HELGOL,~NDER MEERESUNTERSUCHUNGEN Helgolhnder Meeresunters. 49, 495-505 (1995)
P r e d i c t i v e e c o s y s t e m r e s e a r c h in t h e W a d d e n S e a
K. Reise
Biologische Anstalt Helgoland, Wattenmeerstation Sylt; D-25989 I_dst, Germany
ABSTRACT: Predictive ecosystem research n e e d s a pluralistic approach. Retrospective studies reveal the initial causes of ongoing ecological change. In the W a d d e n Sea, i n h e r e n t ecosystem stability may be falsely assumed, b e c a u s e the effects of m o d e r n coastal architecture a n d of a n t h r o p o g e n i c eutrophication to some extent c o m p l e m e n t each other. Expected e n v i r o n m e n t a l c h a n g e s often h a v e corresponding p h a s e s in the past which may serve as a model to predict ecological implications. Historically, quantitative ecology e n t e r e d the W a d d e n Sea, via fisheries research, from the oceanic side. Quantified material fluxes may reveal i m b a l a n c e s which are indicative of the rough direction of ecosystem change. For ecosystem research to contribute to the m a i n t e n a n c e of the W a d d e n Sea as a centre of coastal organisms, quantitative k n o w l e d g e of resources a n d ecosystem metabolism must be s u p p l e m e n t e d by qualitative k n o w l e d g e of habitat r e q u i r e m e n t s and species i n t e r d e p e n d e n c e s . Qualitative ecology e n t e r e d the W a d d e n Sea from the l a n d w a r d side. Extending this approach to anticipatory field experiments may help to predict ecological c h a n g e s at the species level.
I N T R O D U C T I O N
P h y s i c a l l y t h e W a d d e n S e a is a n e d g e ; a g i g a n t i c b o d y of w a t e r p a s s i n g to a n d fro w i t h t h e t i d e s . W i t h its 8 0 0 0 k m 2, t h e W a d d e n S e a c o m p r i s e s m e r e l y 1.4 % of t h e s i z e of t h e N o r t h S e a , a n d a b o u t 1.6 % of its r i v e r i n e w a t e r s h e d ( i n c l u d i n g t h e r i v e r R h i n e w i t h its t r i b u t a r i e s ) . A c c o r d i n g l y , t h e p h y s i c a l p r o p e r t i e s of t h e W a d d e n S e a w a t e r s a r e s e t b y p r o c e s s e s a n d e v e n t s i n t h e s e a d j o i n i n g e c o s y s t e m s .
B i o l o g i c a l l y , h o w e v e r , t h e W a d d e n S e a is a c e n t r e . O r g a n i s m s t e n d to a g g r e g a t e a t h a b i t a t e d g e s . S e a l s c h o o s e s a n d b a n k s a t t i d a l i n l e t s as p e a k h a u l o u t s , a n d f o r a g e i n t h e c h a n n e l s . For E a s t A t l a n t i c w a d i n g b i r d s , b r e e d i n g i n t h e n o r t h a n d w i n t e r i n g i n t h e s o u t h , t h e W a d d e n S e a is a n e s s e n t i a l s t a g m g a r e a to r e p l e n i s h t h e i r f a t r e s e r v e s . Fish, s h r i m p a n d c r a b s u s e t h e t i d a l flats a s n u r s e r i e s . T h e p o p u l a t i o n s of l u g w o r m s (
Arenicola
marina
L.) a n d t e l l i n i d c l a m s(Macoma badthica
[L.]) c o n c e n t r a t e a t t h e l a n d w a r d e d g e s of t h e t i d a l flats, u s i n g t h e s e as n u r s e r i e s . B e n t h i c d i a t o m s a r e m o s t a p p a r e n t o n i n t e r t i d a l m u d i n s h e l t e r e d b a y s . T h e r e a r e m a n y g r o u p s of o r g a n i s m s w h i c h a t t a i n t h e i r p e a k d e n s i t i e s in t h e W a d d e n S e a ; t h u s , t h e e c o l o g i c a l i m p o r t a n c e of t h i s c o a s t a l a r e a r e s i d e s in its b i o l o g i c a l c o m p o n e n t s r a t h e r t h a n its p h y s i c a l a t t r i b u t e s . A n y e c o s y s t e m r e s e a r c h i n t h e W a d d e n S e a h a s to b e a w a r e of t h e e c o t o n e p r o p e r t i e s of t h i s t r a n s i t i o n z o n e b e t w e e n t h e s e a a n d t h e l a n d ~ t h its p h y s i c a l t r a n s p o r t a t i o n of w a t e r m a s s e s , a n d o r g a n i s m s m i g r a t i n g , or d r i f t i n g i n a n d out. E c o s y s t e m r e s e a r c h c a n n o t s t o p a t t h e g e o g r a p h i c a l c o n f i n e s of t h e W a d d e n S e a .Tide . . . . . . .
