGTA-39, R: 59 -AATTTCATTCGACAAGA-CCCCC-39).
Co-IP and Western blots
Co-IP assays and Western blots were performed, as previously described (28, 29, 37). The following commercial Abs were used: anti-myc, HA, anti-Flag (Cell Signaling Technology, Danvers, MA), and anti-GST (Merck Millipore, Darmstadt, Germany). Anti-crucian carp IRF3 and IRF7 Abs were raised by immunization of rabbit with prokaryotically expressed crucian carp IRF3-DBD or IRF7-DBD, as described previously (28). We confirmed the binding specificity of both fish IRF Abs to zebrafish IRF3 and IRF7.
Results
Expression patterns of DrIRF1/3/7 with DrIFNF1 and DrIFNF3 in stimulated cells and tissues
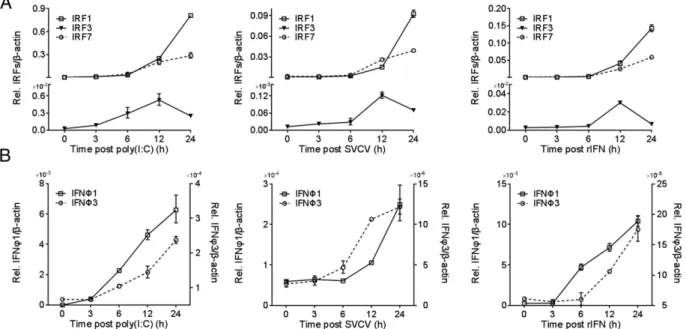
Previous results showed that DrIRF1/3/7 upregulate the expres-sion of DrIFNF1 and DrIFNF3 (29, 36). To further delineate the functional link among them, we determined their expression pat-terns in ZFLs and selected zebrafish tissues. As shown in Fig. 1, all five molecules were constitutively transcribed in nonstimulated ZFLs, albeit at varied expression levels; three stimuli, including poly(I:C), SVCV, and rIFN protein, induced their expression quickly over a 24-h time course. DrIRF1 and DrIRF7 were in-duced continuously, displaying similar levels during early
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
lation and obviously different expression values afterward (6 h posttreatment), whereas DrIRF3 transcripts reached a peak at 12 h posttreatment and decreased slightly thereafter (Fig. 1A). Notably, DrIRF1 and DrIRF7 were transcriptionally expressed much more than IRF3 in nonstimulated and stimulated cells (Fig. 1A). DrIFNF1 and DrIFNF3 were also induced under the same conditions, with a higher constitutive expression and a more rapid induction for DrIFNF1 (Fig. 1B).
Detection of these five transcripts in 13 tissues of healthy zebrafish showed that DrIRF1 had the highest level of constitutive expression among the three IRFs (Fig. 2A). DrIFNF1 has two isoforms (40, 41). RT-PCR analysis showed that the intracellular DrIFNF1 isoform (without a secretion peptide) was more sig-nificantly expressed than the secreted one (with a secretion pep-tide) in healthy tissues, and DrIFNF3 transcripts were richer than secreted DrIFNF1. Despite the varied expression levels, IRF and IFN molecules were generally abundant in thymus, gill, heart, and muscle, whereas their abundance was moderate or poor in the rest of the tissues (Fig. 2A). After SVCV infection, these molecules were transcriptionally upregulated to different extents in all tis-sues, predominantly liver, intestine, and skin (Fig. 2B). With regard to the two DrIFNF1 isoforms, although the secreted one had low basal expression in healthy tissues (Fig. 2A), it was more strongly induced by viral infection than the intracellular one and DrIFNF3 (Fig. 2B). Subsequent analysis of three SVCV gene transcripts, including L, N, and G, showed relatively high ex-pression in hypophysis, thymus, gill, liver, and muscle (Fig. 2C), in which zebrafish IFN transcription was also induced (Fig. 2B). These results suggested that SVCV infection induces simulta-neous expression of DrIFNF1/3 and DrIRF1/3/7 in zebrafish cells and tissues.
Interaction of zebrafish IRF1/3/7 with MyD88
In mammals, IRF1/7 interact with MyD88 (9, 13, 14), and so does mammalian IRF3 (42). To determine whether DrIRF3 and DrIRF7, like salmon homologs (35), interact with DrMyD88, we performed co-IP assays in HEK293T cells co-overexpressing
FLAG-tagged DrMyD88 with hemagglutinin (HA)-tagged DrIRF1 (IRF1-HA), HA-tagged DrIRF3 (IRF3-HA), or HA-tagged DrIRF7 (IRF7-HA). Consistent with our previous results (36), DrIRF1 was physically associated with DrMyD88, because the protein complex immunoprecipitated by FLAG Ab was also recognized by anti-HA Ab (Fig. 3A, upper panels); however, an unrelated protein (EGFP) was not under the same condition (Fig. 3A, lower panels). Similar results were observed for transfection of DrMyD88 with either DrIRF3 or DrIRF7 (Fig. 3B, 3C). Combined with the ex-pression analysis of DrIFN1/3/7 and DrIFNF1/3 (Figs. 1, 2), these results suggest that zebrafish IRF1, IRF3, and IRF7 might simulta-neously regulate IFN gene expression in certain tissues or cell types through similar signaling pathways, such as the MyD88 pathway.
Binding of DrIRF1/3/7 to IRF-E/ISRE motifs within promoters of DrIFNF1/3
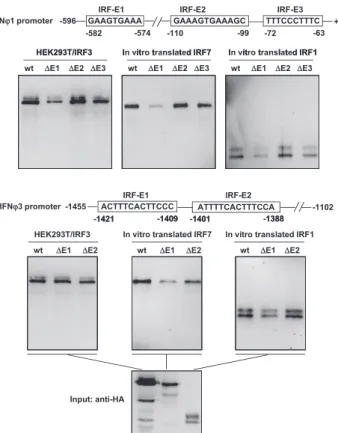
It is well-known that mammalian IRF-activated type I IFN ex-pression depends on direct binding to IFN promoter regions (1). We confirmed that DrIRF1 binds to DrIFNF1 (corresponding to the secreted isoform hereafter, unless indicated) and DrIFNF3 pro-moters (36). Thus, we asked whether this might be the case for DrIRF3 and DrIRF7 as well. To this end, DNA pull-down assays were performed by incubation of biotin-labeled promoter DNAs of DrIFNF1 (2596 to +38) or DrIFNF3 (21455 to21102), which bear three or two putative ISRE/IRF-E–like motifs (36), with in vitro–translated DrIRF1/3/7 proteins to determine whether each of them was able to bind to IFN promoters. As shown in Fig. 4A, in vitro translation of an HA-tagged DrIRF1 plasmid (IRF1-HA) generated a theoretically sized protein and a large-sized one; Western blot analysis of promoter-bound products showed that both forms of DrIRF1 effectively bound to IFNF1/3 promoters. Two protein forms were also observed for in vitro–translated DrIRF7 or DrIRF3, with a main one corresponding to the expected m.w. and a weak one that is likely to be a degraded form. Only the theoretically sized DrIRF7 bound to IFN promoters and, unex-pectedly, none of the in vitro–translated DrIRF3 do so under the same conditions (Fig. 4A).
