r-Indian J. Plant Physiol., Vol. XXV, No.3, pp. 271-275 (September 1982) I
EFFECT OF ANTIBIOTICS ON THE GERMINATION AND
PROTEASE ACTIVITY OF MAIZE SEEDS
BANDANA BOSE, H. S. SRIVASTAVA'" AND S. N. MATHUR
Plant Physiology Laboratory, Department of Botany
University of Gorakhpur, Gorakhpur-273 001
(Revised: April 24, 1982)
•
SUMMARY
The antibiotics chloramphenicol and cycloheximide inhibited the germination of maize seeds and protease activity in the endosperm, the latter be jng more effective tha n the former.
INTRODUCTION
There is evidence that the loss of proteins during germination is accompanied by activation/synthesis of the enzyme protease which mediate the degradation of protein (Pernollet. 1978). It has also been reported earlier that the chloramphenicol and cycloheximide both are inhibitory for the germination of many seeds (Srivastava
et a/., 1972, Srivastava, 1976). In order to gain some insight into the regulatory mechanism of such actiVity, the effect of protein inhibitors has been studied during the germination of
lea mays.
MATERIALS AND METHODS
Seeds of
lea mays
L. cv. Sankar makka Ganga-2 were used in the present investigation. After sterilisation with 1%
solution of bleaching powder for 5 min, the seeds were washed thoroughly with distilled water. Seeds were germinated on moist filter papers (moistened with antibiotics, i. e., chloramphenicol (3.1 mM) or cycloheximide (0.35 mM), prepared in 10 mM Ca (NOah solution in large petri dishes in presence of diffuse light and at 30±2C.For protease activity the endosperms were dissected out from seeds, soaked in sterile water for 2 h and then they were incubated In the desired concentration of antibiotics (Chloramphenicol (3.1 mM) or cycloheximide (0.35 mM). The solution was kept in the sterile incubation flasks each containing ten excised endosperms. To avoid the bacterial contamination, Penicillin (10 !Jog/mn was included in the incubation medium.
Assay for protease. One gm freeze dried endosperms were extracted thoroughly with 2 mt of 0.2M acetate buffer (pH 3.8) containing 5.0 mM (;-mercap toethanol by porcelin mortar and pestle at 2 to 4"C. The extract was centrifuged at
---.----.---~.
-272 BOSE, SRIVASTAVA AND MATHUR
19,000 g for 10 min. The supernatant was assayed for proteolytic activity with a 5% solution of denatured haemoglobin as substrate. Each assay routinely contained 12.5 mg of substrate, 1.2 mM EDTA prepared in 0.05 M acetate buffer of pH 3.8 and 0.025 ml of enzyme adjusted to a total volume of 0.525 ml. The mixture was incubated in a water bath at 40°C with constant shaking. After 30 min the reaction was stopped by adding an equal volume of 10% TCA solution. The mixture was kept for 15 min at 2 to 4°C for complete precipitation. After that it was centrifuged at 19,000 9 for 10 min and supernatant was collected. The release of amino ac~ds
due to hydrolysis of haemoglobin was measured by Folin-Ciocalteau reagent (Lowry
et aI., 1951) and calculated on the basis of a standard curve for Bovine Serum
Albumin (BSA).
RESULTS AND DISCUSSION
Effects of chloramphenicol and cycloheximide on germination of maize seeds were studied (Table I). Soaking seeds either in chloramphenicol or in cycloheximide for 40
to
50 h significantly inhibited the germination of seeds. In all treatments, inhibition of Igermination was more pronounced with cycloheximide than with chloramphenicol. A pre-soaking treatment with Ca (N03h
had liffle effect on the inhibition of germination, although the inhibition decreased from 57 to 26% with . chloramphenicol and 97 to 68% with cycloheximide when treatment period wasreduced from 50 to 40 h as compared
to
control.TABLE I. Effect of antibiotics on the germination of maize seeds
Preliminary soaking time Soaking time with %germination
with Ca (NO,). solution antibiotics solution Chloramphenicol Cyclohexim ide
(h)_..~~_.~_ _.~~...:.(h~)_ _ _~.~_~_._._ _ _ _ _ _ _ _ _ _ _ _ _ _ _.~~
o
50 35 22 48 40 8
4 46 41 12
6 44 50 19
10 40 61 26
- -..
-~-.---Control values fo( germination with chloramphenicol and cycloheximide were 82 and 83, respectively. Seeds were germinated for 50 h.
Inhibition of germination of maize seeds by chloramphenicol has been repor
."
ANTIBIOTICS ON GERMINATION AND PltOTBASB ACTIVITY 273 chloramphenicol inhibits seed germination by inhibiting the process of respiration (Srivastava, 1976). Further growth and development of early seedling are regulated by nucleic acids and proteins (Hadi and Krishna Murty, 1967), which are synthesized
de novo.
The addition of cycloheximide may inhibit seed germination by inhibiting the synthesis of structural and functional proteins.\
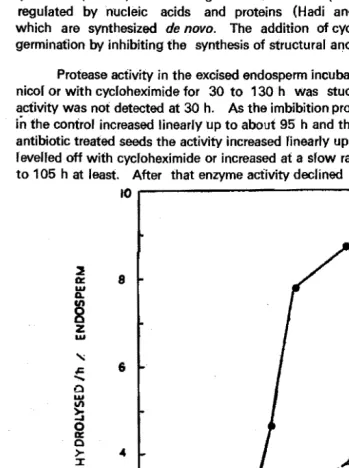
Protease activity in the excised endosperm incubated with either chloramphe nicol or with cycloheximide for 30 to 130h was studied (Fig. 1). The enzyme activity was not detected at 30 h. As the imbibition progressed, the enzyme activity
in
the control increased linearly up to about 95 h and then it started declining. In antibiotic treated seeds the activity increased finearly up to 48 h and then either it reverted off with cycloheximide or increased at a slow rate with chloramphenicol up to 105 h at least. After that enzyme activity declined in these samples also ..to
.2
Q: 8
w
§
Go
Z
w
6
1=
"
...