0 N o r t h e r n / \ "" ~. 1.5 - 3 m {~ W a d d e n
/ \ ~- ~, ~ Sea
i I
\ \
~" ~ ~ , ~
\ ~3 m I
I \
/
I 1.5-3m \ 9
,
S o u t h e r n W a d d e n S e a
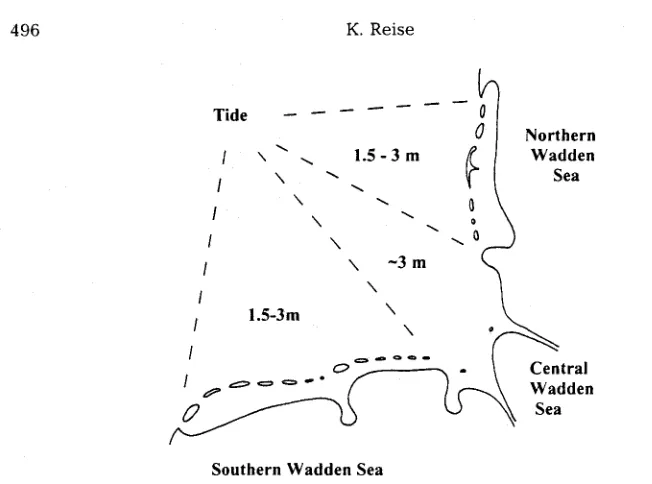
Fig. 1. The Wadden Sea consists of three major regions. Approximate tidal ranges are indicated (see text)
distinction l e a d s to t h r e e major r e g i o n s (Fig. 1). T h e n o r t h e r n and s o u t h e r n r e g i o n s h a v e tidal r a n g e s b e t w e e n 1.5 a n d 3 m. A corollary of this r a n g e is a chain of b a r r i e r islands a n d sand ridges parallel to the coast (Hayes, 1975). T h e C e n t r a l W a d d e n S e a has tidal r a n g e s close to 3 m. T h e resulting strong on- a n d offshore currents p r e v e n t barrier island d e v e l o p m e n t . T h e northern r e g i o n f a c e s the p r e v a i l i n g w e s t e r l y winds, w h i l e the south- ern r e g i o n faces northward. In the north river entries are small, and v a r i a t i o n in salinity is low. In the C e n t r a l W a d d e n Sea. river e n t r i e s are large, a n d variation in salinity is high. In t h e south, salinity variations are i n t e r m e d i a t e . For further r e g i o n a l subdivisions, s e e D i j k e m a et al, (1989). Material fluxes a n d a b u n d a n c e s of organisms are e x p e c t e d to differ b e t w e e n t h e s e regions. No s u b r e g i o n within the W a d d e n Sea is r e p r e s e n t a t i v e of the whole.
In this article, the e c o l o g i c a l history of the W a d d e n Sea as well as t h e history of e c o l o g i c a l r e s e a r c h in this a r e a is s c r e e n e d for implications r e l e v a n t to p r e d i c t i v e e c o s y s t e m research. Predictions should rest on e x p e r i e n c e of past p r o c e s s e s a n d events. Extrapolations from i m p o r t - e x p o r t q u a n t i t i e s and t u r n o v e r rates d e s c r i b e w h a t m a y b e possible. W h a t is likely to be r e a l i z e d should b e e x p l o r e d e x p e r i m e n t a l l y .
E C O L O G I C A L HISTORY
Predictive research in the Wadden Sea
497
Apparently, there were several reversals between phases of inundation and erosion, and
phases of marsh accretion and sedimentation, and these did not occur in concert
throughout the Wadden Sea. The local responses to supraregional trends in sea-level rise
differed according to specific hydrological and sedimentological conditions. This
phenomenon of inherent, ongoing and asynchronous change in geomorphology and
hydrodynamics is of importance, when ecological measurements taken at a particular site
and time are to be extrapolated.
For 1000 years people progressively interfered with these natural dynamics, first in
the south and then in the north of the Wadden Sea (Bantelmann, 1966; Wolff, 1992a).
