TABLE OF CONTENTS: VOL. 49, NO. 3, May 1996 ARTICLES
Animal Ecology
194 An investigation on fire effects within xeric sage grouse brood habitat by Richard A. Fischer, Kerry P Reese, and John W. Connelly
199 Shrub-grassland small mammal and vegetation responses to rest from grazing by Steven S. Rosenstock
Plant Animal 204
209 215 223
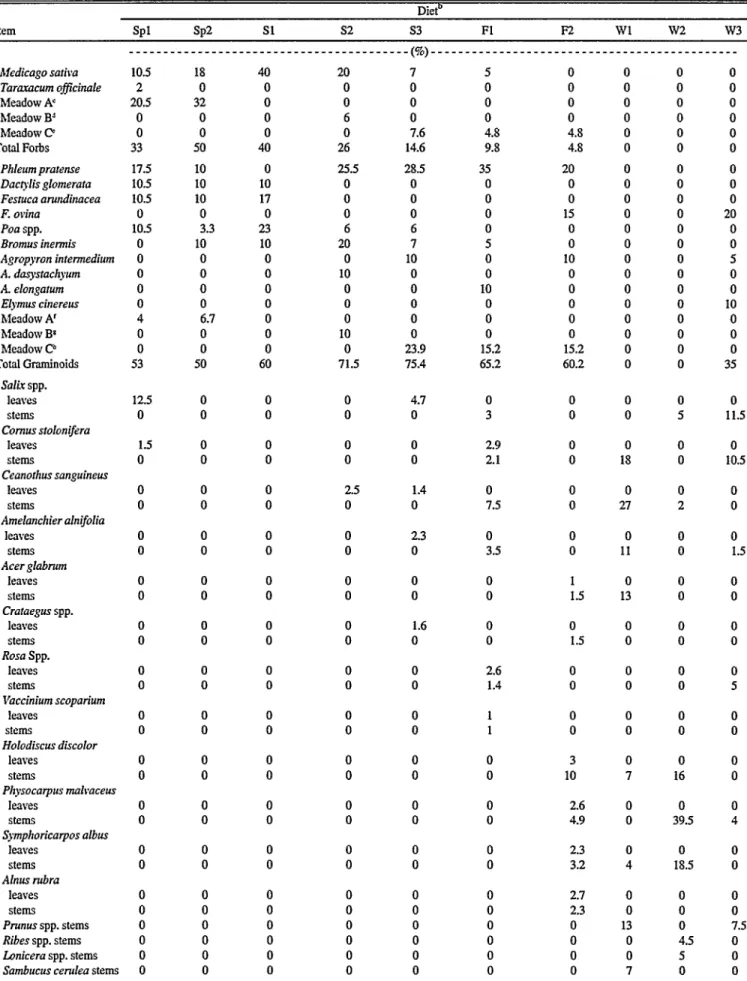
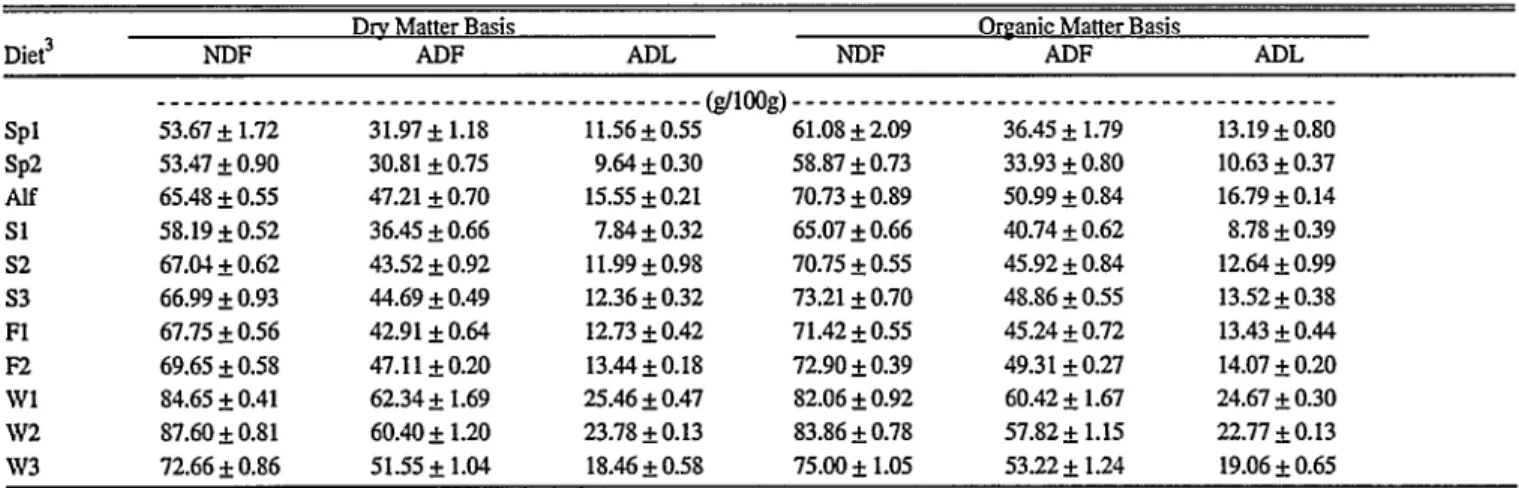
Season and sex influences on botanical composition of cattle diets in south- ern New Mexico by Ayed G. Mohammad, CA. Ferrando, Leigh W. Murray, Rex D. Pieper, and Joe D. Wallace
Effect of breed on botanical composition of cattle diets on Chihuahuan desert range by J.A. Winder, D.A. Walker, and C.C. Bailey
Monitoring mule deer quaility and intake with fecal indices by Thomas P Hodgman, Bruce B. Davitt, and Jack R. Nelson
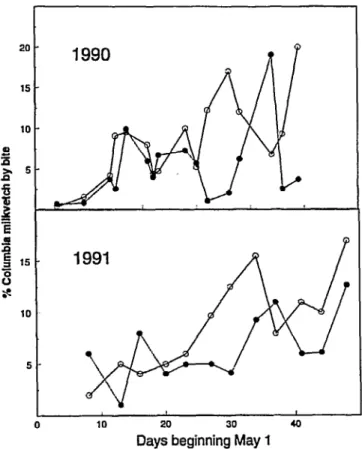
Seasonal grazing of Columbia milkvetch by cattle on rangelands in British Columbia by Walter Majak, Leo Stroesser, John W. Hall, Dee A. Quinton, and Harriet E. Douwes
Plant Ecology
228 Fringed sagebrush response to sward disturbances: Seeding dynamics and plant growth by Yuguang Bai and J.T. Romo
234 Mineral dynamics in forages of the Northern Great Plains by E.E. Grings, M.R. Haferkamp, R.K. Heitschmidt, and M.G. Karl
241 Emergence date effects on resource partitioning between diffuse knapweed seedlings by Roger L. Sheley and Larry L. Larson
Plant Physiology
245 Environmental effects on picloram uptake and ethylene production by broom snakeweed by Tracy M. Sterling, Norman K. Lownds, and Leigh W.