FIGURE 1. RT-PCR analysis of zebrafish IRF1, IRF3, and IRF7 together with zebrafish IFNF1 and IFNF3 in ZFLs. ZFLs seeded in six-well plates were transfected with poly(I:C) (2mg/ml) or treated with SVCV (1000 TCID50/ml) or recombinant IFN protein (5 ng/ml). At the indicated time points, the cells
were sampled for real-time PCR analysis of DrIRF1/3/7 (A) and DrIFNF1/3 (B) expression by different stimuli. Amplification of DrIFNF1 was conducted by universal primers. The expression level is relative to the corresponding expression level ofb-actin. Error bars represent SDs obtained by measuring each sample in triplicate.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
To further characterize the binding nature of DrIRF3, DrIRF3 (IRF3-HA) or DrIRF7 (IRF7-HA) was overexpressed in HEK293T cells, from which the cell lysates were used to repeat DNA pull-down assays. Western blot detection of the transfected cell lysates showed a theoretically sized form and at least one large-sized form of DrIRF3 (Fig. 4B). As a positive control, overexpression of DrIRF7 resulted in an intense theoretically sized protein that bound to DrIFNF1/3 promoters and sometimes a faint slow-migrating band that was undetectable in the promoter-bound products. In contrast to in vitro–translated DrIRF3, all overexpressed DrIRF3 forms bound to both zebrafish IFN promoters (Fig. 4B). Further, a recombinant fusion protein (GST-IRF3) was made by a prokaryotic expression system (Fig. 4C, left panels), and, expectedly, GST-IRF3 exhibited a strong binding affinity to DrIFN promoters (Fig. 4C, right panel). This prokaryotic expression system simultaneously produced a degraded form of GST-IRF3 that retained the IFN promoter binding affinity due to the existence of N-terminal DBD (Fig. 4C). Thus, DrIRF3 physically binds to zebrafish IFN promoters.
In subsequent assays, we constructed a set of IFN promoter mutants lacking the individual IRF-E/ISRE motif to examine the exact binding affinity of DrIRF1, DrIRF3, and DrIRF7 (Fig. 5). Compared with the amounts of three IRF proteins bound to the wild-type DrIFNF1 promoter, significantly less IRF protein was pulled down by the DrIFNF1 promoter lacking the distant IRF-E1
motif. Deleting the proximal IRF-E3 motif caused a moderate re-duction in the band intensities of three DrIRFs, whereas deleting the middle IRF-E2 motif only led to a slight decrease in DrIRF3 binding (Fig. 5, upper panels). With regard to the DrIFNF3 pro-moter, deleting the distant IRF-E1 motif led to a significant re-duction in DrIRF7 binding, a moderate rere-duction in DrIRF1 binding, and a slight decrease in DrIRF3 binding. Deleting the proximal IRF-E2 motif caused a moderate reduction in DrIRF7 binding, a slight decrease in DrIRF3 binding, and no obvious effect on DrIRF1 binding (Fig. 5, lower panels). Collectively, these data suggest that DrIRF1, DrIRF3, and DrIRF7 differentially bind to the IRF-E/ISRE motifs within DrIFNF1 and DrIFNF3 promoters.
Physical binding of DrIRF1/3 to IFN promoters independently of phosphorylation
It was possible that the slow-migrating protein of DrIRF3 in the overexpressed HEK293T cells represented a phosphorylated form. To address this and determine whether such phosphorylation was essential for the binding affinity of IRF proteins to IFN promoters, lambda protein phosphatase (lambda PPase) was used to treat DrIRF1 and DrIRF3 before or after DNA pull-down assays. As shown in Fig. 6, in vitro translation gave two forms of DrIRF1; transfection of HEK293T cells with DrIRF3 plasmid generated three migrating protein forms (i.e., one main band at the expected FIGURE 2. RT-PCR analysis of DrIRF1/3/7 and DrIFNF1/3 in healthy or infected zebrafish. The indicated tissues were collected from wild-type zebrafish (A) or from zebrafish injected i.p. with PBS or SVCV (53106TCID50) (B) for qRT-PCR detection of DrIFNF1/3 and DrIRF1/3/7. Universal
primers were used for analysis of total DrIFNF1, and specific primers were used for the secreted or intracellular isoforms. (C) At the same time, the expression of SVCV genes G, L, and N was determined in virally infected tissues. The expression level is shown relative to the corresponding expression level ofb-actin (A) or as fold induction relative to that in corresponding tissues injected with PBS i.p., which was set to 1 (BandC). Error bars represent SDs obtained by measuring each sample in triplicate.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
m.w. was accompanied by two weak, slow-migrating ones). Incubation of lambda PPase did not affect the mobility of any rapid-migrating protein, but it resulted in the disappearance of the more slowly migrating form of DrIRF1 and only the most slowly migrating one of DrIRF3. Regardless of whether lambda PPase treatment occurred before or after incubation with IFN
promoters, DNA pull-down assays showed that neither IRF1 nor IRF3 lost their binding affinities. As control, pretreatment of DrIRF7 with the protein phosphatase did not influence its binding to IFN promoters. These results suggest that physical binding of DrIRF1/3 to IFN promoters is a phosphorylation-independent action.
FIGURE 3. Immunoprecipitation anal-ysis of the association between zebrafish IRF1 and MyD88. HEK293T cells, seeded in 10-cm dishes, were cotransfected with 5mg of FLAG-tagged DrMyD88 and 5 mg of IRF1-HA (A), IRF3-HA (B), or IRF7-HA (C) or empty vector pCS2+ (A–C, upper panels) and EGFP-HA (A–C, lower panels) as control. Twenty-four hours later, the trans-fected cells were immunoprecipitated with anti-FLAG Ab, followed by analysis of the immunoprecipitates by Western blotting.
