Associative learning is thought to play an important role in human learning and is brought about by either classical or operant conditioning procedures (Kimble, 1961). Unlike classical conditioning procedures, the presentation of a stimulus using operant conditioning is response-contingent. Following operant conditioning, an association between a behavioural response and a particular reinforcing stimulus is learned, resulting in specific behavioural changes.
Invertebrate preparations have been widely used to study the effect of both classical and operant conditioning procedures on various behaviour patterns (Carew and Sahley, 1986; Mpitsos and Lukowiak, 1985). An important element of many of these studies has been the demonstration of long-term memory of the learned behaviour. For example, in gastropod molluscs such as Aplysia californica, Hermissenda crassicornis and Limax maximus, both non-associative and associative training procedures produced memories that persisted for a number of weeks (Carew et al. 1972; Castellucci et al. 1989; Alkon, 1984; Gelperin, 1975, respectively). Furthermore, learning in Lymnaea stagnalis following classical conditioning procedures
has been shown to persist for days to weeks (Alexander et al. 1982; Kemenes and Benjamin, 1994; Kojima et al. 1997). Previously, however, long-term memory following associative learning in molluscs has been demonstrated only after classical conditioning procedures. Whether operant conditioning procedures in molluscan systems are also able to induce long-term memories has not yet been demonstrated. In the present study, we utilized the respiratory behaviour of the pond snail Lymnaea stagnalis to determine whether operant conditioning would result in a long-term memory.
Lymnaea stagnalis breathes bimodally either through its skin or via a rudimentary lung (aerial or pulmonary respiration; Jones, 1961). We have previously demonstrated that aerial respiratory behaviour in L. stagnalis can be operantly conditioned (Lukowiak et al. 1996a) in a hypoxic environment where pulmonary respiratory behaviour should predominate (Lukowiak et al. 1996a). The operant conditioning procedure consisted of presenting a tactile stimulus (using a sharpened wooden applicator stick) to the pneumostome area (the respiratory orifice) every time the snail attempted to open its JEB1347
The freshwater snail Lymnaea stagnalis breaths bimodally either through its skin (cutaneous respiration) or via a rudimentary lung opening called the pneumostome (aerial respiration). Aerial respiratory behaviour can be operantly conditioned. Animals placed in an aquatic, hypoxic environment received a tactile stimulus to the pneumostome area every time they attempted to breathe. Over a period of five training sessions (2.5 days), the animals learned not to breathe, and the number of stimuli received in the fifth session was significantly lower than in the first session. These changes in the respiratory behaviour following the operant paradigm were shown to persist for at least 24 h. We aimed to determine whether the changes in the learned behaviour would persist for longer. We obtained direct evidence that the behavioural changes following operant conditioning persisted for at least 4 weeks following the last training session. However, we
found that the persistence of this memory was dependent upon the training procedure used. Memory persisted longer following a spaced training procedure (4 weeks) as opposed to a massed training procedure (2 weeks). Yoked control animals showed no changes in their respiratory behaviour over the same time periods. However, if these yoked control animals were subjected to an operant conditioning procedure, their ability to learn was not impeded. This study demonstrated that operant conditioning of a behaviour pattern in a molluscan preparation can result in long-term memory and that the persistence of the memory is contingent on the training procedure used.
Key words: Lymnaea stagnalis, mollusc, snail, operant conditioning, long-term memory, learning.
Summary
Introduction
LONG-TERM MEMORY OF AN OPERANTLY CONDITIONED RESPIRATORY
BEHAVIOUR PATTERN IN LYMNAEA STAGNALIS
KEN LUKOWIAK*, RALPH COTTER, JACKIE WESTLY, ERIKA RINGSEIS, GAYNOR SPENCER AND
NAWEED SYED
Neuroscience and Respiratory Research Groups, Faculty of Medicine, University of Calgary, 3330 Hospital Drive NW, Calgary, Alberta, Canada T2N 4N1
*Present address: Department of Physiology, Arabian Gulf University/CMMS, Manama, The State of Bahrain (e-mail: [email protected])
pneumostome to breathe. As a result of this procedure, the animals learned to suppress their aerial respiration even though they were tested in a hypoxic environment. We demonstrated that these behavioural changes persisted for up to 24 h, but no attempts were made to determine whether this memory could persist for longer periods. In the present study, we aimed to determine how long the behavioural changes produced by the operant conditioning procedure would endure and whether memory retention was dependent upon either the number or the spacing of the training sessions.