Murray
Published bimonthly--January, March, May,
July, September, November 251 Soil water effects on growth and nutrition in upland pastures by A.E.S.
Macklon, L.A. Mackie-Dawson, C.A. Shand, and A. Sip Copyright 1996 by the Society for Range Man-
agement Grazing Management
INDIVIDUAL SUBSCRIPTION is by member- 257 Influence of grazing management on intake and composition of cattle diets ship in the Society for Range Management. by David J. Hirschfeld, Donald R. Kirby, Joel S. Caton, Steven S. Silcox, and K.C.
Olson LIBRARY or other INSTITUTIONAL SUB-
SCRIPTIONS on a calendar year basis are 264 A theoretical basis for study and management of trampling by cattle by Fred
$95.00 for the United States postpaid and S. Guthery and Ralph L. Bingham
$112.00 for other countries, postpaid. Payment
from outside the United States should be remit- Economics ted in Us do’lars by intemational money Order
or draft on a New York bank. 270 Public land policy and the market value of New Mexico ranches, 1979-94 by L. Allen Tore11 and Marc E. Kin&d.
BUSINESS CORRESPONDENCE, concerning
subscriptions, advertising, reprints, back Measurement/Sampling
issues, and related matters, should be 277 Evaluation of a site conservation rating system in southeastern Arizona by addressed to the Managing Editor, 1639 York
Street, Denver, Colorado 60206. Susan A. Watters, Mark A. Weltz, and E. Lamar Smith BOOK REVIEWS
EDiToRtAL CoRRESPoNDENCE* conceming
manuscripts or other editorial matters, should 285 Landscape Forestry by Stephen G. Boyce; Sustainable Small-scale Ruminant be addressed to the Editor, Gary Frasier, 7620 Production in Semi-arid and Sub-humid Tropical Areas edited by Klaus Becker,
Peter Lawrence and Rob Orskov.
Stag Hollow Road, Loveland, Colorado 80536.
Page proofs should returned to the Production Editor, 1839 York Street, Denver, Colorado 80206.
INSTRUCTIONS FOR AUTHORS appear on the inside back cover of most issues. T/-/E JOURNAL OF RANGE MANAGEMENT (ISSN 0022409X) is published bimonthly for $56.00 per year by the Society for Range Management, 1839 York Street, Denver, Colorado 80206. SECOND CLASS POSTAGE paid at Denver, Colorado, and additional offices.
POSTMASTER: Return entire journal with address change -RETURN POSTAGE GUARANTEED to Society for Range Management, 1839 York Street, Denver, Colorado 80206.
PRINTED IN USA Managing Editor
CHARLES B. (BUD) RUMBURG 1839 York Street
Denver, Colorado 80208 Editor
GARY FRASIER 7820 Stag Hollow Road Loveland, Colorado 80538 Production Editor PATTY RICH
Society for Range Management 1839 York Street
Denver, Colorado 80206 Book Review Editor
DAVID L. SCARNECCHIA Department of Natural Resource Sciences Washington State University Pullman, Washington 991646410
Associate Editors DON ADAMS
West Central Res. & Ext. Center Route 4, Box 46A
North Platte, Nebraska 69101 TERRANCE G. BIDWELL
Department of Agronomy Oklahoma State University Agric. Hall, Room 373
Stillwater, Oklahoma 74076 MARSHALL R. HAFERKAMP
USDAlARS
Oklahoma State University Agric. Hall, Room 373 Stillwater, Oklahoma 74078 STUART P. HARDEGREE
USDA/ARS
800 Park Blvd., Plaze IV, #IO5 Boise, Idaho 83712.
RICHARD E. JOOST Plant Science Unit University of Missouri 214 Waters Hall Columbia, Missouri 65211
F. TED MCCOLLUM F. MONTE ROQUET-TE
Oklahoma State University Texas A&M University 212G Animal Science Bldg. Agric. Res. & Ext. Center Stillwater, Oklahoma 74078-0425 Overton, Texas 75684-0209 DAVID MICHALK
Agdc. Res. & Veterinary Centre Forest Road
Orange, NSW 2800 AUSTRALIA
SHERM SWANSON
Renewable Resources Center University of Nevada 100 Valley Road Rena, Nevada 89512 M. KEITH OWENS
Texas A&M University Research Center 1619 Garner Field Road Uvalde, Texas 78601
JOHN TANAKA Agr. & Res. Econ.
Oregon State University 213 Ballard Ext. Hall Corvallis, Oregon 97331 JEFF POWELL
Dept. of Range Management University of Wyoming University Station, Box 3354 Laramie, Wyoming 82071
JOHN WALKER US Sheep Exp. Station HC 62, Box 2010 Dubois, Idaho 64323
MIKE RALPHS 1150 East 1400 North Logan, UT 84321
WALTER WILLMS
Agric. Canada Res. Center Lethbndge, AB Tl J 481 CANADA
- -.
President JOHN R. HUNTBR
Texas Tech University Range I% Wildlife Management Lubbock. Texas 79409 1st Vice-President JOHN C. BUCKHOUSE
Oregon State University
Department of Rangeland Resources Corvallis. Oregon 9733 1
2nd Vice-President E. LARhfAR ShlITH, JR.
Div. of Range hlanagement, BSE 325 University of Arizona
Tucson, Arizona 85721 Esecutive Vice-President CHARLES B. RUhlBURG
Society for Range hkmagement IS39 York Street
Dsnver, Colorado SO206 (303) 355-7070 Diictors 1994-1996
LINDA H. HARDESTY Bos 2023
College Station
Pullman, Washington 99165 IAhlES T. O’ROURKE
HC 75. Box 20
Chndron, Nebraska 69337 MS-1997
!.T. “TOM” BARTLETT Colondo State University Department of Range Science Fort Collins, Colorado SO523
&RGUERITE B. “hIEG” ShllTH Box 7
Glen, Montana 59732
!X’6-1998
:ODNEY HEITSCHMIDT t. Keoph LARRL :t 1, Box 2021
files City, Montana 59301 :ONALD E. SOSEBEE 902 Geneva ubbuck, Texas 79413
THE SOCIETY FOR RANGE MANAGEMENT, founded in 1948 as he American Society of Range Management, is a nonprofit association incorporated under the laws If the State of Wyoming. It is recognized exempt from Federal income tax, as a scientific and educa- ional organization, under the provisions of Section 501(c)(3) of the Internal Revenue Code, and also .s classed as a public foundation as described in Section 509(a)(2) of tbe Code. The name of the Society was changed in 1971 by amendment of the Articles of Incorporation.
