GABA
B
receptor subtypes differentially regulate thalamic spindle
oscillations
Daniel Ulrich, Txomin Lalanne, Martin Gassmann, Bernhard Bettler
*Dept. Biomedicine, Institute of Physiology, University of Basel, 4056 Basel, Switzerland
a r t i c l e i n f o
Article history: Received 19 June 2017 Received in revised form 23 October 2017 Accepted 24 October 2017 Available online xxx Keywords: KCTD12 KCTD16 GPCR Epilepsy Sleep
a b s t r a c t
Following the discovery of GABABreceptors by Norman Bowery and colleagues, cloning and biochemical efforts revealed that GABABreceptors assemble multi-subunit complexes composed of principal and auxiliary subunits. The principal receptor subunits GABAB1a, GABAB1band GABAB2form two hetero-dimeric GABAB(1a,2)and GABAB(1b,2)receptors that can associate with tetramers of auxiliary KCTD (Kþ channel tetramerization domain) subunits. Experiments with subunit knock-out mice revealed that GABAB(1b,2)receptors activate slow inhibitory postsynaptic currents (sIPSCs) while GABAB(1a,2)receptors function as heteroreceptors and inhibit glutamate release. Both GABAB(1a,2)and GABAB(1b,2)receptors can serve as autoreceptors and inhibit GABA release. Auxiliary KCTD subunits regulate the duration of sIPSCs and scaffold effector channels at the receptor. GABABreceptors are well known to contribute to thalamic spindle oscillations. Spindles are generated through alternating burst-firing in reciprocally connected glutamatergic thalamocortical relay (TCR) and GABAergic thalamic reticular nucleus (TRN) neurons. The available data implicate postsynaptic GABABreceptors in TCR cells in the regulation of spindle frequency. We now used electrical or optogenetic activation of thalamic spindles and pharmacological experiments in acute slices of knock-out mice to study the impact of GABAB(1a,2)and GABAB(1b,2)receptors on spindle oscillations. We found that selectively GABAB(1a,2)heteroreceptors at TCR to TRN cell synapses regulate oscillation strength, while GABAB(1b,2) receptors control oscillation frequency. The auxiliary subunit KCTD16 influences both oscillation strength and frequency, supporting that KCTD16 regulates network activity through GABAB(1a,2)and GABAB(1b,2)receptors.
©2017 Elsevier Ltd. All rights reserved.
1. Introduction
Elegant pharmacological and biochemical experiments by Nor-man Bowery and colleagues revealed the existence of GABA re-ceptors coupled to G-proteins (Bowery et al., 1980, 1987; Hill and Bowery, 1981; Hill et al., 1984). They named these receptor GABAB
receptors to distinguish them from the known GABAA receptors.
Nearly two decades later cloning efforts revealed the existence of three GABAB receptor subunits that form two heterodimeric
GABAB(1a,2)and GABAB(1b,2)receptors (Kaupmann et al., 1997, 1998;
White et al., 1998). The two receptors exhibited similar pharma-cological and functional properties when expressed in transfected heterologous cells. However,GABAB1a/andGABAB1b/knock-out mice
showed that GABAB(1a,2)and GABAB(1b,2) receptors fulfill distinct
pre- and postsynaptic functions in the brain. Specifically, knock-out
mice revealed that predominantly GABAB(1b,2) receptors activate
postsynaptic Kþchannels and mediate slow inhibitory postsynaptic currents (sIPSCs) (reviewed in Gassmann and Bettler, 2012). GABAB(1a,2)receptors were shown to function as heteroreceptors
and to restrict glutamate release by inhibiting voltage-sensitive Ca2þ-channels. Both GABAB(1a,2) and GABAB(1b,2) receptors can
inhibit GABA release and function as autoreceptors. GABAB(1a,2)and
GABAB(1b,2) receptors were later shown to associate with the
auxiliary subunits KCTD8, -12, -12b and -16 (Schwenk et al., 2010, 2016; reviewed byPin and Bettler, 2016). The KCTD subunits in-fluence activation/deactivation kinetics and desensitization of the receptor response (Fritzius et al., 2017; Turecek et al., 2014). Moreover, KCTD16 was shown to scaffold effector Ca2þ and hyperpolarization-activated cyclic nucleotide gated (HCN) channels at the receptor (Schwenk et al., 2016). Knock-out mice demon-strated that KCTD12 and KCTD16 modulate the postsynaptic GABAB
receptor response and alter the duration of sIPSCs (Booker et al., 2017; Fritzius et al., 2017; Schwenk et al., 2016). Morphological *Corresponding author.
E-mail address:[email protected](B. Bettler).
Contents lists available atScienceDirect
Neuropharmacology
j o u r n a l h o m e p a g e :w w w . e l s e v i e r . c o m / l o c a t e / n e u r o p h a r m
https://doi.org/10.1016/j.neuropharm.2017.10.033 0028-3908/©2017 Elsevier Ltd. All rights reserved.
data support that KCTD12 and KCTD16 also regulate presynaptic GABABreceptor responses (Schwenk et al., 2010).