A preliminary report of these data has appeared in abstract form (Lukowiak et al. 1996b).
Materials and methods
Lymnaea stagnalis (laboratory-bred, but originally derived
from Vrije University, Amsterdam) of a shell length 25–30 mm were used in these studies. The operant conditioning procedure was similar to that used previously (Lukowiak et al. 1996a). Briefly, each snail was individually identified by a unique set of coloured pen marks on its shell and placed in 800 ml of hypoxic pond water contained in a glass beaker (2000 ml). Following a 20 min acclimation period, during which the animals were allowed to breathe freely, the snails entered a 30 min training period (or operant training period). During this period, every time a snail attempted to open its pneumostome to perform pulmonary respiration, a tactile stimulus was presented to the pneumostome area, and the number and time of the stimuli were recorded. The tactile stimulus was delivered to the pneumostome area by the observer using a sharpened wooden ‘Thai-satay’ stick. Following this training period, the animals were given a further 20 min during which no tactile stimuli were presented. Thus, snails were trained and their memory accessed in a hypoxic environment. The animals were eventually returned to the aquarium (normoxic condition) until the next training session (for details, see Lukowiak et al. 1996a).
Training procedures
Three different training procedures were used to test for long-term memory of the learned response. The initial procedure used (procedure 1) was similar to that used in the previous study (Lukowiak et al. 1996a). Briefly, the animals received five training sessions of 0.5 h duration, with two sessions per day, over a period of 2.5 days. In the present study, following the last training session (the fifth), separate groups of animals were then tested for retention of memory 1 or 2 weeks following the last training session.
In procedure 2, also termed the massed training procedure (MTP), each snail received two 0.5 h training sessions per day for 4 days. Thus, these snails received eight training sessions, over a 4 day period. Separate cohorts (N=10) of these MTP snails were then retested for retention of the memory 1, 2 or 4 weeks later.
In procedure 3, termed the spaced training procedure (STP), the animals also received eight training sessions, but these
sessions were spread over 58.5 days instead of 4 days (as in the MTP groups). The STP animals received the initial five training sessions over 2.5 days, as in procedure 1. The STP snails then received further training sessions after 1 week, 2 weeks and 4 weeks, and were finally retested for retention of their memories after a further 4 weeks.
In summary, in procedure 1, each snail received only five training sessions prior to the final test, whereas in both the MTP and STP groups, each snail received eight training sessions prior to the final test. In the MTP groups, the animals received their training sessions over a period of 4 days, whereas in the STP group, the animals received their training over a period of 58.5 days.
A similar number of animals was also subjected to a yoked control procedure for each of the above training procedures. These yoked control animals received the same number of tactile stimuli to the pneumostome area, but these stimuli were not contingent upon the pneumostome openings in the yoked animals, but rather on the pneumostome openings of the snail in the operant group to which it was yoked (see Lukowiak et
al. 1996a). These animals were also tested at the same time
intervals as the operant animals and, when tested, received a tactile stimulus to the pneumostome area each time the animal attempted to breathe.
Statistical analysis consisted of an analysis of variance (ANOVA) followed by the appropriate post-hoc test (Fisher’s LSD protected t-test). Differences were deemed to be significant if P<0.05.