The objectives for which the corporation is established are:
-to properly take care of the basic rangeland resources of soil. plants and water;
-to develop an understanding of range ecosystems and of the principles applicable to the management of range resources;
-to assist all who work with range resources to keep abreast of newfindings and techniques in the science and art of range management:
-to improve the effectiveness of range management to obtain from range resources the products and values necessary for man’s welfare;
-to create a public appreciation of the economic and social benefits to be obtainedfrom the range environment:
-to promote professional development of its members.
Membership in tbe Society for Range Management is open to anyone engaged in or interested in any aspect of the study, management, or use of rangelands. Please contact the Executive Vice- President for details.
The Journal of Range Management is a publication of the Society for Range Management. It serves as a forum for the presentation and discussion of facts, ideas, and philosophies pertaining to the study, management, and use of rangelands and their several resources. Accordingly, all material published herein is signed and reflects the individual views of the authors and is not necessari- ly au official position of the Society. Manuscripts from anyone-nonmembers as well as members-are welcome and will be given every consideration by the editors. Editorial comments by au individual is also welcome and, subject to acceptance by the editor, will be published as a “Viewpoint.”
In Cooperation With: Some of the articles appearing in The Journal of Range Management (JRM) are presented in cooperation with The American Forage and Grassland Council (AFGC). This coopera- tion consists of JRM acceptance of professional papers in forage grazing management and related sub- ject areas from AFGC members and the appointment of two AFGC affiliated associate editors
to JRM’s Editorial Staff. The American Forage and Grassland Council OffIces: 210 W. 8th Street, #A, P.0. Box 94, Georgetown, Texas 78626; C. Pat Bagley. President; Dana Tucker, Executive Secretary.
Contribution POliCy: The Society for Range Management may accept donations of real andlor personal property subject to limitations set forth by State and Federal law. All donations shall be subject to management by the Executive Vice President as directed by the Board of Directors and their discretion in establishing and maintaining trusts, memorials, scholarships or other types of funds.
Individual endowments for designated purposes can be established according to Society policies. Gifts, bequests, legacies, devises, or donations not intended for establishing designated endowments will be deposited into the SRM Endowment Fund. Donations or requests for further information on Society poli- cies can be directed to the Society for Range Management, Executive Vice-President, 1839 York Street, Denver, CO 80206. We recommend that donors consult Tax Advisors in regard to any tax consideration that may result from any donation.
he term of office of all elected officers and irectors begins in February of each year during le Society’s annual meeting.
J. Range Manage.
49:194-198
An investigation on fire effects within xeric sage grouse brood habitat
RICEIARD A. FISCHER, KERRY P. REESE, AND JOFIN W. CONNELLY
Authors are former research assistant and associate professor, Department of Fish and Wildlife Resources, University of Idaho, Moscow, Ida., 83844; and Principal Wildlife Biologist, Idaho Depamnent of Fish and Game, 1345 Barton Rd., Pocatello, Ida., 83205
Abstract
We investigated the short-term iniluence of tire on xeric sage grouse (Centrocercus urophasianus) brood habitat in southeast- ern Idaho from 1990-92. A prescribed fire in 1989 removed Wyoming big sagebrush (Artemisia tridentata wyomingensis Nutt.)/threetip sagebrush (A. trijlartzb Rydb.) canopy cover from approximately 57% of a 5,800-ha area, potentially intluencing brood-rearing habitat. Although the fire created a mosaic of sagebrush areas interspersed with open areas having abundant grasses and forbs, the relative abundance of males, females, and broods on survey routes in burned and unburned habitat were similar. Cover of forbs important in sage grouse summer diets was similar in burned and unburned habitat. However, the abun- dan;e of Hymenoptera, an insect Order important in sage grouse diets, was significantly lower in burned habitat the second and third years postburn. Our research did not support the con- tention that fue may enhance sage grouse brood-rearing habitat.
Key Words: big sagebrush, Centrocercus urophasiunus, insects.
Knowledge of the relationship between bid species and their food resources is a critical component of avian ecology (Raley and Anderson 1990). Invertebrates, mainly insects, are consumed in large quantities and provide essential protein in the diets of most young birds, such as waterfowl (Swanson et al. 1985), passerines (Lack 1954), and galliformes (Kobriger 1965, Klebenow and Gray 1968, King 1969). Wyoming big sagebrush (Artemisia tridentata wyomingensis Nutt.) communities have a diverse insect fauna (Horning and Barr 1970, Stafford 1983) that provides an important food source for many vertebrates (Gleason 1978, Halford 1981). High protein foods such as forbs and insects in these communities are important in sage grouse chick diets
Funding was provided by the Idaho Department of Fish and Game (IDFG) through the University of Idaho. Logistical support was provided by the U.S.
Bureau of Land hlanagment. This is 3 contribution from Idaho Federal Aid in Wildlife Restoration Project W-160-R and Contribution 763 of the University of Idaho College of Forestry, Wildlife and Range Experiment Station.
We gratefully acknowledge R. Bradish, A.W. Farris. M.B. Fischer, J.F.
Kennedy, M.A. Sands, and J.W. Snyder, for their assistance in data collection. W.
Newton, R.K. Stehthorst, and C. Williams provided statistical advice.
Present address of senior author: U.S. Army Corps of Engineer Waterways Experiment Station, Attrc CEWES-EN-S, 3909 Halls Ferry Rd., Vicksburg, Miss.
39180.
hkmscript accepted 29 July 1995.
194
(Klebenow and Gray 1968, Peterson 1970), especially during the first 3 weeks after hatching (Johnson and Boyce 1990).