Because GABABreceptors are mostly localized perisynaptically
they normally require synchronous release of GABA from several synapses to be activated, which typically occurs during high fre-quency action-potential bursts (reviewed in Ulrich and Bettler, 2007). In line with a dependence on burst-firing for activation, GABABreceptors were shown to contribute to brain oscillations,
including slow, theta and gamma oscillations (reviewed byCraig and McBain, 2013; Kohl and Paulsen, 2010). It has long been known that GABABreceptors influence brain oscillations in the
thalamus. The thalamus generates transient 7e14 Hz oscillations during the early stages of non-REM (rapid eye movement) sleep (Steriade and Desch^enes, 1984; Steriade and Llinas, 1988). These oscillations, known as sleep spindles, are one of several rhythms arising in the thalamus during non-REM sleep (McCormick and Bal, 1997; Steriade and Desch^enes, 1984; Steriade et al., 1993). It was noted that GABAB receptors exert a pro-oscillatory effect on
thalamic oscillations during absence seizures - a hypersynchronous aberrant form of sleep spindle oscillations (Steriade et al., 1993). In line with thisfinding, GABABagonists and antagonists were shown
to exacerbate and suppress absence seizures, respectively (Hosford et al., 1992). Thalamic spindles are generated through alternating burst-firing in reciprocally connected thalamocortical relay (TCR) cells and GABAergic neurons of the thalamic reticular nucleus (TRN) (Fig. 1A). As a result of the feedback circuitry between TRN and TCR cells, bursts in relay neurons produce glutamate receptor-dependent bursts in TRN neurons, which in turn transiently inhibit relay cells, leading to rebound excitation. It was demonstrated that hypersynchronous oscillations can be generated in ferrets by GABABreceptor-mediated rebound burst-firing in TCR cells
origi-nating from action potential bursts in the interconnected TRN cells (von Krosigk et al., 1993). However, the experiments in ferrets were done after blocking GABAAreceptors with bicuculline, which
dis-inhibits TRN neurons (Huguenard and Prince, 1994a) and likely produces a stronger activation of GABABreceptors in TCR cells than
under physiological conditions (Kleiman-Weiner et al., 2009). Activation of GABABreceptors in TCR cells prolongs IPSCs and
re-duces the spindle frequency due to delays of several hundred milliseconds in rebound excitation (Huguenard and Prince, 1994b; Bal et al., 1995). Conversely, antagonizing GABAB receptors
in-creases spindle frequency (Jacobsen et al., 2001).In vivo experi-ments addressing the mechanism of hypersynchronous absence seizures failed to observe a significant contribution of GABAB
re-ceptors to the inhibitory postsynaptic potential (IPSP) in TCR cells (Pinault et al., 1998; Staak and Pape, 2001) thus questioning whether postsynaptic GABABreceptors significantly contribute to
the generation of thalamic oscillations. A recent study reports a GABABreceptor-mediated enhancement of tonic GABAA receptor
currents in TCR neurons, which may influence oscillatory behavior by altering the membrane potential (Connelly et al., 2013). Given these somewhat conflicting data, it is still unclear in which cells and cellular compartments GABABreceptors regulate thalamic spindle
oscillations. Electrophysiological recordings in TCR and TRN neu-rons revealed pre- and postsynaptic GABAB receptor responses
(Porter and Nieves, 2004; Soltesz et al., 1989; Ulrich and Huguenard, 1996). The analysis of knock-out mice supports a typical pre- and postsynaptic distribution of GABAB(1a,2) and
GABAB(1b,2)receptors in the thalamus (Ulrich et al., 2007). In situ
hybridization experiments show that the auxiliary GABABreceptor
subunit KCTD12 is expressed in TCR neurons while KCTD16 is expressed in both TCR and TRN neurons (Metz et al., 2011) (Fig. 1A). Whether the KCTD-mediated modulation of GABABreceptor
re-sponses is able to influence network behavior is unknown. In this study we used knock-out mice to study the contribution
of GABAB(1a,2) and GABAB(1b,2) receptors to electrically or
opto-genetically induced thalamic spindle oscillations. We found that the oscillation strength is determined by presynaptic GABAB(1a,2)
receptors expressed at TCR terminals, while oscillation frequency is regulated by GABAB(1b,2)receptors. KCTD16 regulates both
oscilla-tion strength and frequency, supporting that KCTD16 regulates thalamic spindles through a modulation of GABAB(1a,2) and
GABAB(1b,2)receptors.
2. Materials and methods
2.1. Mice and tissue preparation
All mouse experiments underwent institutional review and were approved by the Veterinary Office of Basel-Stadt. To generate KCTD12/ and KCTD16/ mice expressing channelrhodopsin 2 (ChR2) coupled to an enhanced yellowfluorescent protein (EYFP) under control of the vesicular GABA transporter (VGAT) promoter in GABAergic neurons, the VGAT-ChR2(H134R)-EYFP transgene (Zhao et al., 2011) was crossed into the previously described KCTD12/andKCTD16/mouse lines (Metz et al., 2011; Cathomas et al., 2015, 2017). GABAB1a/ and GABAB1b/ mice were bred as
reported earlier (Vigot et al., 2006). Knock-out and control litter-mates of either sex were used in our experiments. Mice (P21-28) were anaesthetized with isoflurane. To preserve intercellular con-nectivity 400
m
m thick horizontal brain slices containing the thal-amus were prepared on a vibratome (Leica VT1200S, Nussloch, Germany) in a sucrose-rich solution containing 234 mM sucrose, 2.5 mM KCl, 1.25 mM NaH2PO4, 10 mM MgSO4, 0.5 mM CaCl2,26 mM NaHCO3 and 11 mM glucose at 4 C (Aghajanian and
Rasmussen, 1989; Huguenard and Prince, 1994b). The slices were incubated at 34 C for 1 h in a submerged chamber containing artificial cerebrospinal fluid (ACSF) composed of 126 mM NaCl, 26 mM NaHCO3, 2.5 mM KCl, 1.25 mM NaH2PO4, 2 mM CaCl2, 1 mM
MgCl2and 10 mM glucose and then left at room temperature until
use. All drugs were from Tocris (Bristol UK), stored as 103 x concentrated aliquots at20C and added to the superfusate in the final concentrations indicated.
2.2. Electrophysiology
2.2.1. Extracellular recordings
Slices were transferred to an interface recording chamber mounted on an Olympus BX51WI microscope (Olympus, Tokyo, Japan) and superfused at 2 ml/min with ACSF at 34C. Extracellular multiunit recordings (MUR) were obtained with 2e5 M
U
tungsten electrodes (FHC, Bowdoin ME) inserted into the somatosensory thalamus under visual guidance. Voltage signals were amplified and band-passfiltered between 100 Hz and 3 kHz with a differ-ential amplifier (Dagan, Minneapolis, MN) and digitized at 5 kHz with a Digidata 1440A digitizer (Molecular Devices, Sunnyvale, CA). Stimuli (300e700m
A, 1 ms, 0.03 Hz) were applied via bipolar Pt-Ir wire electrodes positioned in the internal capsule. Alternatively, ChR2 was activated by shutter-controlled band-passfiltered light pulses of 30 ms duration via a 5objective at a repetition rate of 0.03 Hz. The aperture diaphragm of thefluorescence light path was used to limit the area of illumination to the TRN.2.2.2. Intracellular recordings
composed of 135 mM CsCH3O3S, 8 mM NaCl, 4 mM Mg-ATP, 0.3 mM
Na3-GTP, 0.1 mM TEA-Cl, 5 mM QX-314 and 10 mM HEPES. A liquid
junctional potential of10 mV was left uncorrected. Cells were voltage-clamped at 60 mV with a Multiclamp700B amplifier (Molecular Devices). Current traces were low-pass filtered at
[image:3.595.101.502.68.580.2]10 kHz and digitized at 20 kHz with a Digidata 1550B digitizer (Molecular Devices). EPSCs were evoked with extracellular monopolar current pulses generated by a custom made isolated current stimulator and applied via a patch-pipette that wasfilled with ACSF.