£) w
~
'"
0
,A
Q:
0 I · \
)0
r
,I.
\~.
,
T',
w
t-z
.,.
..
/
"
\
O -,...0---0--- __
....0---'\
'
Q:
,
\,
Q. \
,
,
\
Ot 2 ~ \
E
,
'
\
'
~
\ A
\
\
\
'.
o
100 120 140
o
20 40INCUBATION TIME (hi
Fig. 1. Effect of different antibiotics on protease activity of excised maize endosperms incubated for various periods.
0 - 0 : Control, 0 - 0 : Cycloheximide, l::;.-l::,. : Chloramphenicol
~74 BOSE, SRIVASTAVA AND MATHUll
Inhibition of enzyme activity by CYcloheximide has been reported in other systems 'also (Srivastava
et al.,
1972; Harvey and Oaks, 1974; Peterson and Huffaker, .197~). The insensitivitY of protease to chloramphenicol and cytloheximide during first 48 h of incubation may suggest that during this phase the linear .increase in enzyme activity is due to its activation rather than to its synthesis. In Sorghumsp. the enzyme is present in the seed but it is inactive before germination starts (Adams and Noveflie. 1975). On the other hand in mung bean. induction of pro tease activity during germination involved
de novo
synthesiS (Baumgartner' and Chrispeels, 1977). It appears that the proteases are synthesized on '80 S cytoplasmic ribosomes at least during the senescence of leaves (Peterson and Huffaker. 1975). Therefore. the inhibition of protease with chloramphenicol. a speCific inhibitor of protein synthesis on 70 S ribosomes (Anderson and Smillie, 1966), is lower than that with the cycloheximide. Nevertheless. at least a partial inhibition by chloram phenicol suggests that a part of the enzyme is synthesized in the cell organelles containing 70 S ribosomes. It is also possible that chloramphenicol inhibits protease activity by inhibiting respiration [(Srivastava, 1976). Zein, the major protein of maize endosperm, is .located in protein bodies. In many systems, autolysis of storage protein'in the protein bodies begins while the external membrane remains intact (Ashton, (1976). This may suggest that protease is synthesized in protein bodies. This may be also due to insensitivity of protease to chloramphenicol and cycloheximide during first 48 h of incuba tion. Once the membrane of protein bodies is hydrolysed by peripheral enzymes, the release of protein and amino acids from these bodies may induce synthesis of protease in the cytoplasm. Apparentfythis synthesis (after 48 h) is sensitive to cycloheximide and chloramphenicol.ACKNOWLEDGEMENT
One of us (B. B.) is grateful to C. S. I. R., New Delhi for financial assistance. REFERENCES
Adams. C. A. and Novellie, L (1975). Acid hydrolases and autolytic properties of protein bodies and spherosomes isolated from ungerminated seeds of Sorghum hieolor (linn) Moench. PI.
Physiol.• 55: 7-11.
Anderson. l. A. and Smillie. R. A. (1966). Binding of chloramphenicol by ribosome from chloroplasts.
Biochem. Biophy. Res. Comm., 23 : 535-39.
Ashton. F. M. (1976). Mobilization of storage proteins of seeds. Ann. Rev. PI. Physiol., 27: 95-117.
Baumgartner, B. and Chrispeels. M. J. (1977). Purification and characterization ofVicillin peptidehydro lase. the major endopeptidase in the cotyledons of mung bean seedlings. Eur. J. Biochem••
77: 223-34.
Ching. T. M. (1972). Metabolism of germinating seeds. In. Seed Biology. (ed. T. T. KOZlowski). Acad. Press. N. Y. and Lond.
Hadi, S. M. and Krishna Murty. C. R. (1967). A cell free protein synthesizing system from Cicer arietinum seedlings. Ind. J. Biochem.• 4: 1-6.
,
ANTIBIOTICS ON GERMINATION AND PROTEASE ACTIViTY
275
Harvey, B.M. R. and Oaks, A. (1974). The hydrolysis of endosperm pr~tein in Zea mays, PI. Physiol., 53: 453-57.
Koller, D., Mayer, A. M., Poljakoff-Mayber, A. and Klein, S. (962). Seed germination. Ann. Rev. PI. Physiol., 13 : 436-64.
'\ Lowry, O. H., Rosenbrough, N. J., Farr, A. J. and Randall, R. J. (1951). Protein measurement with the Folin phenol reagent. J. BioI. Chem., 193 : 265-75.
Pernollet, J. C. (1978). Protein bodies of seeds: Ultrastructure, biochemistry, biosynthesis and degrada tion. Phytochem., 17: 1473-80.
Peterson, L. W. and Huffaker, R. C. (1975). Loss'of ribulose-1, 5-diphosphate carboxylase and increase in proteolytic activity during senescence of detached primary barley leaves. PI. Physiol., 55 : 1009-15.
Srivastava, A. K., Azhar, S. and Krishna Murty, C. R. (1972). Inhibition of germination in Cicer al/etinum. Phytochem., 11 : 3181-85.
Srivastava, H. S. (1976). Inhibition of maize seedling growth by chloramphenicol. Indian J. Plant Physiol., 19: 53-59.