Marshlands were diked and drained. Tidal flats were also diked to create freshwater
storage basins, or simply to straighten out the coastline. Natural mud accretion was
promoted. Dams were built across tidal flats and channels. The building material for
dikes and dams was often extracted from the seaward flats. Erosional shores have been
fixed by stone walls. Sluices and storm-surge barriers control the water flow b e t w e e n the
sea, land and estuaries. Deep harbours with strong jetties were built, and some channels
are permanently d e e p e n e d by dredging, to let bigger ships travel further inland.
Together these activities resulted in a modern coastal architecture, where the
smooth, irregular and dynamic transition of the land blending into the sea is displaced by
a seaward, abrupt and straight dikeline (Fig. 2). The relative proportions of habitat sizes
have changed. The slope from high tide line towards the subtidal gullies steepened,
because the dikeline moved seaward and the gullies became wider (Misdorp et al., 1989).
-
kk~______
~ "''":::~:':"::::::'::::':
~ { i i i ! i i i i ~
~
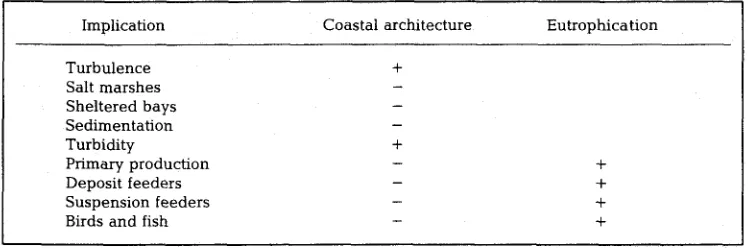
Table 1. Ecological implications of modern coastal architecture and anthropogenic eutrophication in the Wadden Sea, increase (+) and decrease (-)
Implication Coastal architecture Eutrophication
Turbulence +
Salt marshes Sheltered bays Sedimentation
Turbidity +
Primary production Deposit feeders Suspension feeders Birds and fish
+ + + +
T h e s e f u n d a m e n t a l c h a n g e s in size a n d shape of the W a d d e n Sea e c o t o n e have several ecological implications (Table 1):
(1) In spite of sea-level rise, there are no more l a n d w a r d e x t e n s i o n s of the W a d d e n Sea. Dikes or stone wails p r e v e n t the sea g r a d u a l l y d y i n g out a n d d i s s i p a t i n g its e n e r g y while flooding the land. C o n s e q u e n t l y , t u r b u l e n c e has increased.
(2) E m b a n k m e n t s a n d r e c l a m a t i o n s of the w e t l a n d s e l i m i n a t e d salt a n d brackish reed marshes, t h e r e b y d e c r e a s i n g the detrital i n p u t s into the r e m a i n d e r of the W a d d e n Sea. A c c o r d i n g to D i j k e m a (1987), the rate of e m b a n k m e n t was faster t h a n the accretion of n e w marshes. In 1600, a b o u t 15 % of the intertidal zone was covered b y salt marshes. Today this is 3 %.
(3) Almost all sheltered bays v a n i s h e d , b e c a u s e at these sites e m b a n k m e n t s were technically most feasible a n d cost effective. Such bays were the n a t u r a l sites of s e d i m e n - tation a n d m u d accretion. Where b a y s still exist, s e d i m e n t a t i o n seems to k e e p pace with sea-level rise (Frels & Demuth, 1987). However, the overall a m o u n t of s e d i m e n t a t i o n a n d m u d accretion d e c l i n e d b e c a u s e the sheltered areas most suitable for t h e s e processes b e c a m e scarce.
(4) As a c o n s e q u e n c e of h i g h e r t u r b u l e n c e , and d i m i n i s h e d mud a c c r e t i o n in salt m a r s h e s a n d sheltered bays, turbidity in the tidal waters increased. In the S o u t h e r n W a d d e n Sea, secchi disc visibihty has d e c r e a s e d b y 1.4 m since the 1930s (Jonge et al., 1993). D r e d g i n g of h a r b o u r s a n d n a v i g a t i o n c h a n n e l s a n d the e x t r a c t i o n of s a n d a n d shell gravel, also c o n t r i b u t e to h i g h e r turbidity.
(5) Increased turbidity limits light p e n e t r a t i o n , a n d h a m p e r s p h o t o s y n t h e s i s in the tidal waters. T o g e t h e r with the loss of salt a n d reed m a r s h e s a n d the loss of s h e l t e r e d bays w h e r e epipelic diatoms a b o u n d , the overall effect is a d e c r e a s e m p r i m a r y p r o d u c t i o n w i t h i n the W a d d e n Sea.