FIGURE 4. The binding affinity of zebrafish IRF1, IRF3, and IRF7 to zebrafish IFNF1 and IFNF3 promoters. (A) In vitro-translated DrIRF1 and DrIRF7, but not DrIRF3, bind to both zebrafish IFN promoters by pull-down assays. A total of 1mg of biotin-labeled DrIFNF1 (2596 to +38) or DrIFNF3 (21455∼21102) promoter DNAs was incubated with the indicated in vitro–translated DrIRF1, DrIRF3, or DrIRF7 at 10-fold amounts of input. The bead-bound DNA-protein complex was detected by Ab specific to HA tag. (B) DrIRF3 and DrIERF7 overexpressed in HEK293T cells are capable of binding to zebrafish IFNF1 or IFNF3 promoters by pull-down assays. HEK293T cells seeded in 10-cm dishes were transfected with 10mg of DrIRF3 or DrIRF7 or empty vector pCS2 as control. At 24 h posttransfection, cells were harvested for sonication in HKMG buffer. A total of 2% of DrIRF3- or DrIRF7-containing cell lysates was loaded as input, and 45% was used for DrIFNF1 or DrIFNF3 promoter binding analysis, as described in (A). (C) Prokaryotically expressed DrIRF3 is capable of binding to zebrafish IFNF1 or IFNF3 promoters by pull-down assays. Zebrafish GST-IRF3 was expressed by a prokaryotic expression system and purified (left panels). A total of 4mg of the purified GST-IRF3 protein was incubated with 1mg of biotin-labeled DrIFNF1 or DrIFNF3 promoter DNA, and the bead-bound DNA-protein complexes were detected by Ab specific to GST tag.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
Binding of DrIRF1/3/7 to IFN promoters as homo- or heteroprotein complexes
To determine whether zebrafish IRF1, IRF3, and IRF7 could coop-eratively regulate IFN responses, co-IP experiments were first per-formed to examine their potential to form homo- or heteroprotein complexes. Two sets of IRF constructs carrying themyctag or the HA tag were used to transfect HEK293T cells. In IRF1-myc–over-expressing cells and IRF1-HA–overIRF1-myc–over-expressing cells, anti-myc Ab– immunoprecipitated protein complex was recognized by anti-HA Ab (Fig. 7A, upper panels). However, an unrelated protein (EGFP-HA) was not immunoprecipitated by IRF1-myc(Fig. 7A, lower panels). These results indicated that DrIRF1 was able to form homodimers. The same was true for DrIRF3 and DrIRF7 (Fig. 7B, 7C). In another experiment in which HEK293T cells were cotransfected with IRF1-HA and IRF3-myc, anti-HA Ab–immunoprecipitated protein complex was detected by anti-IRF3 Ab (Fig. 7D, upper panels) or anti-mycAb–immunoprecipitated protein complex was detected by anti-HA Ab (Fig. 7D, lower panels), suggesting that DrIRF1 was associated with DrIRF3. Similar results showed that DrIRF1 also interacted with DrIRF7 and so did DrIRF3 with DrIRF7 (Fig. 7E, 7F). Considering that in vitro–translated DrIRF3 protein alone failed to bind to IFN promoters, we wondered whether it would do so by virtue of in vitro–translated DrIRF1 or DrIRF7. DNA pull-down assays were performed with DrIRF1/3/7 individually or collectively at a constant amount for each IRF protein (Fig. 7G). Western blot analysis of DrIFNF1 or DrIFNF3 promoter-bound IRF proteins showed that in vitro–translated DrIRF1 and DrIRF7, but not in vitro–translated DrIRF3, were easily detected in single IRF-included reaction setups. Unexpectedly, in vitro–
translated DrIRF3 was still undetectable in the combined reaction setups in which DrIRF1 or DrIRF7 or both were included. Inter-estingly, compared with the reaction setup that included only DrIRF1, two combined reaction setups (DrIRF1+DrIRF7 or DrIRF1+ DrIRF3+DrIRF7) displayed an obviously enhanced binding of DrIRF1, particularly to the DrIFNF3 promoter (Fig. 7G), implying a benefit of DrIRF7 on DrIRF1 binding to fish IFN promoters. To determine whether DrIRF3 would bind to IFN promoters through the formation of homoprotein complexes, we set up reconstitution reaction-based DNA pull-down assays by mixing in vitro–translated DrIRF3 (IRF3-HA) with the lysates of HEK293T cells transfected or not with IRF3-HA. As shown in Fig. 7H, Western blot with anti-HA Ab failed to detect in vitro– translated DrIRF3 in IRF3-alone–included pull-down assays; however, the combination of the same amount of in vitro– translated DrIRF3 and regular HEK293T cell lysates led to detectable binding (Fig. 7H), probably because human IRF3 proteins that were expressed constitutively in HEK293T cells helped in vitro–translated DrIRF3 bind to IFN promoters by forming a homoprotein complex. No binding was detected in the solely HEK293T cell lysate–included reaction because hu-man IRF3 was not recognized by anti-HA Ab; however, a strong binding was seen when HEK293T cells were transfected with IRF3-HA (Fig. 7H). Further assays used a reconstituted DrIRF3 protein mixture consisting of in vitro–translated IRF3 and HEK293T cell lysates in which free-tagged or HA-tagged DrIRF3 had been overexpressed; anti-HA Ab detection of the pull-down proteins showed an obvious binding for the former mixture (over-expression of free-tagged IRF3) and an enhanced binding for the latter (overexpression of IRF3-HA) (Fig. 7I). Collectively, these data suggested that zebrafish IRF1/3/7 bind to fish IFN promoters as homo- and heteroprotein complexes.
FIGURE 5. The binding of zebrafish IRF1, IRF3, and IRF7 to zebrafish IFNF1- and IFNF3-derived IRF-E/ISRE binding sites. DNA pull-down assays were performed by incubating 1mg of biotin-labeled mutant pro-moter DNA, which lacked the individual IRF-E/ISRE motif of DrIFNF1 or DrIFNF3, with in vitro–translated DrIRF1, in vitro–translated DrIRF7, and DrIRF3-overexpressing HEK293T cell lysates. The bead-bound DNA-protein complexes were detected by Ab specific to HA tag.
FIGURE 6. Physical binding of DrIRF1 and DrIRF3 to DrIFNF1 and DrIFNF3 promoters without phosphorylation. DNA pull-down assays were performed as in Fig. 5. For in vitro–translated DrIRF7, the protein was pretreated or not with lamda PPase before pull-down assays. For in vitro–translated DrIRF1 and DrIRF3-overexpressing HEK293T cell lysates, the indicated proteins were treated with lambda PPase before or after incubation with DrIFNF1 or DrIFNF3 promoters or were left treated. The bead-bound DNA-protein complexes were detected by Western blotting. * and **, phosphorylation form of DrIRF1 and DrIRF3.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
Recruitment of DrIRF1/7 to bind to IFN promoters by DrIRF3
We further reconstituted DrIRF3-overexpressed HEK293T cell ly-sates together with in vitro–translated DrIRF7 and/or DrIRF1 proteins to perform DNA pull-down assays. As shown in Fig. 8, each fish IRF protein could bind to zebrafish IFN promoters, with a strong binding for the overexpressed DrIRF3 and a weak one for DrIRF1 or DrIRF7 alone. In comparison with the relatively weak binding by DrIRF1 or DrIRF7 alone, significantly enhanced binding of DrIRF1 or DrIRF7 was observed for the reconstituted fish IRF protein mixtures that were made up of overexpressed DrIRF3 plus DrIRF7, DrIRF1, or both (Fig. 8). Interestingly, this effect was not reciprocal, because this supplementation resulted in an obvious reduction, not an increase, in
DrIRF3 binding. Notably, all reactions contained equal amounts of the same DrIRF proteins and DrIFN promoter DNAs; however, the amounts of IRF proteins that were pulled down by IFN promoter DNAs varied among different experiment combinations, and the amounts of promoter DNA-bound multiple IRF proteins in a given reconstituted reaction setup are not equal to the sum of those from single component–included reaction. Therefore, DrIRF3 might fa-cilitate the binding of DrIRF1 and/or DrIRF7 to fish IFN promoters.