Results
Procedure 1 animals
We first determined whether the memory persisted for 1 week following the five training sessions given over a 2.5 day period (Fig. 1A; N=12). An analysis of variance (ANOVA) showed that there was a significant effect of training on the number of tactile stimuli received by the snails during the training procedure (F=11.39, P<0.001). The data obtained on session 6 (1 week following the last training session) were not statistically different from those obtained on sessions 5 (P>0.05). That is, the animals retained the learned behaviour for this 1 week interval. However, when a separate cohort of snails (N=10) was tested in a similar manner, but following a 2 week period between session 5 and 6, there was no persistence of the memory (Fig. 1B). That is, the number of responses on session 6 was significantly different from that on session 5 (P<0.05), but was not significantly different from that obtained in session 1 (P>0.05). From these data, we concluded that a 2.5 day training regime was sufficient to produce a memory that persisted for at least 1 week following the last training session, but which did not persist for 2 weeks.
Procedure 2 animals (MTP)
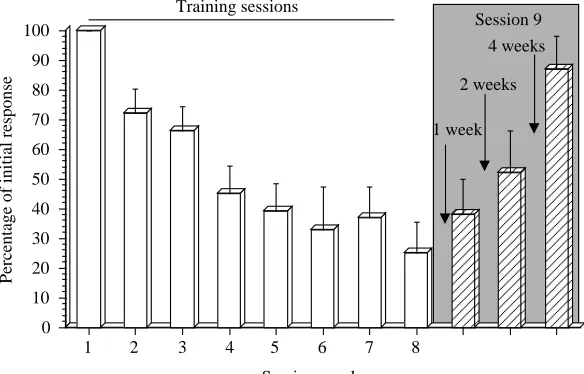
the last training session. The MTP snails retained their memory of the learned behaviour for both week 1 and week 2. An ANOVA performed on each specific cohort of animals (N=10 for each cohort) revealed a significant effect of the operant conditioning procedure (Fig. 2; ANOVA F=10.9871, P<0.001 for tests after 1 week; ANOVA F=5.0125, P<0.001 for tests after 2 weeks). The numbers of stimuli received on session 9 in the week 1 and 2 cohorts were not significantly different from those on session 8 (P>0.05 for both groups), but were significantly different from the response seen in session 1 (P<0.01 for both groups). The MTP snails did not, however, retain the memory of the learned behaviour when tested at week 4; that is, the response on session 9 was significantly different from that on session 8 (P<0.01) but was not significantly different from that on session 1 (P>0.05; Fig. 2). Yoked control snails subjected to a similar stimulation protocol did not exhibit any learning when tested at week 1, week 2 or week 3 (P>0.05 in all cases; data not shown).
Procedure 3 animals (STP)
As with the MTP snails, the STP (N=40) snails also received eight training sessions. These training sessions were, however, spread over a 58.5 day period. The initial five training sessions were received over a 2.5 day period, and the remaining training sessions were given 1, 2 and 4 weeks later (sessions 6, 7 and 8, respectively). Each animal was then tested 4 weeks later (week 8; session 9). An ANOVA showed a significant effect of training on the number of tactile stimuli delivered to the STP animals (F=19.92, P<0.001; Fig. 3). These animals retained their memory on session 6 (1 week). That is, the number of tactile stimuli delivered to their pneumostome area was significantly smaller (P<0.01) than that on session 1, but was not significantly different from the number of stimuli they received on session 5 (P>0.05). In addition, they continued to exhibit a similar retention of this memory when tested at week 2 (session 7, P<0.01, comparison of session 7 with session 1;
P>0.05, comparison of session 7 with session 6) and at week
0 1 2 3 4 5 6 7
1 week
1 2 3 4 5 6
A
Number of attempted pneumostome openings
0 1 2 3 4 5 6 7
Session number
2 weeks
1 2 3 4 5 6
[image:3.609.59.295.73.372.2]B
Fig. 1. Utilization of a 2.5 day training procedure (procedure 1) results in associative learning and its persistence for 1 week but not 2 weeks. (A) The mean number (+S.E.M.) of attempted pneumostome openings (N=12) over the five training sessions and after a 1 week interruption have been plotted. The number of attempted openings in session 6 was not significantly different from that in session 5, but was significantly different from that in session 1. (B) As in A, but with a 2 week interruption between sessions 5 and 6. In this case, however, the number of attempted openings in session 6 was significantly different from that in session 5, but was not significantly different from that in session 1 (N=10).