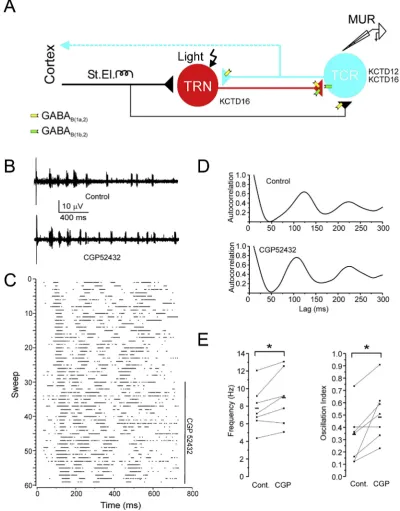
Fig. 1. Increased spindle frequency and strength in WT animals after GABABreceptor blockade.(A) Diagram of the thalamic circuitry. Glutamatergic TCR cells (blue) send their axons to cortex (dashed line) and axon collaterals (solid line) to the TRN (red). TRN neurons make GABAergic synaptic contacts with TCR neurons. Corticothalamic axons project strongly to TRN (thick black line) and weaker to TCR neurons (thin black line). The predominant locations of GABAB(1a,2)and GABAB(1b,2)receptors at pre- and postsynaptic sites are indicated. TRN cells express KCTD16 while TCR cells express KCTD12 and KCTD16. The sites of electrical (St.El.) and optical (Light) stimulation and extracellular multiunit recoding (MUR) are indicated. (B) Sample traces of electrically triggered extracellular MUR activity in the somatosensory thalamus recorded under control conditions and in the presence of CGP52432 (1mM). (C) Raster plot of spikes in consecutive sweeps for the entire experiment. The duration of CGP52432 application is indicated by a vertical bar. (D) Autocorre-lograms of MUR activity under control conditions and in the presence of CGP52432 show enhanced frequency and oscillation strength after blocking GABABreceptors. (E) Pop-ulation data (squares) and means (dashes) of spindle frequency and oscillation strength under control conditions and in the presence of CGP52432 reveal a significant difference. (*) p<0.05, n¼7.
2.3. Data analysis
Extracellular voltage recordings were adjusted for baseline off-sets and a threshold criterion was used to automatically detect spikes (Pclamp 10, Molecular Devices). Post-stimulus time histo-grams (PSTHs) of action potentials were constructed from 5 to 10 consecutive sweeps during stationary periods with a bin width of 10 ms. Autocorrelograms (ACs) of PSTHs were constructed and normalized to the central peak. The amplitude of thefirst satellite peak next to the central peak in the normalized AC was measured from the intervening trough and the resulting value taken as a measure of oscillation strength. The lag of thefirst satellite peak was used to calculate the principal oscillation frequency. EPSC amplitudes were determined by measuring the difference of two visually positioned cursor pairs at baseline and peak.
2.3.1. Statistics
Individual data sets were tested for normality with the Shapiro-Wilk test. Means of drug vs. control groups were compared with a two-tailed pairedt-test, and their variances by an F-test. The means of the different genotypes were compared by ordinary one-way ANOVA with a Tukey post-hoc test, and their variances by Bar-tlett's test.
3. Results
3.1. GABAB(1a,2)and GABAB(1b,2)receptors differentially contribute
to oscillation strength and frequency in the thalamus
For our experiments we used thalamic slice preparations that maintain the reciprocal connectivity between the TRN and TCR cells (Fig. 1A) (Huguenard and Prince, 1994b). We evoked thalamic os-cillations by electrical stimulation of corticothalamic fibers that contact thalamic cells and selected oscillations in the frequency band of sleep spindles (Fig. 1B). Because in our slice experiments the temperature is lower thanin vivothe spindle frequency range was adjusted to 5e11 Hz by applying a temperature coefficient (Q10) of 2.7 (Deboer and Tobler, 1995) to the 7e14 Hz spindle range
in the intact brain. We recorded voltage signals for 15e20 min in control solution, after which we exposed the GABAB receptor
contribution to spindles by adding the antagonist CGP52432 (1
m
M) to the bath. We then continued recording for another 30e45 min. Action potentials were identified offline by visually setting a voltage threshold. We constructed raster plots of spikes for the individual experiments (Fig. 1C) and compiled ACs from PSTHs (Fig. 1D). In wild-type (WT) mice blockade of GABABreceptors withCGP52432 induced a characteristic leftward shift and increase of thefirst satellite peak in the normalized ACs, which indicates an increase in spindle oscillation frequency and strength, respectively (Fig. 1D). All data sets were successfully tested for normality with the Shapiro-Wilk test. Overall the oscillation frequency significantly increased from 7.7±0.9 Hz to 8.9±1.1 Hz (mean±s.e.m, n¼7, p<0.05, pairedt-test) (Fig. 1E). At the same time the oscillation strength was significantly augmented from 0.34 ± 0.06 to 0.48 ± 0.06 (p< 0.05) (Fig. 1E). These results show that GABAB
receptor activity decelerates thalamic spindle oscillations and lowers their synchronicity. While the former could be explained by the decelerating role of GABABreceptors in rebound burst-firing of
TCR cells (von Krosigk et al., 1993) the latter may be attributable to their function in inhibiting glutamate and/or GABA release (Ulrich and Huguenard, 1995). To discriminate between the two possibil-ities, we used GABAB1a/ and GABAB1b/ mice that exhibit specific
deficits in pre- and postsynaptic receptor functions (reviewed in Gassmann and Bettler, 2012).
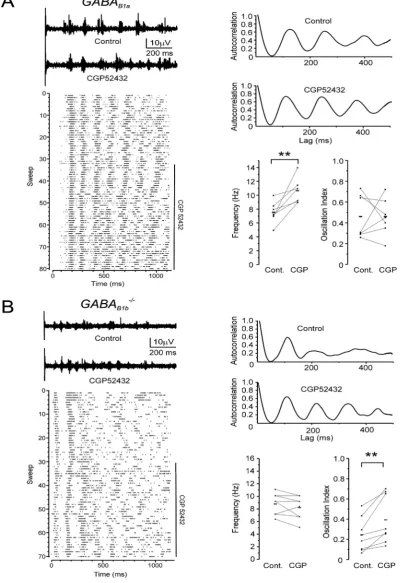
Wefirst performed similar experiments as described above in
GABAB1a/ mice that largely lack heteroreceptor function (Ulrich
et al., 2007) (Fig. 2A). We elicited spindle oscillations similar as in WT mice and compiled ACs of spike trains (Fig. 2A).GABAB1a/mice
exhibited an increase in oscillation frequency after CGP52432 application but, in contrast to WT littermates, the oscillation strength remained unchanged (p>0.5). The oscillation frequency increased from 7.5 ±0.6 Hz to 10.6 ± 0.7 Hz (n ¼7, p< 0.01) indicating that GABAB(1b,2) receptor-activity is rate-limiting for
sleep spindles (Fig. 2A). In separate experiments we elicited spin-dles in thalamic slices ofGABAB1b/mice (Fig. 2B) that largely lack
sIPSCs (reviewed in Gassmann and Bettler, 2012). Remarkably, blockade of GABABreceptors inGABAB1b/mice led to a significant
increase in oscillation strength while spindle frequency was unal-tered (p>0.1) (Fig. 2B). The oscillation strength inGABAB1b/mice
increased from 0.24±0.06 to 0.39±0.08 (p<0.01, n¼8) (Fig. 2B) indicating that GABAB(1a,2) heteroreceptors at glutamatergic
ter-minals are essential regulators of oscillation strength.