Differential response of DrIFNF1/3 promoters to DrIRF/3/7 individually and collectively
Luciferase assays were used to determine the expression regu-lation of DrIFNF1 and DrIFNF3 by DrIRF1, DrIRF3, and FIGURE 7. The binding of zebrafish IRF1, IRF3, and DrIRF7 to zebrafish IFN promoters through the assembly of homo- or heteroprotein complexes. (A–F) Immunoprecipitation analysis of the association among zebrafish IRF1, IRF3, and IRF7. HEK293T cells seeded in 10-cm dishes were cotransfected with IRF1-mycand IRF1-HA (A), IRF3-mycand IRF3-HA (B), IRF7-mycand IRF7-HA (C), IRF1-HA and IRF3 (D), IRF1-HA and IRF7 (E), or IRF3 and IRF7-HA (F), each with 5mg (upper panels). Cells were transfected with the indicated IRF expression plasmids and an unrelated vector EGFP-HA as control (lower panels). Twenty-four hours later, the transfected cells were immunoprecipitated with anti-mycAb (A–C) or anti-HA Ab (D–F), followed by Western blotting with the indicated Abs. Arrows indicate nonspecific binding. (G–I). Cooperative binding of zebrafish IRF1, IRF3, and IRF7 to zebrafish IFN promoters. DNA pull-down assays were performed as in Fig. 4, by incubation of DrIFNF1 and DrIFNF3 promoter DNA with in vitro–translated DrIRF3, in vitro–translated DrIRF1, and in vitro–translated DrIRF7, individually and collectively (G), or with in vitro–translated DrIRF3 and regular HEK293T cell lysates, individually and collectively, and DrIRF3-transfected HEK293T cell lysates alone as control (H), or with in vitro–translated DrIRF3 and free-tagged DrIRF3-transfected HEK293T cell lysates collectively and HA-tagged DrIRF3-transfected HEK293T cell lysates as control (I). A constant amount of in vitro–translated DrIRF3 was included, and the bead-bound DNA-protein complexes were detected by Ab specific to HA tag.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
DrIRF7. Consistent with our previous results (29, 36), over-expression of each fish IRF resulted in activation of the DrIFNF1 promoter in a dose-dependent fashion (Fig. 9A). Similar acti-vation of the DrIFNF3 promoter was seen in response to DrIRF1 or DrIRF7; however, DrIRF3 failed to activate the DrIFNF3 promoter (Fig. 9C). Among the three IRFs, the best stimulator of the DrIFNF1 promoter was DrIRF3, followed by DrIRF1 and then DrIRF7. For instance, a 19-fold increase in DrIFNF1 pro-moter–driven luciferase activities was detected when 200 ng of DrIRF1 was used, whereas a 160-fold increase was seen with 200 ng of DrIRF3 (Fig. 9A, top panel). In another experiment, a 35-fold increase was observed with DrIRF1 compared with a 18-fold increase with DrIRF7 (Fig. 9A, middle panel). However, the DrIFNF3 promoter did not show such a preference for DrIRF1 and DrIRF7 (Fig. 9C, middle panel).
Considering the varied expression levels of three zebrafish IRF genes in stimulated cells and tissues (Figs. 1, 2), different ratios of any two IRFs were used to evaluate the possible synergistic effects of IRF expression levels on regulating zebrafish IFN expression. The results showed that IFN promoter activities obtained with two IRFs were often higher than those obtained with one (Fig. 9A, 9C). For example, compared with transfection of DrIRF1 alone (200 ng), cotransfection of DrIRF1 (200 ng) and increasing amounts (10, 50, 200 ng) of DrIRF3 resulted in a significant and continuous increase in DrIFNF1 promoter ac-tivities (1.5-, 3.2-, and 5.5-fold higher than control, respec-tively); when compared with transfection of DrIRF3 alone at the corresponding amounts (10, 50, 200 ng), DrIFNF1 promoter
activities were quantified as 5.5-, 1.7-, and 1.2-fold, respectively (Fig. 9A, top panel).
To exclude the interference of additional effects, the coopera-tivity values were also calculated as the ratio of luciferase accoopera-tivity values obtained in the presence of two IRFs/sum of luciferase activity values obtained with each alone (Fig. 9B, 9D). We set the criteria that values around or equal to 1 indicated no cooperative effect, values,1 indicated an inhibitory effect, and values.1 indicated a synergistic activation. Using this method to reassess the activation of the DrIFNF1 promoter by DrIRF1 plus DrIRF3, a gradually increasing synergy was observed along with the in-crease in the relative amounts of DrIRF3, with 1.5–2.2-fold higher level of luciferase activities at DrIRF1/DrIRF3 ratios of 1:4–1:20 (Fig. 9B, top panel).
Interestingly, although DrIRF3 alone could not stimulate the DrIFNF3 promoter, cotransfection of low amounts of DrIRF3 (10 and 50 ng) and 200 ng of DrIRF1 (DrIRF1/DrIRF3 ratio.1:1) resulted in significantly decreased luciferase activities relative to transfection of 200 ng of DrIRF1 alone; conversely, significantly increased luciferase activities were observed when the DrIRF1/ DrIRF3 ratio was #1:1 (Fig. 9C, top panel). These results indi-cated a dual effect of DrIRF3 on DrIRF1-mediated DrIFNF3 expression: a synergistic induction (.1.3–3-fold higher than control) at high DrIRF3 concentrations (DrIRF1/DrIRF3 ratio of 1:4 or 1:20) and an inhibitory effect (4–5-fold lower than control) at low DrIRF3 concentrations (DrIRF1/DrIRF3 ratio of 20:1 or 4:1) (Fig. 9D, top panel). No obvious effect of DrIRF1/7 on DrIFNF1 promoter activation was observed when the DrIRF1/7 FIGURE 8. Enhanced binding of zebrafish IRF1 and
IRF7 to zebrafish IFN promoters by zebrafish IRF3. DNA pull-down assays were performed with the indicated pro-teins as in Fig. 4. A total of 1mg of biotin-labeled pro-moter DNA of DrIFNF1 or DrIFNF3 was incubated with in vitro–translated DrIRF1, in vitro–translated IRF7-HA– and IRF3-HA–transfected HEK293T cell lysates, indi-vidually or collectively. Each IRF was included at a constant amount, and the bead-bound DNA-protein complexes were detected by anti-HA tag Ab.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
ratio was#1:1 because the cooperative value was close to 1, but a slight synergistic induction was detected at higher amounts of DrIRF7 (.1.3-fold higher than control) (Fig. 9A, 9B, middle panels). Similar analyses indicated an obvious synergism on DrIFNF3 at DrIRF1/DrIRF7 ratios of 4:1–1:4, with the highest level (nearly 4-fold higher than control) at a 1:1 ratio (Fig. 9C, 9D, middle panels). DrIRF3 and DrIRF7 displayed robust synergism on stimulating the DrIFNF1 promoter with increasing the relative amounts of DrIRF7 (1.8–3-fold higher than control at DrIRF3/ DrIRF7 ratios of 1:1–1:20) (Fig. 9A, 9B, bottom panels) and dis-play modest synergism on the DrIFNF3 promoter (up to 1.8-fold higher than control) only at low DrIRF3/DrIRF7 ratios (1:4 and 1:20) (Fig. 9C, 9D, bottom panels).