0 10 20 30 40 50 60 70 80 90 100
Percentage of initial response
Session number Training sessions
1 week 2 weeks
4 weeks Session 9
1 2 3 4 5 6 7 8
Fig. 2. The massed training procedure (MTP) results in associative learning that persists for 2 but not 4 weeks. The mean numbers (+S.E.M.) of attempted
[image:3.609.272.564.548.735.2]4 (session 8, P>0.05, comparison of session 8 with session 7 and of session 8 with session 6; P<0.01, comparison of session 8 with session 1). Finally, when retested following the 4 week interval, they still demonstrated a memory of the learned behaviour (P>0.05, comparison of session 9 with session 8, and of session 9 with sessions 7, 6 and 5; P<0.01, comparison of session 9 with session 1).
Procedure 3 yoked control animals
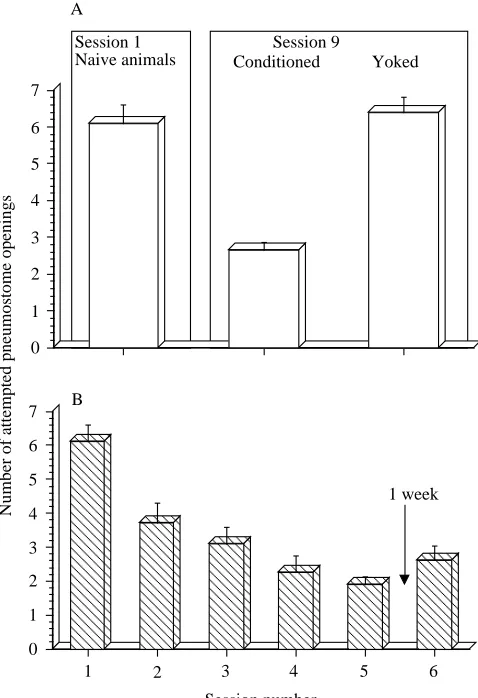
The yoked control animals, despite having received the same stimulation pattern over the same period, did not exhibit any learned behaviour. That is, the number of stimuli they received on session 9 was not significantly different from that received by naive snails on session 1 (Fig. 4A; P<0.05). Moreover, when these animals were subsequently used in the operant training procedure (Fig. 4B), they exhibited a similar decrease in the number of stimuli over the course of the 2.5 day training period as did naive animals. These data showed that subjecting the animals to a long-term yoked procedure did not result in any subsequent impairment of their ability to be operantly conditioned.
Comparison of MTP with STP
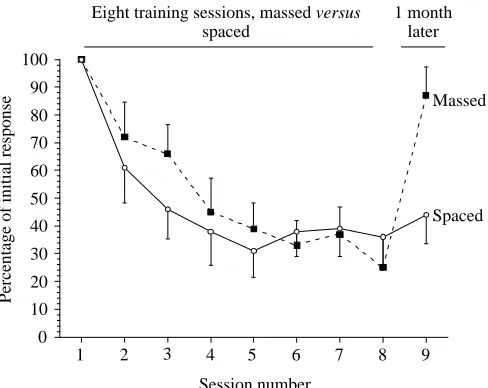
In Fig. 5, the data from the MTP and STP groups have been
normalized and plotted together. The two different training procedures result in similar learning curves, which are indistinguishable from each other up to session 8. That is, each procedure results in a similar level of learning. However, the persistence of the learning was markedly different when tested 1 month following the last training session (comparison of the responses on session 9). Thus, the longer persistence of the memory in the STP group is not due to the achievement of a better level of learning by the STP group.