Sagebrush removal on western rangelands is a common prac- tice for increasing forage availability for livestock. Currently, prescribed fire is a popular and cost-effective method of sage- brush removal (Frandsen 1985, Bunting et al. 1987). However, little is known about how various methods of habitat manipula- tion affect important foods of sage grouse chicks. Sagebrush treatment projects (e.g., herbicide application) that reduce the abundance and diversity of forbs are detrimental to sage grouse brood habitat (Klebenow 1970). Xeric sagebrush habitats (I 25 cm of annual precipitation), usually dominated by Wyoming big sagebrush, typically have relatively low forb abundance com- pared to other sagebrush habitats (Wright et al. 1979, Clifton 1981, Bunting et al. 1987). Thus, insects as a source of protein in sage grouse chick diets may have greater importance in xeric sagebrush habitats than in more mesic habitats. Reduced insect abundance through pesticide application has been implicated in lowered chick survival in several gallinaceous bird species (Godfrey 1975, Warner et al. 1984, Potts 1986). Johnson and Boyce (1990) evaluated the influence of insect reductions on sur- vival of captive sage grouse chicks, and quantity of insects in the diets was positively correlated with growth and survival.
Klebenow (1972) and Gates (1983) suggested that fire may benefit sage grouse brood habitat if the burn produces a mosaic of sagebrush cover interspersed with open areas having increased forb production. However, the impacts of fire on sage grouse foods, especially insects, have not been addressed. Objectives of this study were to (1) examine relative use of burned ungrazed habitat and unburned grazed habitat by sage grouse following a prescribed fire, and (2) measure summer forb and insect abun- dance in burned and unburned sage grouse habitat as an index to food abundance.
Methods Study Location and Burning Treatments
From 1990 to 1992, we conducted research on a 20,000-ha por- tion of the 240,867-ha Big Desert, in Blaine and Butte counties on the Upper Snake River Plain, southeastern Idaho (43” 24’ N,
113” 07’ W). The topography consisted of flat to gently undulat- ing terrain with an interspersion of exposed silicic and basaltic volcanic outcrops and craters. Median precipitation on the Big Desert during the study (16.5 cm; 1990-92), measured 25 km
JOURNAL OF RANGE MANAGEMENT 49(3), May 1996
northwest of the study area in similar habitat, was below the long-term median (24.0 cm; 1956-i-86 [data from Agricultural Engineering Dept., Univ. Idaho]). Peak precipitation typically occurred from April through June, although there was consider- able seasonal and annual variation. Elevations ranged from 1,536 m to 2,304 m. Hironaka et al. (1983) classified the Big Desert as a Wyoming big sagebrushibluebunch wheatgrass (Agropyron spi- cuturn Scribn. & Smith) habitat type; threetip sagebrush (A. fri- pa&z Rydb.), rabbitbrush (Chrysofhmnus spp Nutt.), Sandberg bluegrass (Poa sandbergii Vasey), and bottlebrush squirreltail (Situnion hysrtir Nutt.) were abundant. The Big Desert sagebrush ecosystem provides important sage grouse breeding, nesting, brood-rearing, and wintering habitat (Connelly et al. 1988, Wakkinen 1990, Robertson 1991). Sage grouse on the study area used contiguous areas for wintering and breeding, but migrated as far as 85 km to summering areas (Wakkinen 1990, Fischer 1994).
During late-summer 1989, the Bureau of Land Management burned approximately 5,800 ha of the northern portion of the study area. Vegetation was removed from 57% of this total area, resulting in a mosaic of burned and unburned vegetation (J. W.
Connelly, Idaho Dept. of Fish and Game, unpubl. data). Another site of approximately equal size was 6 km south of the burned area, and was not altered during the study. Mean prebum sage- brush canopy cover was 19.8% and grass cover was 6.4% (J. W.
Connelly, Idaho Dept. Fish and Game, unpubl. data). There was a potentially confounding influence of differential grazing history on these 2 areas. All portions of the study area were grazed by cattle throughout the study except for the burned area, which was ungrazed from fall 1988 to fall 1991. Livestock exclusion for at least 2 years following prescribed bums is a suggested practice in big sagebrush communities (Wright et al. 1979). Wakkinen (1990) and Robertson (1991) provided detailed descriptions of the Big Desert study area.
Bird Censuses
We counted adult sage grouse and broods weekly on two, 20 to 25 km routes through unburned habitat and 1, 21-km route through burned habitat, from mid-May through mid-August, 1990 to 1992. The census route through burned habitat included both burned and unburned areas within the vegetation mosaic. Routes were conducted during the evening with either a truck or all-ter- rain vehicle. All adult sage grouse on census routes were record- ed by sex, and brood sizes were noted. We converted observa- tions for each route to relative abundance (number per km) of males, females, and broods, to standardize for different route dis- tances. Data were available from identical preburn census routes conducted during each July, 1987 to 1989 (J. W. Connelly, Idaho Dept. of Fish and Game, unpubl. data). Relatively few grouse were observed on July postburn census, producing many “zero”
observations. Also, the proportion of grouse remaining in the study area during summer varied with precipitation and plant moisture content (Fischer 1994). These 2 factors precluded a sta- tistical comparison of pre- and postburn July brood data.
However, means for each variable during July, 1987 to 1992, were plotted for a visual comparison.
Food Abundance Insects
Concurrent with brood censuses, we sampled arthropods using 3 methods. We used a 0.38-m diameter sweep net (for grass and forb-dwelling arthropods) and 0.71- x 0.71-m beating sheet (for
shrub-dwelling arthropods) to sample arthropods at brood loca- tions along 1990 census routes. When a brood was observed along a census route, we marked the activity center, and returned to the location at approximately the same time the next day to collect arthropods. We placed the beating sheet under the closest sagebrush shrub in each cardinal direction, and beat all branches within 0.5 m of the ground to remove any arthropods. We then used the sweep net to make 10 sweeps through grasses and forbs in each cardinal direction from the activity center. Samples from each transect (arthropods, shrub detritus, and vegetation) were sealed in separate, air-tight bags. To determine potential differ- ences in insect abundance between brood and non-brood loca- tions, we sampled 6 random sites (3 burned, 3 unburned) each week (May through July) within the study area, using identical sampling methods. Only 7 broods were observed during all 1991 and 1992 census routes, which precluded sufficient arthropod sampling at brood locations during these years.
We also employed a weekly pitfall trapping method at 8 ran- dom locations (4 burned, 4 unburned) within the study area, May through July, 1990 to 1992, to assess differences in ground- dwelling insect abundance between burned and unburned sage grouse habitat. However, to reduce the possibility of bias associ- ated with reduced arthropod populations by repeated sampling at the same sites, we chose 2 additional sub-sites near each of the 8 initial random sites by walking in a randomly generated direction and distance (s 100 m) from the initial site. We then established a 4 x 4 test tube pitfall grid with line intersections at 2-m intervals at each of the 24 sites, and sampled each site every third week (8 sub-sites per week). Test tubes were filled with a 1:l solution of ethylene glycol and water, then buried flush with the ground. We opened traps in the morning, and closed and removed them the evening of the following day. Samples from all tubes at each grid were combined and stored in closed vials filled with a 70%
ethanol solution for 1 to 3 months before analysis. Arthropod sampling techniques follow Cooper and Whitmore (1990).