Predictive r e s e a r c h in the W a d d e n Sea 499 (7) It is e x p e c t e d that birds a n d fish find less food w h e n the b e n t h i c f a u n a is affected. The lost shallow b a y s w e r e p r e f e r r e d f e e d i n g g r o u n d s for w a d e r s a n d fish nurseries. In the case of the e m b a n k e d Z u i d e r z e e , a r a c e of h e r r i n g lost its s p a w n i n g g r o u n d s a n d d i s a p p e a r e d c o m p l e t e l y (Wolff, 1992a).
T e c h n o g e n i c s u b s t a n c e s , fishing, h u n t i n g , a n d tourism a g g r a v a t e d s o m e of t h e s e e c o l o g i c a l implications of the m o d e r n c o a s t a l a r c h i t e c t u r e (Wolff & Zijlstra, 1980; Wolff, 1992b). On the other h a n d , a n t h r o p o g e n i c e u t r o p h i c a t i o n c a u s e d a p a r t i a l reversal. In r e c e n t d e c a d e s , n o n - d i a t o m p l a n k t o n i n c r e a s e d (Cadee, 1992), g r e e n a l g a l m a t s s p r e a d on tidal flats (Reise et al., 1989), a n d t h e b o t t o m f a u n a i n c r e a s e d in a b u n d a n c e a n d b i o m a s s (Beukema, 1992). P r e s u m a b l y , t h e s e e u t r o p h i c a t i o n effects also i n c r e a s e d bird n u m b e r s , but this cannot, with certainty, b e d i f f e r e n t i a t e d from bird p r o t e c t i o n m e a s u r e s (Vauk et al., 1989; Wolff, 1992a). This e n r i c h m e n t p h a s e of e u t r o p h i c a t i o n e v e n t u a l l y p r o c e e d s t o w a r d s d e p a u p e r a t e d b i o t a w h e n o x y g e n conditions b e c o m e d e f i c i e n t (Rosen- berg, 1985). At the time, h o w e v e r , e u t r o p h i c a t i o n effects on the e c o s y s t e m are o p p o s i t e to those of the n e w coastal architecture. T h e results of such a n t a g o n i s t i c c h a n g e s s h o u l d not b e confused with i n h e r e n t e c o s y s t e m stability, w h i c h r e q u i r e s n e g a t i v e f e e d b a c k s b e t w e e n processes.
This m a y e x e m p l i f y the i m p o r t a n c e of r e c o n s t r u c t i n g p a s t d e v e l o p m e n t s to u n d e r - s t a n d the p r e s e n t state. A r e t r o s p e c t i v e a p p r o a c h is also l i k e l y to i m p r o v e our ability to p r e d i c t future directions of e c o l o g i c a l d e v e l o p m e n t . M o s t of the p a s t p r o c e s s e s will also p l a y k e y roles in the future. Of p a r t i c u l a r r e l e v a n c e is a c o m b i n a t i o n of o n g o i n g e u t r o p h i c a t i o n with climatic c h a n g e , a n d t h e c o n c o m i t a n t a c c e l e r a t i o n in s e a - l e v e l rise. The l a t t e r will p r o v o k e new, a m b i t i o u s p l a n s b y coastal e n g i n e e r s to k e e p the s e a a w a y from the land. Ecologists will b e a s k e d to e s t i m a t e the e c o l o g i c a l i m p l i c a t i o n s of a l t e r n a t e options. A t h o r o u g h e v a l u a t i o n is n e e d e d of the e c o l o g i c a l effects of the s e a w a r d p r o g r e s s i o n of the d i k e l i n e d u r i n g the last centuries, p a r t i c u l a r l y in this case. This r e q u i r e s the joint efforts of p a l e o e c o l o g i c a l research, historical inquiries, r e t r o s p e c t i v e m o d e l l i n g a n d field e x p e r i m e n t s s i m u l a t i n g p a s t events. In m a n y respects, the future is m e r e l y an e x t e n s i o n of the past. Therefore, a s o u n d u n d e r s t a n d i n g of the p a s t m a y b e the b e s t p r e c a u t i o n to give e c o l o g i c a l a d v i c e in a c h a n g i n g W a d d e n Sea.
HISTORY O F E C O L O G I C A L RESEARCH
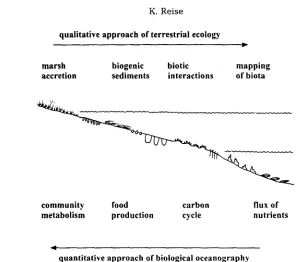
Two tribes of ecologists i n v a d e d the W a d d e n Sea. O n e e n t e r e d the tidal zone from the terrestrial side. a n d a n a l y s e d the e c o l o g i c a l w e b p r i m a r i l y with a q u a l i t a t i v e a p p r o a c h . The o t h e r tribe is of o c e a n i c origin, was n u r s e d by fishery research, a n d followed p r i m a r i l y a q u a n t i t a t i v e a p p r o a c h (Fig. 3).