Discussion
In mammals, accumulating data suggested that the type I IFN response is contingent on many issues, including intracellular locations of pathogen, sensing receptors, and IRF members; these issues create a functional link directing a specific signaling pathway in a given cell type (1, 2). Generally, such a link triggers an IRF3/7-dependent IFN response from cytosolic location in
most cell types (8), an IRF7-dependent pathway, and/or an IRF1-dependent pathway from endosomes of plasmacytoid DCs, cDCs, or other immune cells (9, 11–14). In other instances, invading pathogens trigger multiple signaling pathways to activate an IFN response, such as a Listeria monocytogenes mutant that is pri-marily recognized in phagosomes and also in the cytosol of cDCs, thus stimulating IFNb dependently of IRF1 in phagosomes and IRF3/7 in cytosol (11), and L. acidophilus, which upregulates IFN-bin mouse DCs involving IRF1, IRF3, and IRF7 through the MyD88 pathway (21). Therefore, IRF1, IRF3, and IRF7 are three crucial transcription factors and might be simultaneously assem-bled to IFN gene promoters, ensuring full release of IFN response. Like mammalian IRF1, DrIRF1 is constitutively expressed in various tissues and is inducible by IFN and IFN stimuli (Fig. 2). Unlike mammalian IRF3 that is not inducible by IFN stimulation and viral infection (43), and mammalian IRF7, whose expression is limited in lymphoid cell types but is highly induced in various cell types (1), DrIRF3 and DrIRF7 are transcriptionally expressed in all normal tissues tested and significantly induced by SVCV infection (Fig. 2). This disparate property of DrIRF3 and DrIRF7 is also seen in other fish species (28, 44–47), implying the pos-FIGURE 9. Differential effects of zebrafish IRF1, IRF3, and IRF7 on zebrafish IFN promoter activation. (A) Differential response of DrIFNF1 promoter to DrIRF1, DrIRF3, and DrIRF7 by luciferase assays. Crucian carp (C. auratusL.) blastula embryonic cells seeded in 24-well plates were cotransfected with DrIFNF1 promoter–driven luciferase plasmid (0.25mg) in the presence of two zebrafish IRF plasmids at the indicated amounts; 0.025mg pRL-TK was introduced as an internal control. At 48 h posttransfection, cells were harvested for detection of luciferase activity. The data shown are representative of three independent experiments, each performed in triplicate. (B) Cooperativity analysis of zebrafish IRF1/3/7-mediated DrIFNF1 activation based on more than three independent luciferase assays, as in (A). Cooperativity values between any two of zebrafish IRF1/3/7 on DrIFNF1 promoter activation were calculated as the ratio of luciferase activities obtained in the presence of both IRF proteins/sum of luciferase activities obtained when each factor was expressed. The values around or equal to 1 indicated no cooperative effects, values,1 indicated an inhibitory effect, and values.1 indicated a synergistic activation. (C) Differential response of DrIFNF3 promoter to DrIRF1, DrIRF3, and DrIRF7 by luciferase assays. Luciferase assays were performed, as in (A), using a DrIFNF3 promoter-driven luciferase plasmid instead of a DrIFNF1 promoter–driven luciferase plasmid. (D) Cooperativity analysis of zebrafish IRF1/3/7-mediated DrIFNF3 activation, based on more than three independent luciferase assays, as in (C).
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
sibility that fish IRF3 and IRF7 are required for IFN responses in various cell types. Interestingly, very weak transcription of two zebrafish IFN genes, DrIFNF1 and DrIFNF3, is detected in healthy zebrafish tissues in which DrIRF1/3/7 are also constitu-tively abundant (Fig. 2A). Following SVCV infection, DrIFNF1/3 and DrIRF1/3/7 are significantly induced in another set of zebra-fish tissues (Fig. 2B), which correlates with the replication of SVCV (Fig. 2C). SVCV is a negative ssRNA virus (48, 49). Al-though it is uncertain how SVCV infection is recognized in fish cells, the fact that these fish molecules highly respond to SVCV in some common tissues prompted us to hypothesize that DrIRF1/3/7, like their homologs in mammals (11, 21), might cooperatively regulate zebrafish IFN gene expression on unknown occasions. In support of this hypothesis, each of DrIRF1/3/7 interacts with MyD88 (Fig. 3); DrIRF1, together with MyD88, upregulates DrIFNF3 (36), and in Atlantic salmon, IRF3 or IRF7 synergizes with MyD88 to activate the salmon IFN response (35). Notably, zebrafish IFNF1 has two isoforms (40, 41). In the current study, the secreted isoform of DrIFNF1 is induced more significantly than the intracellular one by SVCV infection (Fig. 2B). The dif-ferential induction of both DrIFNF1 isoforms is likely ascribed to their promoter composition. The secreted DrIFNF1 isoform pro-moter has three IRF-E motifs (IRF-E1, IRF-E2, and IRF-E3 in Fig. 5), and the intracellular isoform promoter has only the distant IRF-E1. Our previous results showed that the proximal IRF-E motif primarily contributes to induction of fish IFN gene expres-sion (28), supporting the idea that the secreted one is highly in-ducible by SVCV. Therefore, the secreted DrIFNF1 isoform promoter was used in the subsequent assays.
The data in the current study indicate that DrIRF1, DrIRF3, and DrIRF7 differentially regulate zebrafish IFN expression, likely through assembly of homo- and heteroprotein complexes binding to IRF-E/ISRE sites in IFN promoters. First, DrIRF1, DrIRF3, and DrIRF7 possess highly conserved N-terminal DBD (36), guaran-teeing their binding to the IFN promoter. Second, like human IFNagene promoters whose IRF-E motifs are selectively recog-nized by IRF3 and IRF7 (6, 7), each fish IRF protein binds to cognate IRF-E/ISRE motifs of the DrIFNF1 and DrIFNF3 pro-moters (Fig. 5). Actually, the IRF-E/ISRE motifs are directly re-sponsible for either binding or activation of crucian carp IRF3 and zebrafish IRF1 to fish IFN promoters (28, 36). Third, DrIRF1, DrIRF3, and DrIRF7 are associated with one another in vitro (Fig. 7A–F), and they bind to zebrafish IFN promoters through the formation of homo- and heteroprotein complexes (Figs. 7G, 7H, 8). Finally and importantly, overexpression of DrIRF1, DrIRF3, and DrIRF7 individually and collectively differentially activates two fish promoters depending on their combination and the relative amounts (Fig. 9).
Although eukaryotically expressed DrIRF3 in HEK293T cells or prokaryotically expressed DrIRF3 binds to both zebrafish IFN promoters by pull-down assays, the binding is not detectable with in vitro–translated DrIRF3 protein alone (Fig. 4A). Interestingly, the same phenomenon is seen for in vitro–translated human IRF3 by EMSA assays, and binding to the IFN-bpromoter is observed with a high concentration of recombinant human IRF3 in baculovirus-infected cells (4). However, the failure to bind the IFN promoter is not a result of fish IRF3 protein concentration, because a lower concentration of prokaryotically expressed DrIRF3 than in vitro– translated IRF3 used in our experiments still binds to fish IFN promoters (data not shown). The exact causes of such a phe-nomenon are unknown, but it is possible that a misfolded protein conformation in in vitro–translated IRF3 restricts its binding to IFN promoters. Notably, in vivo assays showed that, in mammals, IRF3 binding to target gene promoters requires the activation of
two coactivators,b-catenin and CBP, by HDAC6 and PKC-b(50). Therefore, it is important to analyze in vivo binding of fish IRF3 to IFN promoters in the future.
The binding features of DrIRF1, DrIRF3, and DrIRF7 seem to determine the differential response of both fish IFN gene pro-moters. For example, DrIRF3 alone activates the DrIFNF1, but not the DrIFNF3, promoter, whereas DrIRF1 and DrIRF7 activate both; they differ in that DrIRF1 has no preference, and DrIRF7 prefers to activate the DrIFNF3 promoter (Fig. 9). Consistently, pull-down assays reveal an unusual binding feature of DrIRF3: it does not have a preference for either of the IRF-E/ISRE motifs within the DrIFNF3 promoters, which is very different from DrIRF3 binding to the DrIFNF1 promoter and DrIRF1/7 binding to DrIFNF1/3 promoters, where a preferred motif is selected (Fig. 5). In mammals, IRF3 possesses a restricted recognizing DNA sequence, but IRF7 has a broader one (51); this selective binding contributes to differential regulation of early-phase IFN genes, including IFNb, and later-phase IFN genes, including most IFNas: the former is primarily controlled by IRF3, and the latter are controlled by IRF7 (6–8, 51–53). Considering that the IRF3/7-dependent IFN response is activated from cytosolic locations in most cell types (8), the similar binding and regulatory properties of DrIRF3 and DrIRF7 support our previous notion that RLR pathway–triggered DrIFNF1 expression is primarily regulated by DrIRF3, thereby resembling mammalian IFNb, and DrIFNF3 is primarily regulated by DrIRF7, thereby resembling mammalian IFNas (29). The binding property of DrIRF1 also correlates with its regulatory potential to activate DrIFNF1 and DrIFNF3 (36).