In the laboratory, insects (Class Insecta) were separated from shrub detritus, vegetation, and other arthropods with the aid of magnifying glasses. Because we were specifically interested in the effects of fire on sage grouse foods, we separated insects into the 3 major Orders [Hymenoptera (ants), Coleoptera (beetles), and Orthoptera (grasshoppers)] known to be important in juvenile sage grouse diets (Klebenow and Gray 1968, Peterson 1970).
Numbers of individuals in each sample were then counted.
Forbs
Twenty-two 50-m permanent transects were established 2 months before the 1989 fire at random locations in (n = 12) and adjacent to (n = 10) the area to be burned. Vegetation characteris- tics were measured on all transects in late June/early July, 1989 to 1993 (K. P. Reese, unpubl. data). Thus, there was 1 year of prebum and 4 years of postbum vegetation data on the burned and unburned habitat. Along each transect, forb cover was sam- pled using 0.2 x 0.5-m plots (Daubenmire 1959) spaced at I-m intervals. Forbs were categorized as major forbs, those species known to be important in sage grouse diets (common dandelion [Taraxacum oficinale Weber], milkvetch [Astragulus spp.], com- mon salsify [Tragupogon spp.], sego lily [Culochortus spp.], common yarrow [Achillea spp.], prickly lettuce [Lacruca spp.], lupine [Lupinus spp.], hawksbeard [Crepis spp.]) (Klebenow and Gray 1968, Peterson 1970, Wallestad et al. 1975), and minor forbs, those species not known to be important food components.
JOURNAL OF RANGE MANAGEMENT 49(3), May 1996 195
Statistical Analysis
We used either SAS (SAS Inst., Inc. 1988) or STATISTIX (Analytical Software, Inc., St. Paul, MN) to analyze data, and considered differences significant if P 5 0.05. All variables were also tested for normality using a Kolmogorov-Smimov D-statistic (Stephens 1974), and appropriate transformations were used to approximate the normal distribution before statistical tests (Zar 1984). To assess differences between burned and unburned habi- tat, we used a repeated measures analysis of variance (ANOVA, PROC GLM) with orthogonal contrasts to compare potential dif- ferences in relative abundance of males, females, and broods between areas after the fire, and potential differences in major and minor forb cover before and after fire. We used a mixed- model ANOVA to compare insect abundance between brood and non-brood locations (sweep net/beating sheet), and between burned and unburned locations (pitfalls). Because few Orthoptera were captured with any collection method, we used a binomial test on frequency of nonzero data to determine if Orthoptera were captured more or less frequently in either area, and a t-test or Mm-Whitney-U test (depending on normality) on nonzero data to detect differences in abundance among samples in which Orthoptera were captured. We tested all main effects and interac- tions using appropriate error terms.
than non-brood sites with a sweep net, but there were no differ- ences in abundance between burned and unburned areas at sites where Orthoptera were captured (Mann-Whitney-U test; P = 0.805). No difference in Coleoptera abundance between brood and non-brood locations was detected, however these 2 tech- niques are not effective in capturing ground-dwelling arthropods such as most beetle species. Relative abundance of Hymenoptera and Coleoptera captured in pitfall traps was similar between burned and unburned areas in 1990, the first year following the fire (F1,gl = 0.01, P = 0.920; F, 89 = 0.58, P = 0.450, respective- ly). However, there were signkicantly fewer Hymenoptera in burned habitat the second (F = 9.20, P = 0.003) and third (F = 17.11, P = 0.0001) years postburn (Table 2). There were no dif- ferences in Orthoptera capture frequency or abundance in 1991 or 1992 pitfall traps (Table 2). Coleoptera abundance was similar between burned and unburned areas during all 3 postbum years.
Hymenoptera abundance was similar among months in burned (ANOVA; F2,139 = 0.83, P = 0.437) and unburned areas (Fz,136 = 0.13, P = 0.875), as was frequency of Orthoptera captures.
Forbs
ReSlllt.9
There was variation in annual major and minor forb cover, however, there were no significant differences in either variable (Ft,g = 1.55, P = 0.216; Fig = 0.37, P = 0.544, respectively) before or after the fire (Fig. 1).
Bird Censuses Discussion
From 1987 to 1992, the relative abundance of broods declined in burned and unburned habitat, except for 1990. Visual compar- isons of July pre- and postbum data revealed no large differences in relative abundance of males, females, and broods between areas. After the fire, the relative abundance of males (F = 2.91, P
= 0.20), females 0; = 3.49, P = 0.15), and broods (F = 1.26, P = 0.25) observed on routes was also similar between burned and unburned habitat (Table 1).
Food Abundance Insects
The abundance of Hymenoptera at 1990 brood activity areas (n
= 33) was higher than at non-brood locations (n = 74) using both the sweep net (x = 7.03 vs. x= 3.51, P = 0.019) and beating sheet (x = 12.90 vs. x= 2.93, P = 0.022). Orthoptera were captured more frequently (binomial test; P = 0.024) at brood activity areas
Prescribed fire created a patchy mosaic of sagebrush inter- spersed with open areas containing forbs and grasses (K.P. Reese, Univ. of Idaho, unpub. data). Bunting et al. (1987) found that forb cover in Wyoming big sagebrush habitats is typically low, and these xeric habitats have little potential for forb increase fol- lowing fue, relative to the more mesic sagebrush habitats. Our forb cover data before and after fire were consistent with these findings. Martin (1990) found a significant increase in forbs the first 2 years following fire in Eastern Idaho, but his study was conducted in a more mesic area (i.e., 30-50 cm annual precipita- tion) dominated by threetip sagebrush and mountain big sage- brush (A. f. vuseyuna Nutt.). Hymenopteran abundance was lower within the burned portion of our study area, suggesting that fire negatively influenced insects important in sage grouse diets.
Table 1. Number of sage grouse broods; distance travelled; and relative abundance of broods, females, and males on survey routes in burned and unburned habitat, Big Desert, southeastern Idaho (1987 to 1992).