The terrestrial tribe s t a r t e d with Karl M 6 b i u s (1877, 1893). The n a t u r a l o y s t e r b e d s in the N o r t h e r n W a d d e n s e a suffered from o v e r - e x p l o i t a t i o n . M 6 b i u s was c a l l e d for, came, a n d d r e d g e d the first e c o l o g i c a l c o m m u n i t y c o n c e p t from the b o t t o m of the W a d d e n S e a (Reise. 1980, 1990). His c o n c e p t is still alive, while the o y s t e r b e d s are l o n g gone. MSbius v i e w e d a n o y s t e r b e d as an i n t e r a c t i n g a s s e m b l a g e of populations. U n d e r the local p h y s i c a l conditions, planktivors, p r e d a t o r s a n d p a t h o g e n s m a i n t a i n a c o m m u n i t y in a state of equilibrium, u n l e s s o v e r - e x p l o i t a t i o n c a u s e s a c o m p l e t e r e o r g a n i z a t i o n .
qualitative approach of terrestrial ecology
marsh biogenic biotic mapping
accretion sediments interactions of biota
community food carbon flux of
metabolism production cycle nutrients
quantitative approach of biological oceanography
Fig. 3. Topics of ecological research chosen by scientists entering the Wadden Sea from the landward side labove} and from the oceanic side
W o h l e n b e r g {1931, 1937. 1953) a n d Linke (1939) a n a l y s e d salt m a r s h accretion, b i o g e n i c m u d [ormation a n d other o r g a n i s m - s e d i m e n t interactions. They also m a p p e d a n d des- cribed the p a t t e r n of habitats a n d species associations, as it was practised in terrestrial p l a n t sociology.
Later cascades along this stream of research included, a m o n g others, z o n a t i o n of arthropod n i c h e s in salt marshes ( H e y d e m a n n , 1967. 1979), field e x p e r i m e n t on biotic interactions a n d o r g a n i s m - e n v i r o n m e n t i n t e r d e p e n d e n c e on tidal flats (Reise, 1985), a n d r e p e a t e d m a p p i n g of habitats a n d o r g a n i s m s i n the intertidal zone to trace ecological history Icf. Michaelis, 1987).
This qualitative research on ecological interactions r e m a i n e d s e p a r a t e d from q u a n - titative r e s e a r c h on the m e t a b o l i s m of the W a d d e n Sea ecosystem. I n s p i r e d b y the planktologist Victor H e n s e n , Dahl (1893) was the first to c o u n t the tidal flat f a u n a per square metre He f o u n d few species, b u t m a n y individuals, a n d a t t r i b u t e d this to physical h a r s h n e s s a n d resource richness, respectively. T h e latter i m p l i e d that t h e r e w a s p l e n t y to [eed u p o n i n t h e W a d d e n Sea
Fishery biologists identified the W a d d e n Sea as a major n u r s e r y area for plaice a n d herring (Garstang, 1909; B i i c k m a n n , 1934]. T h a m d r u p (1935) q u a n t i f i e d c o m m u n i t y m e t a b o l i s m o n tidal flats. He f o u n d o x y g e n c o n s u m p t i o n to b e h i g h on tidal flats, c o m p a r e d to that of a d e c i d u o u s forest floor. In the same r e g i o n of the N o r t h e r n W a d d e n Sea, Smidt (1951} estimated food p r o d u c t i o n in the plaice nurseries, a n d G m n t v e d (1962} m e a s u r e d p r i m a r y production.
Predictive research in the W a d d e n Sea 501 p l i n a r y individuals. Only t h e n did Dutch projects, in the Ems-Dollard estuary a n d in the w e s t e r n W a d d e n Sea (with their joint efforts) start to quantify the c a r b o n cycle a n d n u t r i e n t fluxes. This research was no l o n g e r a n outgrowth of fishery science. It was stimulated b y a c o n c e r n a b o u t the W a d d e n Sea as a n a t u r e area, t h r e a t e n e d by e m b a n k m e n t , waste discharge a n d other h u m a n impacts (Wolff & Zijlstra, 1980).