0 1 2 3 4 5 6 7
Number of attempted pneumostome
openings
Session number 1 week
1 week 2 weeks
4 weeks
[image:4.609.280.559.76.238.2]1 2 3 4 5 6 7 8 9
Fig. 3. The spaced training procedure (STP) results in associative learning that persists for at least 4 weeks. The mean numbers (+S.E.M.) of attempted pneumostome openings in the nine sessions have been plotted. Snails (N=40) were initially trained as for procedure 1, five sessions over a 2.5 day period followed by additional training so that they ultimately received eight training sessions over a 58.5 period. When tested after a further 4 week interval (session 9), the number of attempted pneumostome openings was not significantly different from those in sessions 5, 6, 7 or 8; however, it was significantly different from that in session 1.
Fig. 4. Animals yoked to STP animals did not show any learning, but could be subsequently trained. (A) The mean number (+S.E.M.) of
attempted pneumostome openings observed in session 1 for the snails in the STP group are plotted (N=40). These animals have been labelled in this graph as naive animals since this was their first training session. The yoked controls received the same number of tactile stimuli to their pneumostome area as did the snails in the STP group. When the STP snails were tested in session 9, there was a significant reduction in the number of attempted pneumostome openings (mean + S.E.M.). However, when the yoked animals were
tested following the 4 week interval, they performed (mean + S.E.M.) as would be expected of naive animals; that is, they did not shown any learning. (B) When tested using procedure 1, these animals exhibited learning similar to that expected of naive animals. The mean (+S.E.M.) numbers of attempted pneumostome openings on each session have been plotted.
0 1 2 3 4 5 6 7
Number of attempted pneumostome openings
Session 9 Session 1
Naive animals Conditioned Yoked A
0 1 2 3 4 5 6 7
Session number
1 week
1 2 3 4 5 6
[image:4.609.317.556.387.736.2]Discussion
In the present study, we demonstrated that aerial respiratory behaviour of L. stagnalis can be operantly conditioned and that the memory of this associatively learned event can persist for up to 4 weeks. Operant conditioning paradigms have not been extensively employed in molluscan preparations (for a review of this literature, see Lukowiak et al. 1996a). The most direct comparison we can make from our data, therefore, is with long-term memory induced by classical conditioning procedures. For example, using L. stagnalis (obtained from the same Amsterdam stock), Kojima et al. (1997) demonstrated that an aversive classical conditioning procedure induced an associative behavioural change that persisted for 1 month. Alexander et al. (1982) used a non-aversive classical conditioning procedure which induced learning in wild L.
stagnalis that persisted for 11 days. Similarly, an appetitive
classical conditioning procedure in wild L. stagnalis resulted in a memory that persisted for up to 12 days (Kemenes and Benjamin, 1994). In other molluscan studies (Carew et al. 1972; Castellucci et al. 1989; Alkon, 1984; Gelperin, 1975), memory retention following either non-associative classical conditioning or one-session learning procedures was found to last for at least 2 weeks. Consistent with these previous studies, we have shown that the memory of an operantly conditioned behaviour in a molluscan preparation can persist for long periods.
We showed that the persistence of the learned behaviour
following operant conditioning in L. stagnalis was dependent upon the training procedure used. Our initial training procedure (procedure 1) resulted in a memory of the operantly conditioned behaviour that persisted for 1 week but not for 2 weeks. When a greater number of training sessions (eight
versus five) was employed in the MTP group, however,
memory retention persisted for 2 weeks. Finally, we showed that employing the same number of training sessions as in the MTP group, but with spacing (the spaced training procedure) over a period of almost 60 days rather than 4 days, a memory was induced that persisted for at least 4 weeks. We have not yet attempted to determine the shortest training period required to produce a memory that persisted for such a period.