Interestingly, the magnitude of zebrafish IFN gene expression is dependent on the combinatory binding of DrIRF1/3/7, as well as their relative amounts. In the current study, the cooperativity values were determined as the ratio of luciferase activity values obtained in the presence of both IRFs/the sum of luciferase activity values obtained with each alone, and the values.1 indicate a synergistic activation (Fig. 9B, 9D). Based on this criterion, overexpression of DrIRF1 and DrIRF7 together results in a significantly synergistic activation of the DrIFNF3 promoter and less activation of DrIFNF1 promoter, but at higher amounts of DrIRF7 (Fig. 9, middle panels). This result is consistent with the binding assays showing that DrIRF7 facili-tates an enhanced IFN promoter binding of DrIRF1, particularly to the DrIFNF3 promoter (Fig. 7G). The same is true for cooperative activation of both IFN promoters by DrIRF3 together with DrIRF1 or DrIRF7, which correlates with enhanced promoter binding of DrIRF1 and DrIRF7 as a consequence of DrIRF3 recruitment (Fig. 8). These results indicate a molecular mechanism underlying IRF combination-mediated differential activation of fish IFN pro-moters. Generally, the best cooperation occurs when DrIRF3 or DrIRF7 is present at higher amounts, likely due to the fact that IRF3 and IRF7 are two essential transcription factors for distinct IFN expression in most cell types (8, 29), with the exception that DrIRF1/7-dependent DrIFNF3 expression is most significantly induced at equal amounts of DrIRF1 and DrIRF7. The IRF concentration-dependent IFN gene-expression patterns might re-flect an important physiological significance. In healthy or in-fected zebrafish tissues, DrIRF1/3/7 display various constitutive or inducible expression levels; therefore, when these factors are activated simultaneously in a given cell type, even downstream of different pattern recognition receptor–mediated signaling, they might work together to fine-tune the activation of DrIFNF1 and DrIFNF3 genes, depending on their relative expression levels.
Although DrIRF3 alone does not activate the DrIFNF3 pro-moter, it still regulates DrIRF1- or DrIRF7-mediated DrIFNF3 gene expression. Strikingly, DrIRF3 facilitates DrIRF7-mediated DrIFNF3 activation only at high concentrations (Fig. 9C, 9D,
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
motifs within IFNa and IFNb gene promoters (51); however, this factor activates the expression of early-phase IFN genes like IFNbbut has no effect on most IFNagene expression (6, 7, 51). Similar to DrIRF3, human IRF3 selectively exerts a similarly dual effect on IRF7-mediated IFNagene expression (7) and an inhibitory effect on IRF1-mediated IFNa4 gene expression (5). Obviously, the mechanism of IRF-mediated IFN expression in humans is not identical to that in zebrafish, and we cannot ex-clude the possibility that there are dual effects of zebrafish IRF3 on IRF7-mediated IFN responses, because two other zebrafish IFN genes (DrIFNF2/4) are not included in the current study. Despite these differences, these findings suggest that zebrafish IFN genes are regulated in a way similar to mammalian type I IFNs. It is well known that fish IFN genes and mammalian type I IFN genes have evolved independently, indicating that extant fish IFN genes do not display a one-to-one orthologous rela-tionship with mammalian IFNa/bgenes (22, 25). On the con-trary, IRF family genes seem to pre-exist in the common ancestor of both fish and mammals, displaying a clearly orthologous relationship between fish and mammals (3, 28, 45, 54). Based on these findings, it is likely that fish and mammals might adopt the pre-existing transcription factors IRF1/3/7 independently to develop a similar regulatory mechanism, whereby the IFN re-sponse is intricately and adequately modulated depending on the fluctuating expression levels of IRF1/3/7 in a given tissue or cell type.
In mammals, IRF3 phosphorylation leads to its homo or het-erodimerization, nuclear translocation, and association withb-catenin and CBP/p300, and this is the transcriptional complex for IFNb expression (4, 7, 51, 55). Consistently, fish IRF3 and IRF7 lo-calize in cytoplasm of unstimulated cells (28, 44, 56), and undergo a cytoplasmic-to-nuclear translocation following poly(I:C) treat-ment or virus infection, likely through phosphorylation (28, 56). In the current study, physical binding of DrIRF3 and DrIRF1 to IFN promoters is independent of phosphorylation (Fig. 6). Therefore, it is likely that in vivo phosphorylation activation is required for translocation of IRF3 and IRF7 across the nuclear membranes into the nucleus but not for physical binding to the IFN promoters. Unlike IRF3 and IRF7, fish and mammalian IRF1s reside in the nucleus (31, 36). Although phosphorylation of DrIRF1 is observed, its physiological relevance remains to be determined. Interestingly, a recent study showed that phosphory-lation of human IRF1 in primary CD4+T cells blocks induction of IFN-bfollowing treatment with poly(I:C) (57).
Collectively, the data from the present study suggest that there is similar IRF1/3/7-regulated IFN activation between fish and mammals. Like human IRF3 (5, 7), DrIRF3 functions as a positive regulator and as a negative regulator of different IFN genes, depending on different amounts of DrIRF1/3/7. These results in-dicate a similar regulatory mechanism of IRF-mediated IFN re-sponses in fish and mammals. In contrast with mammalian IFNa/b (8, 52, 53), DrIFNF1 and DrIFNF3 appear not to be induced as early- or late-phase IFNs (29), and fish IRF3 is an IFN-inducible transcription factor and lacks an inhibitory domain (28). Therefore, even if fish IRF1/3/7 function as partners to coordinately regulate distinct fish IFN activation, ensuring an efficient, but not excessive,
1. Tamura, T., H. Yanai, D. Savitsky, and T. Taniguchi. 2008. The IRF family tran-scription factors in immunity and oncogenesis.Annu. Rev. Immunol.26: 535–584. 2. G€urtler, C., and A. G. Bowie. 2013. Innate immune detection of microbial
nucleic acids.Trends Microbiol.21: 413–420.
3. Stein, C., M. Caccamo, G. Laird, and M. Leptin. 2007. Conservation and di-vergence of gene families encoding components of innate immune response systems in zebrafish.Genome Biol.8: R251.
4. Wathelet, M. G., C. H. Lin, B. S. Parekh, L. V. Ronco, P. M. Howley, and T. Maniatis. 1998. Virus infection induces the assembly of coordinately activated transcription factors on the IFN-beta enhancer in vivo.Mol. Cell1: 507–518. 5. Schafer, S. L., R. Lin, P. A. Moore, J. Hiscott, and P. M. Pitha. 1998. Regulation
of type I interferon gene expression by interferon regulatory factor-3.J. Biol. Chem.273: 2714–2720.
6. Civas, A., P. Ge´nin, P. Morin, R. Lin, and J. Hiscott. 2006. Promoter organization of the interferon-A genes differentially affects virus-induced expression and responsiveness to TBK1 and IKKepsilon.J. Biol. Chem.281: 4856–4866. 7. Ge´nin, P., R. Lin, J. Hiscott, and A. Civas. 2009. Differential regulation of
hu-man interferon A gene expression by interferon regulatory factors 3 and 7.Mol. Cell. Biol.29: 3435–3450.