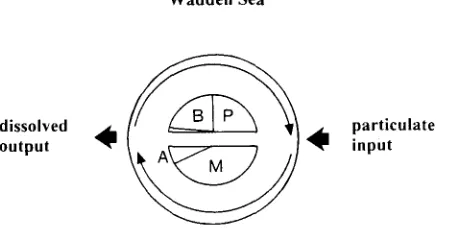
From the q u a n t i t a t i v e perspective, the W a d d e n Sea is r e g a r d e d as a n importer of organic particles a n d a n exporter of dissolved metabolic e n d p r o d u c t s (Fig. 4), a m o n g them the n u t r i e n t s phosphorus a n d n i t r o g e n (Postma, 1982, 1984). In spring a n d s u m m e r ,
Wadden Sea
dissolved
output
particulate
~ll
~ll
input
Fig. 4. Metabolism of the Wadden Sea ecosystem. Primary production is primarily accomplished by benthic (B) and pelagic microalgae (P), mineralization by "microorganisms" (MI in a wider sense,
and by larger animals (A)
e n d - p r o d u c t s of m i n e r a l i z a t i o n are t a k e n up by p l a n t s within the W a d d e n Sea. Primary p r o d u c t i o n is a c c o m p l i s h e d primarily by b e n t h i c a n d pelagic microalgae in a b o u t e q u a l proportions (Cadee & H e g e m a n n , 1977).
In addition to imported seston, this p r i m a r y p r o d u c t i o n is c o n s u m e d by m i c r o o r g a n - isms a n d the m a c r o b e n t i c fauna, r o u g h l y in a 4 to 1 ratio (Kuipers et al., 1981; Wilde & B e u k e m a , 1984; Witte & Zijlstra, 1984). A b o u t 1 0 % of the c a r b o n c o n s u m e d b y the m a c r o b e n t h i c f a u n a is e v e n t u a l l y p a s s e d on to shrimp, fish a n d birds in a p p r o x i m a t e l y e q u a l proportions. A n i n c r e a s e d i n p u t of o r g a n i c material d u r i n g the last d e c a d e s p r e s u m a b l y c a u s e d a n increase in all major divisions of the c a r b o n cycle (Jonge & Postma, 1974~ Cad6e, 1984; B e u k e m a & Cad6e, 1986).
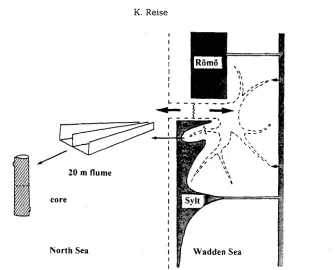
Recently, ecosystem research in the N o r t h e r n W a d d e n Sea started with the hypothesis that i n p u t s a n d outputs were i m b a l a n c e d (Reise. 1990). Net differences b e t w e e n imports a n d exports are expected to indicate direction a n d m a g n i t u d e of ecosystem c h a n g e . A major p r o b l e m is that the q u a n t i t i e s t r a n s p o r t e d b a c k a n d forth with the tides are both large a n d variable. C o n s e q u e n t l y , small n e t differences b e t w e e n inflow a n d outflow are difficult to measure. Therefore, i n p u t - o u t p u t m e a s u r e m e n t s are per- formed at four orders of m a g n i t u d e (Fig. 5).
I:
I
I
J l
I x*.~
I t~ 4 ~ "~" ~91'
rt r ~ ~
I s L.'I
I. . . . -- ~lll
/ I 11 \ ~x
1,, ~l \
20 m flume
B , '/"
%
core
~ y ~
Wadden Sea
NorthSea
Fig. 5. Exchange processes between the Wadden Sea and the North Sea are measured in the tidal inlet between two islands connected with darns to the mainland, in a tidal channel of a smafl bay within the area, in the tidal waters of a flume of 20 m in length set up on specific habitat structures,
and in sediment cores or enclosed water parcels
s u c h as m u s s e l beds, by an outdoor f l u m e of 20 m in l e n g t h (Asmus et, al., 1990). S a m p l e s are t a k e n from the e b b a n d flood currents passing t h r o u g h this flume. Finally, b a c t e r i a l p r o c e s s e s a n d gas e x c h a n g e with the a t m o s p h e r e are s t u d i e d in e n c l o s e d s e d i m e n t cores or w a t e r parcels.