3.2. The auxiliary subunit KCTD16 influences oscillation strength and frequency
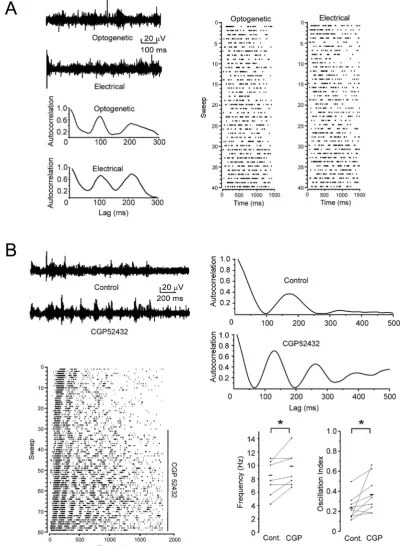
To address a possible contribution of auxiliary KCTD subunits in the regulation of sleep spindles we performed complementary sets of experiments in mice with a genetic lack of KCTD12 or KCTD16. To activate thalamic spindles directly from within the thalamus, without the involvement of corticothalamic fibers, we crossed KCTD12/ andKCTD16/ mice with transgenic mice expressing ChR2 together with an enhanced yellowfluorescent protein (EYFP) under control of the VGAT promoter (Zhao et al., 2011). This allows to elicit spindle oscillations via photostimulation of the TRN in thalamic slices kept in the interface chamber, similarly as previ-ously shown in live mice (Halassa et al., 2011). A comparison of electrically and optically induced spindles under control conditions in the same slice of VGAT-ChR2 mice revealed no significant dif-ference in frequency or oscillation strength (p>0.1, n¼5) (Fig. 3A). Similarly, the variability of both parameters was independent of the stimulation method (F-test, p>0.5). These results support earlier findings suggesting that electrical stimulation of the thalamic pathway predominantly activates TRN cells via cortico-reticular inputs (Destexhe et al., 1998) (Fig. 1A).Fig. 3B shows an example of optogenetically stimulated spindles in VGAT-ChR2-EYFP mice under control conditions and in the presence of CGP52432. The spike raster plot of the entire experiment is dis-played alongside with the ACs under control and CGP52432 con-ditions. Similar to the effects of CGP52432 on electrically evoked spindles in WT mice the oscillation frequency of optogenetically stimulated spindles in VGAT-ChR2-EYFP mice significantly increased from 8.2±0.8 Hz to 9.5±0.8 Hz (mean±s.e.m, n¼9, p<0.05, pairedt-test) in the presence of CGP52432. Likewise, the oscillation strength concomitantly increased from 0.23± 0.04 to 0.37±0.06 (p<0.05). Blocking GABABreceptors with CGP52432 in
KCTD12/ mice increased the oscillation frequency from 8.0±0.64 Hz to 9.6±0.73 Hz (p<0.05, n¼9) and the oscillation strength from 0.2±0.03 to 0.4±0.06 (p<0.05) (Fig. 4A), similarly as in WT mice (Fig. 3B). In contrast, spindle frequency as well as oscillation strength remained unchanged in the presence of CGP52432 inKCTD16/mice (p> 0.1, n¼11) (Fig. 4B). Overall, these results support that thalamic oscillations are predominantly influenced by GABABreceptors incorporating the auxiliary subunit
KCTD16.
Comparison of spindle frequency and strength of the different genotypes under control conditions with electrical or optical stimulation revealed no statistically significant difference in spin-dle frequency (ordinary one-way ANOVA, F2,20 ¼ 0.64 and
Fig. 2. Differential effects of GABAB(1a,2)and GABAB(1b,2)receptors on thalamic spindles.(A, top left) Sample traces of electrically evoked MUR signals in somatosensory thalamus of aGABAB1a/mouse recorded under control conditions and in the presence of CGP52432 (1mM). (A, bottom left) Raster plot of spikes in consecutive sweeps for the entire experiment. CGP52432 application is indicated by a vertical bar. (A, top right) Autocorrelograms of MUR activity under control conditions and in the presence of CGP52432 show a left-shift of the satellite peaks after GABABreceptor blockade. (A, bottom right) Population data (squares) and averages (dashes) of spindle frequency and oscillation strength in control and CGP52432; (**) p<0.01, n¼7. (B, top left) Individual traces of MUR activity in the somatosensory thalamus recorded of aGABAB1b/mouse in control conditions and after blocking GABABreceptors with CGP52432 (1mM). (B, bottom left) Raster plot of spikes in successive trials for the duration of the complete experiment. (B, top right) Auto-correlograms of MUR activity in control and CGP52432 show increased satellite peaks in the presence of CGP52432 compared to control. (B, bottom right) Population data (squares) and averages (dashes) under control conditions and in the presence of CGP52432 reveal an increase of oscillation strength but no change in oscillation frequency with CGP52432. (**) p<0.01, n¼8.
F2,27¼3.34, p>0.05). However, intragroup variability of the
un-paired data sets could have prevented the detection of subtle intergroup differences (data not shown). Furthermore, no statisti-cally significant difference in the variance of either spindle
[image:6.595.90.491.68.615.2](F2,20¼0.45 and F2,27¼0.04, p>0.5).