The finding that spaced training is more effective in producing longer memory retention than massed training has previously been shown in molluscs for both non-associative and associative (classical conditioning) learning procedures. For example, spaced training was more effective in producing long-term retention of habituation of the Aplysia californica gill-withdrawal response than was massed training (Carew et
al. 1972). A similar result using an analogue of a
non-associative learning procedure was obtained in reconstructed neuronal circuits, which underlie some aspects of the siphon withdrawal response in A. californica (Kandel, 1989). Furthermore, Alexander et al. (1982) demonstrated that spaced training (15 sessions over 3 days) versus massed training (15 sessions on one day) resulted in a longer persistence of memory in L. stagnalis (11 versus 3 days). Our demonstration, in this present study, that a spaced training procedure was more effective in producing long-term memory in L. stagnalis was thus not totally unexpected. However, it is the first instance that we know of in a molluscan preparation where an operant conditioning procedure was employed to determine the duration of a long-term memory.
Another interesting and somewhat unexpected finding that arose from these studies was that yoked animals subjected to the long-term stimulation procedure did not exhibit any apparent impairment of their ability to be subsequently associatively conditioned. Previous studies on other preparations, including molluscs, have shown that yoked animals have a tendency to acquire a conditioned response significantly more slowly than do naive animals (Mackintosh, 1974; Cook and Carew, 1989). It was suggested that the reason for this was that the reinforcing stimulus experienced by the yoked animal during the time it was in the control group made it more difficult for the association to be made between the occurrence of the behaviour and the contingent presentation of the reinforcing stimulus. A possible explanation for our finding may be that the 4 week interval was enough time for the ‘memory’ of the yoked procedure to fade from the animals. Thus, they acted just as naive animals would. Further experimentation will be required to determine whether this is indeed the explanation. For example, it may be that, following only a 1 week interval, the yoked animals might acquire the associative response at a significantly slower rate.
The central pattern generator (CPG), sensory and motor
Massed
0 10 20 30 40 50 60 70 80 90 100
Percentage of initial response
Session number
Spaced 1 month
later Eight training sessions, massed versus
spaced
[image:5.609.53.297.72.266.2]1 2 3 4 5 6 7 8 9
neurones that mediate aerial respiration in L. stagnalis have previously been identified, and their synaptic interactions are well characterized (Syed et al. 1990, 1992; Inoue et al. 1996a). Preliminary electrophysiological studies have also indicated that changes in these identified neuronal networks may underlie the operant conditioning of respiratory behaviour (Spencer et al. 1996). Future studies can now be conducted to investigate neuronal and network changes which may result from long-term training procedures and to determine whether the underlying neuronal correlates and mechanisms of learning and memory are similar for operant conditioning and for classical conditioning.
In conclusion, this study has established an effective procedure for producing operant conditioning of aerial respiratory behaviour in L. stagnalis, the memory of which can persist for at least 1 month. The extent of the persistence of the memory was dependent upon the training procedure used; spaced training was found to be more effective than massed training.
We wish to thank Drs Quentin Pittman and Richard Hawkes and Ms Kim McKenney for their comments and suggestions on an earlier draft of this manuscript, Drs Wic Wildering and Petra Hermann for their assistance in the rearing of the snails used in these studies, Ms Susan Poon and Ms Brittany Salkeld for their help performing some of the preliminary experiments and Dr Wic Wildering for his advice regarding the statistical analyses of the data. This study was supported by the MRC (Canada). E.R. and J.W. were initially supported by an AHFMR Summer Research Studentship. R.C. was supported in part by the MRC and Employment Canada Pilot Project Scheme. G.S. is a Killam and AHRMR postdoctoral fellow. N.S. is an AHFMR Scientist.
References
ALEXANDER, J. E., AUDESIRK, T. E. AND AUDESIRK, G. J. (1982). Rapid, nonaversive conditioning in a freshwater gastropod. Behav.
neural Biol. 36, 391–402.
ALKON, D. L. (1984). Calcium-mediated reduction of ionic currents:
A biophysical memory trace. Science 226, 1037–1045.