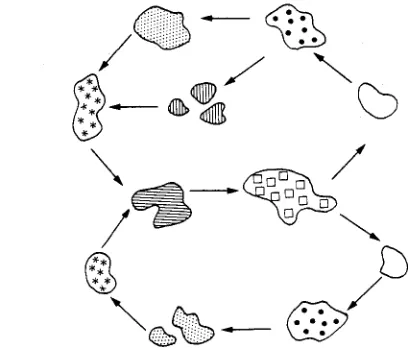
A n o t h e r r e c e n t e n d e a v o u r , this o n e m o r e in t h e tradition of q u a l i t a t i v e e c o s y s t e m research, explores the p a t c h d y n a m i c s in t h e intertidal z o n e of the W a d d e n Sea. Patches of habitat structure or biotic a s s e m b l a g e s are v i e w e d as transient, d e s y n c h r o n i z e d p h a s e s of a common, often quasi-cyclical d e v e l o p m e n t (Reise, 1991; Fig. 6). P a t c h i n e s s is a s s u m e d to slow d o w n an a d v a n c i n g process, a n d allows for partial i n d e p e n d e n c e . T h e hypothesis is that s u c h m o s a i c d y n a m i c s c o n t r i b u t e to h o m e o s t a t i c p r o p e r t i e s at the l a n d s c a p e level. T h e i d o s y n c r a c i e s of t h e p a r t i c u l a r are e x p l o r e d with r e s p e c t to t h e i r effects on t h e stability a n d r e s i l i e n c e of the whole. M o s a i c e l e m e n t s p e r c e i v e d are m u s s e l beds, g r e e n algal mats or b l a c k spots. T h e latter are sites w h e r e b l a c k a n o x i c s e d i m e n t a p p r o a c h e s the surface from below,
Predictive research in the W a d d e n Sea 503
/
Fig. 6. Mosaic dynamics on tidal flats which may contribute to overall homeostasis by asynchrony, partial independence and singularities
a s s e m b l a g e s . The results m a y t h e n be s c r e e n e d for implications on material flux rates, c a r b o n sources a n d other q u a n t i t a t i v e aspects of ecosystem relevance.
C O N C L U S I O N S
Ecosystem research provides the most c o m p r e h e n s i v e k n o w l e d g e a v a i l a b l e on specific sections of the biosphere. This k n o w l e d g e is a s k e d for w h e n familiar features in our e n v i r o n m e n t change, w h e n external processes are expected to alter the b o u n d a r y conditions for ecosystem functions, or w h e n the ecological implications "of optional policies are to be predicted.
Ecological predictions c a n n o t b e accurate b e c a u s e of the probabilistic, c o n t i n g e n t n a t u r e of the ecological processes. However. the reliability that our predictions point in the right direction can b e i m p r o v e d b y c o m b i n i n g different methodologies. (1) Historical ecology: e v e n t s a n d d e v e l o p m e n t s of the past are u s e d as models [or predictions c o n c e r n i n g e x p e c t e d situations, which r e s e m b l e some phases in the past. (2) Q u a n t i t a t i v e ecosystem research: the sign of i m b a l a n c e s in the import-export relations of the ecotone indicates the r o u g h direction of further ecological d e v e l o p m e n t s . (3) Qualitative ecosys- t e m research: field a n d mesocosm e x p e r i m e n t s a n t i c i p a t i n g expected or optional e n v i r o n m e n t a l conditions i n d i c a t e c h a n g e s i n species responses a n d species composition
This m e t h o d o l o g i c a l pluralism of c o m b i n i n g historical e x p e r i e n c e with m o d e l calcu- lations b a s e d on q u a n t i f i e d i m b a l a n c e s i n the c u r r e n t processes, and with q u a h t a t i v e insights o b t a i n e d from anticipatory experiments, afford the b e s t possible r o b u s t n e s s for ecological predictions, g i v e n the i n h e r e n t u n c e r t a i n t i e s .
c o n d i t i o n s . T h e q u a n t i t a t i v e a p p r o a c h is d e f i c i e n t b e c a u s e i t s m o d e l s i n c o r p o r a t e a
l i m i t e d s e t o f c o m p a r t m e n t s , s t a t e v a r i a b l e s , p r o c e s s e s , a n d u n i t s i n w h i c h t h e r e s u l t s a r e
e x p r e s s e d . W h e r e t h e u n i t i s m g c a r b o n , t h e r e w i l l b e n o p r e d i c t i o n o n s p e c i e s c o m p o s i -
t i o n . E x p e r i m e n t s a r e d e f i c i e n t , b e c a u s e t h e s e a r e to b e c o n d u c t e d o n s p a t i a l a n d
t e m p o r a l s c a l e s s m a l l e r a n d s h o r t e r t h a n t h o s e o f t h e s y s t e m u n d e r q u e s t i o n , a n d
t h e r e f o r e c a n n o t p r e d i c t d e v e l o p m e n t s w h i c h c h a n g e t h e i r c o u r s e a s t h e d i m e n s i o n s
e x t e n d . T h e s e d e f i c i e n c i e s m a y b e p a r t l y o v e r c o m e b y a p l u r a l i s t i c a p p r o a c h to p r e d i c t i v e
e c o s y s t e m r e s e a r c h c o n c e r n i n g t h e W a d d e n S e a .