3.3. Heteroreceptors at intrathalamic synapses are the regulators of spindle oscillation strength
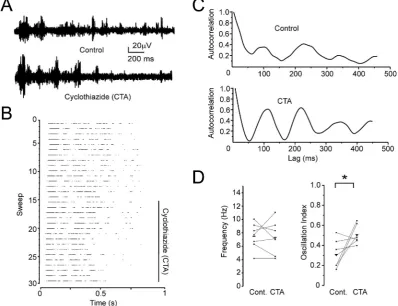
To assess whether increases in thalamoreticular EPSP ampli-tudes, as expected after blocking GABABheteroreceptors, increases
spindle oscillation strength we performed experiments with cyclothiazide (CTA), a positive allosteric modulator of AMPA- and kainate-type glutamate receptors that potentiates glutamate-mediated currents. We activated spindles optogenetically in VGAT-ChR2-EYFP mice to exclude a potentially confounding acti-vation of TCR cells by corticothalamic synapses (Fig. 5A). Adding CTA (20
m
M) to the bath increased spindle synchronicity without changing their frequency (Fig. 5B and C). Overall, CTA increased the spindle oscillation strength from 0.31±0.05 to 0.51±0.03 (n¼7, p<0.05) (Fig. 5D). This result confirms that increasing the size of EPSPs in TRN cells increases spindle oscillation strength.To further substantiate a role for GABAB heteroreceptors and
KCTD16 in controlling thalamic rhythmogenesis at synaptic inputs to TRN cells we performed whole-cell voltage-clamp recordings in visually identified TRN cells of WT,KCTD16/andGABAB1a/mice. TRN
cells were located in a narrow band delineated by the extramedullary lamina and the capsula interna. Fast inhibitory synaptic transmission was blocked by adding the Cl-channel blocker picrotoxin (0.1 mM) to the bath. Action potentials and sIPSCs were abolished by intra-cellularly applying QX314 (5 mM), which blocks Kir3-type Kþ chan-nels, the main effectors of postsynaptic GABABreceptors, as well as
Nav-channels (Nathan et al., 1990). EPSCs were electrically elicited in
the somatosensory thalamus (Fig. 6A) using a paired-pulse paradigm while washing in baclofen (100
m
M) to quantify GABAB-mediatedpresynaptic inhibition (Fig. 6B). The thalamic versus cortical origin of the EPSCs was confirmed by their characteristic strong short-term synaptic depression as revealed by a paired-pulse ratio of less than one, which was however similar in all genotypes (WT: 0.64±0.3, KCTD16/: 0.66 ± 0.04, GABAB1a/: 0.65± 0.04, one-way ANOVA,
p>0.5) (Fig. 6B) (Gentet and Ulrich, 2003). Moreover, the decay time constants of the EPSCs determined from monoexponential fits revealed no significant difference between genotypes (WT: 8.1±0.36 ms,KCTD16/: 8.4±0.32 ms,GABAB1a/: 8.4±0.24 ms,
one-way ANOVA, p>0.5). The GABABreceptor-mediated inhibition of
EPSC amplitudes differed between WT andKCTD16/orGABA
B1a
/ mice (one-way ANOVA, F2,10¼15.1, p<0.01,Fig. 6C). A strong
inhi-bition of the EPSC amplitudes was observed in WT mice in presence of baclofen (49.6± 2.5%, n¼4). The EPSC amplitudes were still significantly reduced in both the KCTD16/ (22.9± 3.16, n ¼5, p<0.01) andGABAB1a/(25.0±5.31%, n¼4, p<0.01) mice, however to
a significantly lesser degree in both mutants when compared to WT mice (Tukey posthoc test, p<0.01) (Fig. 6C). Interestingly, the amount of inhibition inKCTD16/andGABAB1a/was similar (p>0.5). In all
genotypes, no decrease of the EPSCs amplitudes was observed over time in the absence of baclofen (data not shown) excluding EPSC rundown as a confounding factor. Taken together, these results sup-port that KCTD16 significantly influences GABABreceptor-mediated
[image:8.595.94.493.385.691.2]presynaptic inhibition at the TCR-TRN synapse.
4. Discussion
In this study we addressed how GABAB receptor subtypes
contribute to thalamic spindle oscillations. We induced spindle os-cillations electrically or optogenetically in acute thalamic slices comprising reciprocally connected TCR and TRN cells. The contri-bution of GABABreceptors to spindle oscillations was revealed by
antagonizing receptors with CGP52432. In slices of WT mice CGP52432 led to a significant increase in both spindle frequency and oscillation strength. Antagonizing GABAB(1a,2)receptors inGABAB1b/
mice led to an increase in spindle oscillation strength without altering spindle frequency. Using patch-clamp electrophysiology we confirmed that GABAB(1a,2)receptors function as heteroreceptors at
the TCR-to-TRN synapse, as has been observed at excitatory synap-ses in other brain regions (Ulrich and Bettler, 2007; reviewed in Gassmann and Bettler, 2012). A GABAB(1a,2)heteroreceptor function
in the axon terminals of TCR neurons is also supported by earlier experiments in the amygdala (Shaban et al., 2006) and the syn-chronicity enhancing effect of CTA (this study), a positive allosteric modulator of AMPA/kainate receptors. During optogenetically evoked spindles TCR-to-TRN synapses are the only participating excitatory synapses thus rendering the CTA effect synapse specific. It therefore is safe to conclude that GABA(B1a,2)heteroreceptors on TCR
cells are the key regulators of oscillation strength. Antagonizing GABAB(1b,2)receptors inGABAB1a/mice increased spindle oscillation
frequency without changing oscillation strength. GABAB(1b,2)
re-ceptors are mainly responsible for generating sIPSPs at postsynaptic sites. The frequency enhancing function of a blockade of GABAB(1b,2)
receptors in the present study is therefore likely the result of a postsynaptic blockade of GABABreceptors in TCR neurons. The loss
of GABAB(1b,2)-mediated sIPSPs is expected to result in more rapid
rebound bursting, since the remaining GABAA-mediated fast IPSPs
are of shorter duration (Huguenard and Prince, 1994b). Blockade of postsynaptic GABABreceptors on TCR neurons may also decrease the
strength of spindle oscillations (von Krosigk et al., 1993), which we however did not observe in our experiments. This effect on oscilla-tion strength may either be too subtle to be detected or may be masked by CGP52432 effects that enhance synchronicity. For example, CGP52432 may block a GABABreceptor-mediated
upre-gulation of a tonic GABAAcurrent (Connelly et al., 2013), leading to a
slight depolarization of TCR cells and an increasedfiring synchro-nicity and enhancement of oscillation strength (Sohal et al., 2006). While GABAB(1b,2)receptors also contribute to autoreceptors in some
brain regions (Ulrich and Bettler, 2007; Gassmann and Bettler, 2012) it appears unlikely that the inhibition of GABAB(1b,2)autoreceptors
by CGP52432 underlies the frequency enhancing effect on thalamic spindles observed inGABAB1a/mice. For one, a relief of presynaptic
inhibition at TRN-to-TCR synapses would be expected to increase GABA output and, if anything, prolong IPSPs. In addition, autor-eceptors are generally formed in parity by GABAB(1a,2) and
GABAB(1b,2)receptors and the loss of one subtype would be offset by
[image:9.595.141.461.62.360.2]the remaining subtype (Ulrich et al., 2007; Vigot et al., 2006). However, all other comparisons between genotypes of the basal oscillation frequency and strength did not reach significance levels. Experimental variability within the unpaired data sets likely masks subtle differences. The variability could, for example, result from Fig. 6. Loss of KCTD16 or GABAB1areduces baclofen-mediated presynaptic inhibition of glutamate release at the TCR-TRN synapse.(A) Scheme of a connected TCR-TRN cell pair with the experimental stimulation (St.El.) and recording electrodes. (B) Sample recordings of EPSCs before (black) and after (red) baclofen application (100mM) to acute slices of WT,KCTD16/andGABA
B1a
/mice. The extracellular stimulations consisted of two pulses of 0.5 ms duration at 40 Hz repeated every 20 s. Monoexponentials werefitted to the decaying phase of the EPSCs (blue) using the Chebyshev algorithm (R2>0.9 for allfits). (C) Average inhibition of thefirst EPSC in the three genotypes. Squares indicate single data points and the dashes indicate the means. The paired-pulse ratio was similar in all genotypes in the absence and presence of baclofen (paired-pulse ratio without baclofen: WT¼0.64±0.03,KCTD16/¼0.65±0.04,GABA
B1a
/¼0.64±0.04, one-way ANOVA p>0.5; paired-pulse ratio with baclofen: WT¼0.65±0.04,KCTD16/¼0.66±0.03,GABA B1a / ¼0.66±0.04, one-way ANOVA p>0.5). (**) p<0.01, one-way ANOVA with posthoc Tukey-test. WT, n¼4;KCTD16/, n¼5;GABA
B1a /, n¼4.