Number of Broods Unburned Bumed
Distance travelled1 Unburned Bumed
Broods Unburned Bumed
Relative Abundance
Females Males
Unburned Bumed Unburned Bumed Prebum (July) ---(km) ---- - ---___________ (nm&perhx 10-3) __________________
1987 43 13 688 614 63 21 58 125 18 44
1988 10 3 494 329 20 91 57 27 2 3
1989 6 2 317 209 19 100 181 96 17 38
postbum (May through July)
1990 22 6 666 449 33 13 14 5 14 11
1991 4 1 880 579 5 2 28 4 10 0
1992 2 0 777 516 3 0 0 0 0 0
1 Total dlstmce travelled each year
196 JOURNAL OF RANGE MANAGEMENT 49(3), May 1996
Table 2. Comparison of insect abundance from pi&ii trap samples at burned versus unburned locations, Big Desert, southeastern Idaho (May to July, l990 to 1992).
YW
1990 cn 9% 1991 In= 851 1992 In = 101)
Order Raw dafar Logb Raw data Log Rawdata Log
Hymenoptera
BUtTled 241.0 2.24a 191.7 2.20a 86.1 l.Sa
Unburned 261.7 2.23a 289.8 2.3Sb 151.7 2.lb
Coleoptera
Burned 3.8 0.56a 2.7 0.47a 2.9 OSa
Unburned 3.9 OSOa 2.4 0.44a 2.0 0.4a
ortbopteraC
BtKIled -d - 1.sa - 1.3” -
Unburned - - 1.5” - 1.7a -
3 Numbers indicate mean number of insects captured in pi&II grids per week.
b Statistid comparisoas were conducted on log-traasformed dataA pair of means ia a column followed by a different letter is significaatly different (P < 0.05).
c Few Orthopten were captured in pitfall tmps.A binomial test on frequency of nonzero L ‘13 was not significant in 1991or 1992.hIeaas are reported for a t-test on noazero data
Data not collected
Drought probably also influenced both forb and insect popula- tions on our study area, given the low precipitation (31.3% below long-term median) during the study. Because there was no posi- tive response (i.e., increase in relative abundance) of sage grouse to burned habitat following the fire, our results from a xeric envi- ronment did not support Klebenow’s (1972) and Gates’ (1983) speculation that fire may benefit sage grouse brood habitat.
However, our data were collected under a pseudoreplicated design (1 control, 1 treatment); true replication in this study was not possible. We investigated a migratory population that collec- tively ranges over several thousand square miles during the year.
Additional prescribed fries, regardless of their size, within the range of this population would not have been true replicates, but rather sub-samples. Additional bums at the scale used in this study also would have occurred in different precipitation zones, sagebrush habitat types, or sage grouse populations. Thus, addi- tional research is needed to fully understand the influence of fire on sage grouse brood-rearing habitat, especially in other sage- brush habitat types and under conditions not encountered in the present study.
Relatively few Orthoptera were captured in our study area (brood or non-brood sites) using any of the collection methods, suggesting low availability for Big Desert sage grouse. Bock and Bock (1991) found reduced Orthoptera abundance 1 year follow- ing fire in an Arizona grassland, but no difference 3 years post- bum. Coleoptera abundance in our study area was also low in burned and unburned habitat and appeared unaffected by fire.
However, Rickard (1970) investigated Coleoptera abundance in burned and unburned sagebrush habitat and found more beetles in unburned habitat. Lower abundance and biomass of Hymenoptem in the burn the second and third years postbum suggested that fire negatively influenced insects important in juvenile sage grouse diets for several years following fire. We did not sample insect abundance prior to the fire, thus we cannot attribute differences in abundance between areas to fire. However, because insect abun- dance was similar between areas the first year following the fire, and Hymenoptera abundance significantly decreased with time, a difference between burned and unburned habitat was indicated.
Timing of breeding activities may correspond to temporal
JOURNAL OF RANGE MANAGEMENT 49(3), May 1996
abundance of food resources so that chicks hatch when insects are most abundant (Lack 1954, Poulin et al. 1992). Timing of sage grouse hatching may also coincide with peak insect abun- dance during the spring. We found that Coleoptera abundance was highest in late May/early June and declined steadily over the summer. Hymenoptera and Orthoptera abundance remained fairly stable over this same period. In Idaho, Stafford (1983) sampled summer Coleoptera abundance and found highest numbers occur- ring during the first 2 weeks of June, followed by a decline in late summer. Peak hatching of sage grouse nests in our study was late May/early June (Wakkinen 1990), which would correspond with peak abundance of important insects. Dalke et al. (1960) also found peak hatch during late May/early June in eastern Idaho.
Other important factors that probably influenced timing of sage grouse nesting in xeric environments include renesting opporhmi- ties (Connelly et al. 1993), snowmelt, and plant-phenology.
Conchsions
The results of our research indicated that the short-term effects of prescribed fire in a xeric environment did not enhance brood- rearing habitat, and may have been detrimental to Hymenoptera which are important in sage grouse diets. Prescribed fire can vari- ably affect sagebrush-grass habitats depending on the intensity, duration, location, shape, and extent of the fire (Bendell 1974,
Minor Forb Burned
. . .** .e.... l.. .-
3
iMajor Forb Unburned
t Major Forb Burned
1990 1991 1992 1993
Fig. 1. Abundance of major forbs and minor forbs, expressed in Daubenmire classes (1 = 05% cover; 2 = 525% cover; 3 = 25 50% cover), measured on permanent transects in burned and unburned sage grouse habitat on the Big Desert, southeastern Idaho, 1990 to 1993. The burn occurred after the 1989 data were collected.
197
Sapsis and Kauttmann 1991), and environmental conditions, species composition, and available fuel (Winward 1985). Caution is needed when using tire in xeric sagebrush habitats (i.e., I 25 cm ammal precipitation) as a management tool for sage grouse brood habitat because forb cover and abundance may not be enhanced by burning. Because the response of sagebrush/grass vegetation to fire is highly dependent on geographic area and sagebrush subspecies (Bunting et al. 1987), forbs and insect pop- ulations in more mesic sage grouse habitats may respond differ- ently to tire.
Literature Cited
BendeB, J.F. 1974. Effects of fire on bii and mammals, pp. 73-138. In:
T. T.Kozlowski and C. E. Ahlgren (eds.), Fire and ecosystems.
Academic Press, N.Y.
Bock, C.E. and J.H. Bock. 1991. Response of grasshoppers (Orthoptera:
Acrididae) to wildfire in a southeastern Arizona grassland. Amer.
hlidl. Natur. 125:162-167.
Bunting, S.C., B.M. Kiigore, and C.L. Bushey. 1987. Guidelines for prescribed burning in sagebrush-grass rangelands in the Northern Great Basin. USDA Forest Service, Intermountain Research Station, Gen. Tech. Rep. INT-231. Ogden, Utah.
Clifton, N.A. 1981. Response of a prescribed fire in a Wyoming Big Sagebmsh5luebunch Wheatgrass habitat type. M.S. Thesis, Univ. of Idaho. Moscow, Ida.