CAREW, T., PINSKER, H. AND KANDEL, E. (1972). Long-term
habituation of a defensive withdrawal reflex in Aplysia. Science 175, 451–454.
CAREW, T. ANDSAHLEY, C. (1986). Invertebrate learning and memory: from behavior to molecules. A. Rev. Neurosci. 9, 435–487. CASTELLUCCI, V., BLUMFELD, H., GOELET, P. ANDKANDEL, E. (1989).
Inhibitor of protein synthesis blocks long-term behavioral
sensitization in the isolated gill withdrawal reflex of Aplysia. J.
Neurobiol. 20, 1–12.
COOK, D. AND CAREW, T. (1989). Operant conditioning of head-waving in Aplysia. III. Cellular analysis of possible reinforcement pathways. J. Neurosci. 9, 3115–3122.
GELPERIN, A. (1975). Rapid food aversion learning by a terrestrial
mollusc. Science 189, 567–570.
INOUE, T., TASASAKI, M., LUKOWIAK, K. AND SYED, N. (1996a).
Identification of a putative mechanosensory neuron in Lymnaea: characterization of its synaptic and functional connections with the whole-body withdrawal interneuron. J. Neurophysiol. 76, 3230–3238.
INOUE, T., TASASAKI, M., LUKOWIAK, K. AND SYED, N. (1996b). Hypoxia induced respiratory patterned activity in Lymnaea originates at the periphery: role of the dopamine cell. Soc. Neurosci. 26, 452.2.
JONES, J. (1961). Aspects of respiration in Planorbis corneus L. and
Lymnaea stagnalis L. (Gastropoda: Pulmonata). Comp. Biochem. Physiol. 4, 1–29.
KANDEL, E. (1989). Genes, nerve cells and the remembrance of things
past. J. Neuropsychiatry 1, 103–125.
KEMENES, G. AND BENJAMIN, P. (1994). Training in a novel
environment improves the appetitive learning performance of the snail, Lymnaea stagnalis. Behav. neural Biol. 61, 139–149. KIMBLE, G. A. (1961). ‘Hilgard and Marquis’ Conditioning and
Learning, second edition. New York: Appleton-Century-Croft.
KOJIMA, S., NANAKAMURA, H., NAGAYAMA, S., FUJITO, Y. ANDITO, E. (1997). Enhancement of an inhibitory input to the feeding central pattern generator in Lymnaea stagnalis during conditioned taste-aversion learning. Neurosci. Lett. 230, 179–182.
LUKOWIAK, K., RINGSEIS, E., SPENCER, G., WILDERING, W. ANDSYED, N. (1996a). Operant conditioning of aerial respiration in Lymnaea.
J. exp. Biol. 199, 683–691.
LUKOWIAK, K., SPENCER, G., INOUE, T., COTTER, R. ANDSYED, N.
(1996b). Long-term memory of operant conditioning behaviour in
Lymnaea. Soc. Neurosci. Abstr. 26, 573.1.
MACKINTOSH, N. J. (1974). The Psychology of Animal Learning. New York: Academic Press.
MPITSOS, G. J. AND LUKOWIAK, K. (1985). Associative and non-associative learning in gastropod molluscs. In The Mollusca, vol. 8, Neurobiology and Behavior (ed. A. O. D. Willows), pp. 95–267. New York: Academic Press.
SPENCER, G., INOUE, T., COTTER, R., SYED, N. AND LUKOWIAK, K. (1996). Alterations in synaptic activity to and within a CPG may underlie the operant conditioning of aerial respiratory behaviour in
Lymnaea. Soc. Neurosci. Abstr. 26, 553.7.
SYED, N., BULLOCH, A. AND LUKOWIAK, K. (1990). In vitro reconstruction of the respiratory central pattern generator of the mollusk Lymnaea. Science 250, 282–285.
SYED, N. I., RIDGWAY, R. L., LUKOWIAK, K. ANDBULLOCH, A. G. M.