variable positions of the recording electrodes in different experi-ments, variable stimulation efficacy in different slices, or variabil-ities in slice preparations. However, a potential compensatory upregulation of functional GABABresponses inGABAB1a/andGABAB1b/
mice seems rather unlikely as a previous study found no exper-imental evidence for a functional compensation in subtype specific knock-out mice (Vigot et al., 2006).
An earlier report already found distinct effects of GABAB(1a,2)and
GABAB(1b,2)receptors on neural network activity in the entorhinal
cortex (Craig et al., 2013). The results showed that activation of postsynaptic GABAB(1b,2)receptors initiates down-state transitions,
consistent with a function of these receptors in hyperpolarizing neurons by activating Kþ efflux. Conversely, activation of GABAB(1a,2)heteroreceptors was shown to limit the duration of the
up-states, in agreement with their role in inhibiting glutamate release.
We further addressed whether the kinetic effects of auxiliary KCTD subunits on the GABABreceptor response (Turecek et al.,
2014; Fritzius et al., 2017) influence thalamic spindle oscillations. Antagonizing GABABreceptors inKCTD12/mice led to a signifi
-cant increase in spindle frequency and oscillation strength, simi-larly as observed in WT mice. KCTD12 therefore does not appear to influence spindle oscillations in a detectable manner. In contrast, no effects of a GABABreceptor blockade on spindle oscillations were
observed inKCTD16/mice, implying that KCTD16 influences both oscillation strength and frequency. Likely therefore, KCTD16 asso-ciates with GABAB(1a,2) heteroreceptors as well as postsynaptic
GABAB(1b,2) receptors in TCR cells to regulate oscillation strength
and frequency, respectively. The absence of a synchronicity enhancing effect of CGP52432 inKCTD16/mice may be due to a compromised coupling of GABABreceptors to N-type Ca2þchannels
(Schwenk et al., 2016) and/or to a reduced agonist affinity at the receptor (Rajalu et al., 2015). Inactivation of N-type Ca2þchannels, presumably at presynaptic sites, was shown to reduce excitatory synaptic transmission between thalamic neurons (Pfrieger et al., 1992) and a reduced affinity for GABA may lower activation of GABAB receptors. Thus, KCTD16 appears to facilitate GABAB(1a,2)
heteroreceptor function, explaining why CGP52432 effects were observed in WT and GABAB1b/ but not in KCTD16/ mice. The
absence of an effect on oscillation frequency of CGP52432 in KCTD16/ mice may be explained by the faster decay of GABAB(1b,2)-mediated sIPSCs than in WT mice (Fritzius et al., 2017),
which reduces the contribution of sIPSCs to rebound burst-firing. Uncontrolled release of glutamate in the thalamus may induce hypersynchronicity and eventually epilepsy because of the strong excitatory drive exerted by TCR cells onto TRN neurons (Gentet and Ulrich, 2003). In support of this notion,GABAB1a/mice were found to
be prone to seizures whileGABAB1b/and WT littermate mice were
seizure resistant (Vienne et al., 2010). Furthermore, a genetic link-age study associates a KCTD16 polymorphism with epilepsy in humans (Angelicheva et al., 2009). Thus GABA(B1a,2)
hetero-receptors containing the auxiliary subunit KCTD16 may be key for regulating thalamic oscillations and their selective upregulation would be beneficial in the therapy of absence seizures. Already in 2003 Norman Bowery and collaborators found decreased GABAB1
transcript levels in the somatosensory thalamus of GAERS (Genetic Absence Epilepsy Rat of Strasbourg), supporting that a GABAB
heteroreceptor hypofunction underlies thalamic hyperactivity (Princivalle et al., 2003).
Acknowledgments
This study was supported by the Swiss National Science Foun-dation (Grant 31003A-172881 to BB).
References
Aghajanian, G.K., Rasmussen, K., 1989. Intracellular studies in the facial nucleus illustrating a simple new method for obtaining viable motoneurons in adult rat
brain slices. Synapse 3, 331e338.
Angelicheva, D., Tournev, I., Guergueltcheva, V., Mihaylova, V., Azmanov, D.N., Morar, B., Radionova, M., Smith, S.J., Zlatareva, D., Stevens, J.M., Kaneva, R., Bojinova, V., Carter, K., Brown, M., Jablensky, A., Kalaydjieva, L., Sander, J.W., 2009. Partial epilepsy syndrome in a Gypsy family linked to 5q31.3-q32.
Epi-lepsia 50, 1679e1688.
Bal, T., von Krosigk, M., McCormick, D.A., 1995. Synaptic and membrane mecha-nisms underlying synchronized oscillations in the ferret lateral geniculate
nu-cleus in vitro. J. Physiol. 483, 641e663.
Booker, S.A., Althof, D., Gross, A., Loreth, D., Müller, J., Unger, A., Fakler, B., Varro, A.,
Watanabe, M., Gassmann, M., Bettler, B., Shigemoto, R., Vida, I., Kulik,A., 2017.
KCTD12 auxiliary proteins modulate kinetics of GABABreceptor-mediated
in-hibition in cholecystokinin-containing interneurons. Cereb. Cortex 27,
2318e2334.