Couuelly, J.W., H.W. Browem, and R.J. Gates. 1988. Seasonal move- ments of sage grouse in southeastern Idaho. J. Wildl. Manage.
52:1X-122.
Connelly, J.W., R.A. Fischer, A.D. Apa, K.P. Reese, and W.L.
Wakkinen. 1993. Renesting of sage grouse in southeastern Idaho.
Condor 95:1041-1043.
Cooper, RJ. and RC. Whitmore. 1990. Arthropod sampling methods in omithology. Studies in Avian Biol. 13:29-37.
Dalke, P.D., D.B. Pyrah, D.C. Stanton, J.E. Crawford, and E.F.
Schiatterer. 1960. Seasonal movements and breeding behavior of sage grouse in Idaho. Trans. N.Amer. Wildl. Conf. 25396-406.
Dauhenmire, R.F. 1959. A canopy-coverage method of vegetation analysis. Northwest Sci. 33:43-64.
Fischer, R.A. 1994. The effects of prescribed fire on the ecology of migratory sage grouse in southeastern Idaho. Ph.D. Diss., Univ. of Idaho, Moscow. Ida.
Frandsen, O.A. 1985. Fiie as a management tool in Southeast Idaho, pp.
85-87. In: K. Sanders and J. Durham (eds.), Rangelaud fire effects:
Proceediugs of a symposium. Boise, Ida.
Gates, J.G. 1983. Sage grouse, lagomorph and pronghom use of a sage- brush-grassland burn site on the Idaho National Engineering Laboratory. M.S. Thesis, Mont. State Univ. Bozemau. Mom.
Gleason, R.S. 1978. Aspects of the breeding biology of burrowing owls in southeastern Idaho. M.S. Thesis, Univ. of Idaho. Moscow, Ida.
Godfrey, G.A. 1975. Home range characteristics of r&fed grouse broods in Minnesota. J. Wildl. Manage. 39:287-298.
Halford, D.K. 1981. Repopulation and food habits of Perornyscus man- icularus on a burned sagebrush desert in southeast Idaho. Northwest Sci. 55&l-49.
Hironaka, M., M.A. Fosberg, and A.H. Winward. 1983. Sagebmsh- grass habitat types in southern Idaho. University of Idaho, Forestry, Wildlife, and Range Exp. Sta. Bull. 35.
Horning, D.S. Jr., and W.F. Barr. 1970. Insects of Craters of the Moon National Monument, Idaho. Univ. of Idaho College of Agr. Misc. Ser.
No. 8.
Johnson, G.D. and M.S. Boyca 1990. Feeding trials with insects in the diet of sage grouse chicks. J. Wildl. Manage. 5489-91.
King, R.D. 1969. Spring and summer foods of ruffed grouse on Vancouver Island. J. Wildl. Manage. 33:4404%2.
Klebenow, D.A. 1970. Sage grouse versus sagebrush control in Idaho. J.
Range Manage. 23:396XlO.
Klebenow, D.A. 1972. The habitat requirements of sage grouse and the role of fire in management. Proc. Ann. Tall Timbers Fiie Ecol. Conf.
12:305-315.
Kiebenow, D.A. and G.M. Gray. 1968. Food habits of juvenile sage grouse. J. Range. Manage. 21:SO-S3.
Kobriger, G.D. 1965. Status, movements, habitats, and foods of prairie grouse on a samlhills refuge. J. Wildl. Manage. 29: 78&800.
Lack, D. 1954. The natural regulation of animal numbers. Oxford University Press, London.
Martin, B.C. 1990. Sage grouse responses to wildfire in spring and sum- mer habitats. M.S. Thesis, Univ. Idaho. Moscow, Ida
Peterson, J.G. 1970. The food habits and summer distribution of juve- nile sage grouse in central Montana. J. Wildl. Manage. 34~147-155.
Potts, G.R 1986. The partridge. Pesticides, predation and conservation.
Collins, London.
Pouiin, B., G. Lefehvre, and R McNeil. 1992. Tropical avian phenolo- gy in relation to abundance and exploitation of food resources.
Ecology. 73:2295-2309.
Raley, C.M. and S.H. Anderson. 1990. Availability and use of arthro- pod food resources by Wilson’s Warblers and Lincoln’s sparrows in southeastern Wyoming. Condor 92:141-150.
Richard, W.H. 1970. Ground-dwelling beetles in burned and unburned vegetation. J. Range Manage. 23:293-294.
Robertson, M.D. 1991. Winter ecology of migratory sage grouse and associated effects of prescribed fire in southeastern Idaho. M.S. Thesis, Univ. of Idaho. Moscow, Ida.
Sapsis, D.B. and J.B. Kauffman. 1991. Fuel consumption and fire behavior associated with prescribed fires in sagebrush ecosystems.
Northwest Sci. 65:173-179.
SAS Institute, Inc. 1988. SAS/STAT User’s Guide, Release 6.03 Edition. SAS Institute Inc., Cary, NC.
Stafford, M.P. 1983. Surface-dwelling Coleoptera inhabiting sagebrush iu southeastern Idaho. M.S. Thesis, Univ. of Idaho. Moscow, I&.
Stephens, M.A. 1974. EDF statistics for goodness of fit and some com- parisons. J.Amer. Stat. Assoc. 69:730-737.
Swanson, G.A., M.I. Meyer, and V.A. Adomaitis. 1985. Foods con- sumed by breeding mallards on wetlands of south-central North Dakota. J. Wihll. Manage. 49:197-203.
Wakkinen, W.L. 1990. Nest site characteristics and spring-summer movements of migratory sage grouse in southeastern Idaho. M.S.
Thesis, Univ. of Idaho. Moscow, Ida.
WaBestad, RO., J.G. Peterson, and RL. Eng. 1975. Foods of adult sage grouse in central Montana. J. Wildl. Manage. 39:62S-630.
Warner, RE., S.L. Etter, G.B. Joselyn, and J.A. Eiiis. 1984. Declining survival of ring-necked pheasant chicks in Illinois agricultural ecosys- tems. J. Wildl. Manage. 48:82-S%
Winward, AH. 1985. Fiie in the sagebrush-grass ecosystem- the eco- logical setting, pp. 2-6. In: K. Sanders and J. Durham (eds.), Rangelaud fire effects: a symposium; Proceedings. Boise, Ida.
Wright, H.A., L.F. Neuenschwander, and C.M. Britton. 1979. The role and use of fire in sagebrush-grass and pinyon-jumper plant com- munities: a state-of-the-art review. USDA Forest Ser., Intermountain Forest and Range Exp. Station. Gen. Tech. Rep. IN’I-58. Ogden, Utah.