8. Honda, K., H. Yanai, H. Negishi, M. Asagiri, M. Sato, T. Mizutani, N. Shimada, Y. Ohba, A. Takaoka, N. Yoshida, and T. Taniguchi. 2005. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Na-ture434: 772–777.
9. Kawai, T., S. Sato, K. J. Ishii, C. Coban, H. Hemmi, M. Yamamoto, K. Terai, M. Matsuda, J. Inoue, S. Uematsu, et al. 2004. Interferon-alpha induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6.Nat. Immunol.5: 1061–1068.
10. Barbalat, R., L. Lau, R. M. Locksley, and G. M. Barton. 2009. Toll-like receptor 2 on inflammatory monocytes induces type I interferon in response to viral but not bacterial ligands.Nat. Immunol.10: 1200–1207.
11. Mancuso, G., M. Gambuzza, A. Midiri, C. Biondo, S. Papasergi, S. Akira, G. Teti, and C. Beninati. 2009. Bacterial recognition by TLR7 in the lysosomes of conventional dendritic cells.Nat. Immunol.10: 587–594.
12. Dietrich, N., S. Lienenklaus, S. Weiss, and N. O. Gekara. 2010. Murine toll-like receptor 2 activation induces type I interferon responses from endolysosomal compartments.PLoS One5: e10250.
13. Negishi, H., Y. Fujita, H. Yanai, S. Sakaguchi, X. Ouyang, M. Shinohara, H. Takayanagi, Y. Ohba, T. Taniguchi, and K. Honda. 2006. Evidence for li-censing of IFN-gamma-induced IFN regulatory factor 1 transcription factor by MyD88 in Toll-like receptor-dependent gene induction program. Proc. Natl. Acad. Sci. USA103: 15136–15141.
14. Schmitz, F., A. Heit, S. Guggemoos, A. Krug, J. Mages, M. Schiemann, H. Adler, I. Drexler, T. Haas, R. Lang, and H. Wagner. 2007. Interferon-regulatory-factor 1 controls Toll-like receptor 9-mediated IFN-beta production in myeloid dendritic cells.Eur. J. Immunol.37: 315–327.
15. Oganesyan, G., S. K. Saha, E. M. Pietras, B. Guo, A. K. Miyahira, B. Zarnegar, and G. Cheng. 2008. IRF3-dependent type I interferon response in B cells reg-ulates CpG-mediated antibody production.J. Biol. Chem.283: 802–808. 16. Schoggins, J. W., S. J. Wilson, M. Panis, M. Y. Murphy, C. T. Jones, P. Bieniasz,
and C. M. Rice. 2011. A diverse range of gene products are effectors of the type I interferon antiviral response.Nature472: 481–485.
17. Schoggins, J. W., D. A. MacDuff, N. Imanaka, M. D. Gainey, B. Shrestha, J. L. Eitson, K. B. Mar, R. B. Richardson, A. V. Ratushny, V. Litvak, et al. 2014. Pan-viral specificity of IFN-induced genes reveals new roles for cGAS in innate immunity.Nature505: 691–695.
18. Tailor, P., T. Tamura, H. J. Kong, T. Kubota, M. Kubota, P. Borghi, L. Gabriele, and K. Ozato. 2007. The feedback phase of type I interferon induction in den-dritic cells requires interferon regulatory factor 8.Immunity27: 228–239. 19. Li, P., J. J. Wong, C. Sum, W. X. Sin, K. Q. Ng, M. B. Koh, and K. C. Chin.
2011. IRF8 and IRF3 cooperatively regulate rapid interferon-b induction in human blood monocytes.Blood117: 2847–2854.
20. Lazear, H. M., A. Lancaster, C. Wilkins, M. S. Suthar, A. Huang, S. C. Vick, L. Clepper, L. Thackray, M. M. Brassil, H. W. Virgin, et al. 2013. IRF-3, IRF-5, and IRF-7 coordinately regulate the type I IFN response in myeloid dendritic cells downstream of MAVS signaling. [Published erratum appears in 2013PLoS Pathog. 9.]PLoS Pathog.9: e1003118.
21. Weiss, G., K. Maaetoft-Udsen, S. A. Stifter, P. Hertzog, S. Goriely, A. R. Thomsen, S. R. Paludan, and H. Frøkiær. 2012. MyD88 drives the IFN-b response toLactobacillus acidophilusin dendritic cells through a mechanism involving IRF1, IRF3, and IRF7.J. Immunol.189: 2860–2868.
22. Zhang, Y. B., and J. F. Gui. 2012. Molecular regulation of interferon antiviral response in fish.Dev. Comp. Immunol.38: 193–202.
23. Zou, J., and C. J. Secombes. 2011. Teleost fish interferons and their role in immunity.Dev. Comp. Immunol.35: 1376–1387.
24. Gui, J. F., and Z. Y. Zhu. 2012. Molecular basis and genetic improvement of economically important traits in aquaculture animals. Chin. Sci. Bull.57: 1751–1760.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/
25. Zou, J., C. Tafalla, J. Truckle, and C. J. Secombes. 2007. Identification of a second group of type I IFNs in fish sheds light on IFN evolution in vertebrates.J. Immunol.179: 3859–3871.
26. Aggad, D., M. Mazel, P. Boudinot, K. E. Mogensen, O. J. Hamming, R. Hartmann, S. Kotenko, P. Herbomel, G. Lutfalla, and J. P. Levraud. 2009. The two groups of zebrafish virus-induced interferons signal via distinct receptors with specific and shared chains.J. Immunol.183: 3924–3931.
27. Yu, F.-F., Y.-B. Zhang, T.-K. Liu, Y. Liu, F. Sun, J. Jiang, and J.-F. Gui. 2010. Fish virus-induced interferon exerts antiviral function through Stat1 pathway. Mol. Immunol.47: 2330–2341.
28. Sun, F., Y. B. Zhang, T. K. Liu, L. Gan, F. F. Yu, Y. Liu, and J. F. Gui. 2010. Characterization of fish IRF3 as an IFN-inducible protein reveals evolving regulation of IFN response in vertebrates.J. Immunol.185: 7573–7582. 29. Sun, F., Y. B. Zhang, T. K. Liu, J. Shi, B. Wang, and J. F. Gui. 2011. Fish MITA
serves as a mediator for distinct fish IFN gene activation dependent on IRF3 or IRF7.J. Immunol.187: 2531–2539.
30. Shi, J., Y. B. Zhang, T. K. Liu, F. Sun, and J. F. Gui. 2012. Subcellular locali-zation and functional characterilocali-zation of a fish IRF9 from crucian carpCarassius auratus. Fish Shellfish Immunol.33: 258–266.
31. Shi, Y., Y. B. Zhang, Z. Zhao, J. Jiang, Q. Y. Zhang, and J. F. Gui. 2008. Mo-lecular characterization and subcellular localization ofCarassius auratus inter-feron regulatory factor-1.Dev. Comp. Immunol.32: 134–146.
32. Li, S., L. F. Lu, H. Feng, N. Wu, D. D. Chen, Y. B. Zhang, J. F. Gui, P. Nie, and Y. A. Zhang. 2014. IFN regulatory factor 10 is a negative regulator of the IFN responses in fish.J. Immunol.193: 1100–1109.