Bowery, N.G., Hill, D.R., Hudson, A.L., Doble, A., Middlemiss, D.N., Shaw, J., Turnbull, M., 1980. (-)Baclofen decreases neurotransmitter release in the
mammalian CNS by an action at a novel GABA receptor. Nature 283, 92e94.
Bowery, N.G., Hudson, A.L., Price, G.W., 1987. GABAAand GABABreceptor site
dis-tribution in the rat central nervous system. Neuroscience 20, 365e383.
Cathomas, F., Stegen, M., Sigrist, H., Schmid, L., Seifritz, E., Gassmann, M., Bettler, B., Pryce, C.R., 2015. Altered emotionality and neuronal excitability in mice lacking
KCTD12, an auxiliary subunit of GABABreceptors associated with mood
disor-ders. Transl. Psychiatry 5, e510.
Cathomas, F., Sigrist, H., Schmid, L., Seifritz, E., Gassmann, M., Bettler, B., Pryce, C.R.,
2017. Behavioural endophenotypes in mice lacking the auxiliary GABABreceptor
subunit KCTD16. Behav. Brain Res. 317, 393e400.
Connelly, W.M., Fyson, S.J., Errington, A.C., McCafferty, C.P., Cope, D.W., Di
Giovanni, G., Crunelli, V., 2013. GABABreceptors regulate extrasynaptic GABAA
receptors. J. Neurosci. 33, 3780e3785.
Craig, M.T., McBain, C.J., 2013. The emerging role of GABABreceptors as regulators of
network dynamics: fast actions from a 'slow' receptor? Curr. Opin. Neurobiol.
26, 15e21.
Craig, M.T., Mayne, E.W., Bettler, B., Paulsen, O., McBain, C.J., 2013. Distinct roles of
GABAB1a- and GABAB1b-containing GABABreceptors in spontaneous and evoked
termination of persistent cortical activity. J. Physiol. 591, 835e843.
Destexhe, A., Contreras, D., Steriade, M., 1998. Mechanisms underlying the syn-chronizing action of corticothalamic feedback through inhibition of thalamic
relay cells. J. Neurophysiol. 79, 999e1016.
Deboer, T., Tobler, I., 1995. Temperature dependence of EEG frequencies during
natural hypothermia. Brain Res. 670, 153e156.
Fritzius, T., Turecek, R., Seddik, R., Kobayashi, H., Tiao, J., Rem, P.D., Metz, M., Kralikova, M., Bouvier, M., Gassmann, M., Bettler, B., 2017. KCTD
hetero-oligomers confer unique kinetic properties on hippocampal GABAB
receptor-induced Kþ-currents. J. Neurosci. 37, 1162e1175.
Gassmann, M., Bettler, B., 2012. Regulation of neuronal GABABreceptor functions by
subunit composition. Nat. Rev. Neurosci. 13, 380e394.
Gentet, L.J., Ulrich, D., 2003. Strong, reliable and precise synaptic connections be-tween thalamic relay cells and neurones of the nucleus reticularis in juvenile
rats. J. Physiol. 546, 801e811.
Halassa, M.M., Siegle, J.H., Ritt, J.T., Ting, J.T., Feng, G., Moore, C.I., 2011. Selective optical drive of thalamic reticular nucleus generates thalamic bursts and
cortical spindles. Nat. Neurosci. 14, 1118e1120.
Hill, D.R., Bowery, N.G., 1981. 3H-baclofen and 3H-GABA bind to
bicuculline-insensitive GABABsites in rat brain. Nature 290, 149e152.
Hill, D.R., Bowery, N.G., Hudson, A.L., 1984. Inhibition of GABABreceptor binding by
guanyl nucleotides. J. Neurochem. 42, 652e657.
Hosford, D.A., Clark, S., Cao, Z., Wilson Jr., W.A., Lin, F.H., Morrisett, R.A., Huin, A.,
1992. The role of GABABreceptor activation in absence seizures of lethargic (lh/
lh) mice. Science 257, 398e401.
Huguenard, J.R., Prince, D.A., 1994a. Clonazepam suppresses GABAB-mediated
in-hibition in thalamic relay neurons through effects in nucleus reticularis.
J. Neurophysiol. 71, 2576e2581.
Huguenard, J.R., Prince, D.A., 1994b. Intrathalamic rhythmicity studied in vitro: nominal T-current modulation causes robust antioscillatory effects. J. Neurosci.
14, 5485e5502.
Jacobsen, R.B., Ulrich, D., Huguenard, J.R., 2001. GABAB and NMDA receptors
contribute to spindle-like oscillations in rat thalamus in vitro. J. Neurophysiol.
86, 1365e1375.
Kaupmann, K., Huggel, K., Heid, J., Flor, P.J., Bischoff, S., Mickel, S.J., McMaster, G.,
Angst, C., Bittiger, H., Froestl, W., Bettler, B., 1997. Expression cloning of GABAB
receptors uncovers similarity to metabotropic glutamate receptors. Nature 386,
239e246.
Kaupmann, K., Malitschek, B., Schuler, V., Heid, J., Froestl, W., Beck, P., Mosbacher, J.,
Bischoff, S., Kulik, A., Shigemoto, R., Karschin, A., Bettler, B., 1998. GABAB
-re-ceptor subtypes assemble into functional heteromeric complexes. Nature 396,
683e687.
Kleiman-Weiner, M., Beenhakker, M.P., Segal, W.A., Huguenard, J.R., 2009.
Syner-gistic roles of GABAAreceptors and SKchannels in regulating thalamocortical
oscillations. J. Neurophysiol. 102, 203e213.
activity. Adv. Pharmacol. 58, 205e229.
McCormick, D.A., Bal, T., 1997. Sleep and arousal: thalamocortical mechanisms.
Annu. Rev. Neurosci. 20, 185e215.
Metz, M., Gassmann, M., Fakler, B., Schaeren-Wiemers, N., Bettler, B., 2011.
Distri-bution of the auxiliary GABABreceptor subunits KCTD8, 12, 12b, and 16 in the
mouse brain. J. Comp. Neurol. 519, 1435e1454.
Nathan, T., Jensen, M.S., Lambert, J.D., 1990. The slow inhibitory postsynaptic po-tential in rat hippocampal CA1 neurones is blocked by intracellular injection of
QX-314. Neurosci. Lett. 110, 309e313.
Pfrieger, F.W., Veselovsky, N.S., Gottmann, K., Lux, H.D., 1992. Pharmacological characterization of calcium currents and synaptic transmission between
thalamic neurons in vitro. J. Neurosci. 12, 4347e4357.
Pin, J.P., Bettler, B., 2016. Organization and functions of mGlu and GABABreceptor
complexes. Nature 540, 60e68.