Zar, J.H. 1984. Biostatistical Analysis. 2nd ed. Prentice-Hall, Englewood Cliffs, N.J.
198 JOURNAL OF RANGE MANAGEMENT 49(3), May 1996
J. Range Manage.
49:199-203
Shrub-grassland small mammal and vegetation responses to rest from grazing
STEVEN S. ROSENSTOCK
Author is research biologist, Arkona Game and Fish Department, 2221 W. Greenway Rd. Phoenix, A& 85023. At the time this study was conducted, he was independent consultant, dba Wingate Eco-Logic, P.O. Box 462, Fredonia, A&
86022.
Abstract
Between 1989-1991, I studied the effects of livestock grazing on vegetation and small mammals in semiarid shrub-grassland habitats of south-central Utah. Responses were measured at 2 spatial habitat scales; patches and macrohabitats. Patch-scale data were obtained from 4 small (cl ha) livestock exclosures and nearby grazed areas. Macrohabitat-scale data were collected at 4 actively grazed sites and 4 comparable, excellent condition sites, ungrazed for 30+ years. Ungrazed patch and macrohabitat sites had more surface litter, greater perennial grass cover, and taller perennial grass plants, but treatment response varied among sites. Small mammal responses were apparent only at the macro- habitat scale, where ungrazed sites had 50 % greater species rich- ness and 80% higher abundance. Small mammal reproductive activity and biomass were not affected by rest from grazing at either scale. Small mammal community composition varied greatly among sites and within treatments. This variability has important implications for ecological monitoring efforts involv- ing these species.
Key Words: small mammals, grazing effects, scale, Colorado Plateau
Livestock grazing has been a pervasive influence on shrub- grassland communities of the Intermountain United States (West 1988). Previous studies have shown that livestock grazing can significautly affect the floristic composition and vegetative struc- ture of these habitats (Kleiner and Harper 1972, Tuhy and MacMahon 1988). Recently, considerable attention has been focused on the effects of livestock grazing on faunal abundance and diversity in arid rangelands (Fleischner 1994).
Because small mammal communities can be sampled relatively easily, they have been used as indicators to monitor ecosystem responses to management (Douglass 1984, Olson et al. 1994).
Small mammals may also be useful ecological units with which to examine the effects of grazing on biodiversity in arid environ- ments. Small mammals select habitat at multiple spatial scales;
Research was supported by the U.S. Department of the Interior, National Park Service. Valuable assistance was provided by Norm Henderson, Mike Bogan, Susi hlacvean, Tii Grabam, David Willey. and Bill Booker. The author thanks Brian Wakeling and 3 anonymous reviewers for constructive comments on earlier ver- sions of tbis manuscript.
hianuscript accepted 11 Sept. 1995.
JOURNAL OF RANGE MANAGEMENT 49(3), May 1996
microhabitats, patches, and macrohabitats (Price 1978, Brown 1987), and are strongly influenced by habitat structure (Rosenzweig and Winakur 1969, Bowers and Flanagan 1988).
Selection is typically for habitat components providing favorable microclimates, food, cover, and avoidance of competitors (Brown 1989, Brown et al. 1988); many of which can be directly or indi- rectly affected by grazing.
Small mammal responses to grazing in shrub-grasslands of the Colorado Plateau have not been previously described. To address this information gap, I examined changes in shrub-grassland veg- etation and small mammal communities, following rest from grazing. I measured small mammal community and demographic parameters at 2 spatial habitat scales, patches and macrohabitats.
These scales reflected distinct changes to which small mammals might respond. Patches were defined as isolated ungrazed sites
>l ha, smaller than the home ranges of resident small mammal species. If ungrazed patches provided more resources (food or cover) than available in the surrounding grazed area, they should receive greater small mammal use. Animals using these patches could also have a higher level of reproductive activity during the breeding season. I also hypothesized that similar responses would occur at a macrohabitat scale. I defined macrohabitats as exten- sive tracts (>lOO ha) of the same shrub-grassland habitat, large enough to contain the home ranges of numerous small mammals.
Methods Study Areas
Study sites were located in Capitol Reef National Park (38” 15”
N 111” 15’ W), Canyonlands National Park (38” 15” N 109” 55”
W), and adjacent lands administered by the Bureau of Land Management (BLM), in Wayne and Grand Counties, south-cen- tral Utah, the northwest portion of the Colorado Plateau. The cli- mate of this area is semiarid, temperate continental, with average annual precipitation of 17 cm per year (U.S. Weather Bureau, Climate and Precipitation Summaries, Utah). Study sites were located on Semidesert Sandy Loam range sites (USDA Soil Conservation Service, Unpublished data), and were dominated by cool-season bunchgrasses (primarily Stipa hymenoides and S.
comata), warm-season sod-forming grasses (primarily Hilaria jamesii and Bouteluoa gracilis), and low-medium stature shrubs
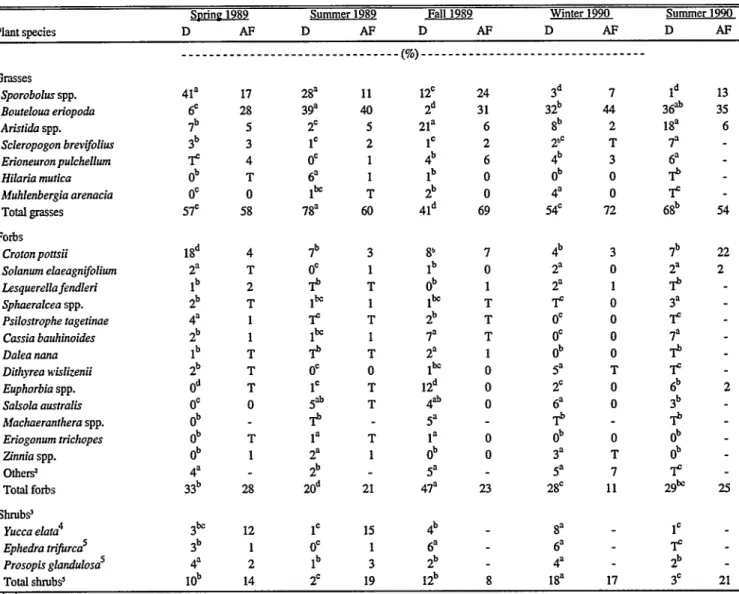
of several genera (primarily Gutierrizia, Ceratoides and Atriplex). Detailed descriptions of these plant communities can be found in Rormne et al. (1993).
199