33. Caipang, C. M., I. Hirono, and T. Aoki. 2005. Induction of antiviral state in fish cells by Japanese flounder,Paralichthys olivaceus, interferon regulatory factor-1. Fish Shellfish Immunol.19: 79–91.
34. Caipang, C. M., I. Hirono, and T. Aoki. 2009. Modulation of the early immune response against viruses by a teleostean interferon regulatory factor-1 (IRF-1). Comp. Biochem. Physiol. A Mol. Integr. Physiol.152: 440–446.
35. Iliev, D. B., M. Sobhkhez, K. Fremmerlid, and J. B. Jørgensen. 2011. MyD88 interacts with interferon regulatory factor (IRF) 3 and IRF7 in Atlantic salmon (Salmo salar): transgenic SsMyD88 modulates the IRF-induced type I inter-feron response and accumulates in aggresomes.J. Biol. Chem.286: 42715– 42724.
36. Feng, H., Y. B. Zhang, Q. M. Zhang, Z. Li, Q. Y. Zhang, and J. F. Gui. 2015. Zebrafish IRF1 regulates IFN antiviral response through binding to IFNf1 and IFNf3 promoters downstream of MyD88 signaling.J. Immunol.194: 1225–1238. 37. Liu, T. K., Y. B. Zhang, Y. Liu, F. Sun, and J. F. Gui. 2011. Cooperative roles of fish protein kinase containing Z-DNA binding domains and double-stranded RNA-dependent protein kinase in interferon-mediated antiviral response. J. Virol.85: 12769–12780.
38. Liu, Y., Y. B. Zhang, T. K. Liu, and J. F. Gui. 2013. Lineage-specific expansion of IFIT gene family: an insight into coevolution with IFN gene family.PLoS One 8: e66859.
39. Lu, D., C. Han, and T. Wu. 2012. Microsomal prostaglandin E synthase-1 pro-motes hepatocarcinogenesis through activation of a novel EGR1/b-catenin sig-naling axis.Oncogene31: 842–857.
40. Levraud, J. P., P. Boudinot, I. Colin, A. Benmansour, N. Peyrieras, P. Herbomel, and G. Lutfalla. 2007. Identification of the zebrafish IFN receptor: implications for the origin of the vertebrate IFN system.J. Immunol.178: 4385–4394. 41. Briolat, V., L. Jouneau, R. Carvalho, N. Palha, C. Langevin, P. Herbomel,
O. Schwartz, H. P. Spaink, J. P. Levraud, and P. Boudinot. 2014. Contrasted innate responses to two viruses in zebrafish: insights into the ancestral repertoire of vertebrate IFN-stimulated genes.J. Immunol.192: 4328–4341.
42. Siednienko, J., T. Gajanayake, K. A. Fitzgerald, P. Moynagh, and S. M. Miggin. 2011. Absence of MyD88 results in enhanced TLR3-dependent phosphorylation of IRF3 and increased IFN-b and RANTES production.J. Immunol. 186: 2514–2522.
43. Au, W. C., P. A. Moore, W. Lowther, Y. T. Juang, and P. M. Pitha. 1995. Identification of a member of the interferon regulatory factor family that binds to the interferon-stimulated response element and activates expression of interferon-induced genes.Proc. Natl. Acad. Sci. USA92: 11657–11661. 44. Holland, J. W., S. Bird, B. Williamson, C. Woudstra, A. Mustafa, T. Wang,
J. Zou, S. C. Blaney, B. Collet, and C. J. Secombes. 2008. Molecular charac-terization of IRF3 and IRF7 in rainbow trout,Oncorhynchus mykiss: functional analysis and transcriptional modulation.Mol. Immunol.46: 269–285. 45. Zhang, Y. B., C. Y. Hu, J. Zhang, G. P. Huang, L. H. Wei, Q. Y. Zhang, and
J. F. Gui. 2003. Molecular cloning and characterization of crucian carp (Carassius auratusL.) interferon regulatory factor 7.Fish Shellfish Immunol. 15: 453–466.
46. Bergan, V., Ø. Kileng, B. Sun, and B. Robertsen. 2010. Regulation and func-tion of interferon regulatory factors of Atlantic salmon.Mol. Immunol.47: 2005–2014.
47. Zhang, J., Y. X. Li, and Y. H. Hu. 2015. Molecular characterization and ex-pression analysis of eleven interferon regulatory factors in half-smooth tongue sole,Cynoglossus semilaevis.Fish Shellfish Immunol.44: 272–282.
48. Hoffmann, B., H. Sch€utze, and T. C. Mettenleiter. 2002. Determination of the complete genomic sequence and analysis of the gene products of the virus of spring viremia of Carp, a fish rhabdovirus.Virus Res.84: 89–100.
49. Zhang, Q., and J. F. Gui. 2015. Virus genomes and virus-host interactions in aquaculture animals.Sci. China Life Sci.58: 156–169.
50. Chattopadhyay, S., V. Fensterl, Y. Zhang, M. Veleeparambil, J. L. Wetzel, and G. C. Sen. 2013. Inhibition of viral pathogenesis and promotion of the septic shock response to bacterial infection by IRF-3 are regulated by the acetylation and phosphorylation of its coactivators.MBio4: e00636-12.
51. Lin, R., P. Ge´nin, Y. Mamane, and J. Hiscott. 2000. Selective DNA binding and association with the CREB binding protein coactivator contribute to differential activation of alpha/beta interferon genes by interferon regulatory factors 3 and 7. Mol. Cell. Biol.20: 6342–6353.
52. Marie´, I., J. E. Durbin, and D. E. Levy. 1998. Differential viral induction of distinct interferon-alpha genes by positive feedback through interferon regula-tory factor-7.EMBO J.17: 6660–6669.
53. Sato, M., H. Suemori, N. Hata, M. Asagiri, K. Ogasawara, K. Nakao, T. Nakaya, M. Katsuki, S. Noguchi, N. Tanaka, and T. Taniguchi. 2000. Distinct and es-sential roles of transcription factors IRF-3 and IRF-7 in response to viruses for IFN-alpha/beta gene induction.Immunity13: 539–548.
54. Huang, B., Z. T. Qi, Z. Xu, and P. Nie. 2010. Global characterization of inter-feron regulatory factor (IRF) genes in vertebrates: glimpse of the diversification in evolution.BMC Immunol.11: 22.
55. Lin, R., Y. Mamane, and J. Hiscott. 1999. Structural and functional analysis of interferon regulatory factor 3: localization of the transactivation and auto-inhibitory domains.Mol. Cell. Biol.19: 2465–2474.
56. Cui, H., Y. Yan, J. Wei, X. Huang, Y. Huang, Z. Ouyang, and Q. Qin. 2011. Identification and functional characterization of an interferon regulatory factor 7-like (IRF7-like) gene from orange-spotted grouper,Epinephelus coioides.Dev. Comp. Immunol.35: 672–684.
57. Sgarbanti, M., G. Marsili, A. L. Remoli, E. Stellacci, A. Mai, D. Rotili, E. Perrotti, C. Acchioni, R. Orsatti, N. Iraci, et al. 2014. IkB kinaseεtargets interferon regulatory factor 1 in activated T lymphocytes.Mol. Cell. Biol.34: 1054–1065.
at Univ of NC at Chapel Hill Health Sciences Library on August 16, 2019
http://www.jimmunol.org/