Pinault, D., Leresche, N., Charpier, S., Deniau, J.M., Marescaux, C., Vergnes, M., Crunelli, V., 1998. Intracellular recordings in thalamic neurones during spon-taneous spike and wave discharges in rats with absence epilepsy. J. Physiol. 509,
449e456.
Porter, J.T., Nieves, D., 2004. Presynaptic GABABreceptors modulate thalamic
exci-tation of inhibitory and excitatory neurons in the mouse barrel cortex.
J. Neurophysiol. 92, 2762e2770.
Princivalle, A.P., Richards, D.A., Duncan, J.S., Spreafico, R., Bowery, N.G., 2003.
Modification of GABAB1and GABAB2receptor subunits in the somatosensory
cerebral cortex and thalamus of rats with absence seizures (GAERS). Epilepsy
Res. 55, 39e51.
Rajalu, M., Fritzius, T., Adelfinger, L., Jacquier, V., Besseyrias, V., Gassmann, M.,
Bettler, B., 2015. Pharmacological characterization of GABABreceptor subtypes
assembled with auxiliary KCTD subunits. Neuropharmacology 88, 145e154.
Schwenk, J., Perez-Garci, E., Schneider, A., Kollewe, A., Gauthier-Kemper, A.,
Fritzius, T., Raveh, A., Dinamarca, M.C., Hanuschkin, A., Bildl, W., Klingauf, J., Gassmann, M., Schulte, U., Bettler, B., Fakler, B., 2016. Modular composition and
dynamics of native GABABreceptors identified by high-resolution proteomics.
Nat. Neurosci. 19, 233e242.
Schwenk, J., Metz, M., Zolles, G., Turecek, R., Fritzius, T., Bildl, W., Tarusawa, E., Kulik, A., Unger, A., Ivankova, K., Seddik, R., Tiao, J.Y., Rajalu, M., Trojanova, J.,
Rohde, V., Gassmann, M., Schulte, U., Fakler, B., Bettler, B., 2010. Native GABAB
receptors are heteromultimers with a family of auxiliary subunits. Nature 465,
231e235.
Shaban, H., Humeau, Y., Herry, C., Cassasus, G., Shigemoto, R., Ciocchi, S., Barbieri, S., van der Putten, H., Kaupmann, K., Bettler, B., Lüthi, A., 2006. Generalization of amygdala LTP and conditioned fear in the absence of presynaptic inhibition.
Nat. Neurosci. 9, 1028e1035.
Sohal, V.S., Pangratz-Fuehrer, S., Rudolph, U., Huguenard, J.R., 2006. Intrinsic and synaptic dynamics interact to generate emergent patterns of rhythmic bursting
in thalamocortical neurons. J. Neurosci. 26, 4247e4255.
Soltesz, I., Lightowler, S., Leresche, N., Crunelli, V., 1989. On the properties and origin
of the GABAB inhibitory postsynaptic potential recorded in morphologically
identified projection cells of the cat dorsal lateral geniculate nucleus.
Neuro-science 33, 23e33.
Staak, R., Pape, H.C., 2001. Contribution of GABAAand GABABreceptors to thalamic
neuronal activity during spontaneous absence seizures in rats. J. Neurosci. 21,
1378e1384.
Steriade, M., Desch^enes, M., 1984. The thalamus as a neuronal oscillator. Brain Res.
320, 1e63.
Steriade, M., Llinas, R.R., 1988. The functional states of the thalamus and the
associated neuronal interplay. Physiol. Rev. 68, 649e742.
Steriade, M., McCormick, D.A., Sejnowski, T.J., 1993. Thalamocortical oscillations in
the sleeping and aroused brain. Science 262, 679e685.
Stuart, G.J., Dodt, H.U., Sakmann, B., 1993. Patch-clamp recordings from the soma and dendrites of neurons in brain slices using infrared video microscopy.
Pflügers Arch. 423, 511e518.
Turecek, R., Schwenk, J., Fritzius, T., Ivankova, K., Zolles, G., Adelfinger, L., Jacquier, V., Besseyrias, V., Gassmann, M., Schulte, U., Fakler, B., Bettler, B., 2014. Auxiliary
GABABreceptor subunits uncouple G proteinbgsubunits from effector channels
to induce desensitization. Neuron 82, 1032e1044.
Ulrich, D., Besseyrias, V., Bettler, B., 2007. Functional mapping of GABAB-receptor
subtypes in the thalamus. J. Neurophysiol. 98, 3791e3795.
Ulrich, D., Bettler, B., 2007. GABABreceptors: synaptic functions and mechanisms of
diversity. Curr. Opin. Neurobiol. 17, 298e303.
Ulrich, D., Huguenard, J.R., 1995. Purinergic inhibition of GABA and glutamate release in the thalamus: implications for thalamic network activity. Neuron 15,
909e918.
Ulrich, D., Huguenard, J.R., 1996. GABABreceptor-mediated responses in GABAergic
projection neurones of rat nucleus reticularis thalami in vitro. J. Physiol. 493,
845e854.
Vienne, J., Bettler, B., Franken, P., Tafti, M., 2010. Differential effects of GABAB
re-ceptor subtypes,g-hydroxybutyric acid, and baclofen on EEG activity and sleep
regulation. J. Neurosci. 30, 14194e14204.
Vigot, R., Barbieri, S., Br€auner-Osborne, H., Turecek, R., Shigemoto, R., Zhang, Y.P.,
Lujan, R., Jacobson, L.H., Biermann, B., Fritschy, J.M., Vacher, C.M., Müller, M.,
Sansig, G., Guetg, N., Cryan, J.F., Kaupmann, K., Gassmann, M., Oertner, T.G., Bettler, B., 2006. Differential compartmentalization and distinct functions of
GABABreceptor variants. Neuron 50, 589e601.
von Krosigk, M., Bal, T., McCormick, D.A., 1993. Cellular mechanisms of a
synchro-nized oscillation in the thalamus. Science 261, 361e364.
White, J.H., Wise, A., Main, M.J., Green, A., Fraser, N.J., Disney, G.H., Barnes, A.A., Emson, P., Foord, S.M., Marshall, F.H., 1998. Heterodimerization is required for
the formation of a functional GABABreceptor. Nature 396, 679e682.
Zhao, S., Ting, J.T., Atallah, H.E., Qiu, L., Tan, J., Gloss, B., Augustine, G.J., Deisseroth, K.,
Luo, M., Graybiel, A.M., Feng, G., 2011. Cell typeespecific channelrhodopsin-2
transgenic mice for optogenetic dissection of neural circuitry function. Nat.
Methods 8, 745e752.