L I T E R A T U R E C I T E D
A s m u s , H., A s m u s , R. & Reise, K., 1990. E x c h a n g e p r o c e s s e s in a n i n t e r t i d a l m u s s e l b e d : a S y l t - f l u m e s t u d y in t h e W a d d e n Sea. - Ber. Biol. A n s t H e l g o l a n d 6, 1-79.
B a n t e l m a n n , A., 1966. Die L a n d s c h a f t s e n t w i c k l u n g a n d e r s c h l e s w i g - h o l s t e i n i s c h e n W e s t k f i s t e , d a r g e s t e l l t a m B e i s p i e t N o r d f r i e s l a n d s . - Kfiste 14, 5 - 9 5 .
B e u k e m a , J. J., 1992. L o n g - t e r m a n d r e c e n t c h a n g e s in t h e b e n t h i c m a c r o f a u n a l i v i n g o n tidal flats in t h e w e s t e r n p a r t of t h e W a d d e n Sea. - Publ. Ser. N e t h . Inst. S e a Res. 20, 1 3 5 - 1 4 1 .
B e u k e m a , J. J. & C a d e e , G. C,, 1986. Z o o b e n t h o s r e s p o n s e s to e u t r o p h i c a t i o n of t h e D u t c h W a d d e n Sea. - O p h e l i a 26, 5 5 - 6 4 .
B f i c k m a n n , A., 1934. l~lber die J u n g s c h o l l e n b e v 6 1 k e r u n g d e r d e u t s c h e n W a t t e n k f i s t e d e r N o r d s e e . - Bet. dt. wiss. K o m m n . M e e r e s f o r s c h . 7, 2 0 5 - 2 1 3 .
C a d 4 e , G. C., 1984. H a s i n p u t of o r g a n i c m a t t e r into t h e w e s t e r n p a r t of t h e D u t c h W a d d e n S e a i n c r e a s e d d u r i n g t h e l a s t d e c a d e s ? - Publ. Ser. N e t h . Inst. S e a Res. I0, 7 1 - 8 2 .
C a d e e , G. C., 1992. T r e n d s in M a r s d i e p p h y t o p l a n k t o n . - Publ. Ser. N e t h . Inst. S e a Res. 20, 1 4 3 - 1 4 9 . C a d 4 e G. C. & H e g e m a n J.. 1977. D i s t r i b u t i o n of p r i m a r y p r o d u c t i o n of t h e b e n t h i c m i c r o f l o r a a n d a c c u m u i a u o n of o r g a n i c m a t t e r on a tidal flat area, B a l g z a n d . D u t c h W a d d e n S e a . - N e t h . J. S e a Res. I i 2 4 - 4 1 .
Dahl. F.. 1893. U n t e r s u c h u n g e n fiber die T h i e r w e l t d e r U n t e r e l b e . - Ber. K o m m n . w i s s . Unters. dt. M e e r e s 6 f3L149-185.
D i l k e m a . K. S., 1987. C h a n g e s in s a l t - m a r s h a r e a s in t h e N e t h e r l a n d s W a d d e n S e a after 1600. In: V e g e t a t i o n b e t w e e n l a n d a n d sea. Ed. b y A. H. L. H u i s k e r s C. W. P. M. Blom & J. R o z e m a . J u n k . D o r d r e c h t . 4 2 - 4 9 .
D i j k e m a , K. S., v a n T i e n e n . G. & v a n B e e k . J. G. 1989. H a b i t a t s of t h e N e t h e r l a n d s . G e r m a n a n d D a n i s h W a d d e n S e a 1:100.000. R e s e a r c h I n s t i t u t e for N a t u r e M a n a g e m e n t . T e x e l . 30 pp. Frels. G. & D e m u t h . A.. 1987. D e r J a d e b u s e n u n d s e i n e W a t t e n : V e r l a n d u n g d e s J a d e b u s e n s o d e r
n u r A u s g l e i c h d e s M e e r e s s p i e g e l a n s t i e g s ? - Kfiste 45. 117-121.
G a r s t a n g W . I909. T h e d i s t r i b u t i o n of t h e p l a i c e in t h e N o r t h Sea. S k a g e r a k a n d K a t t e g a t , a c c o r d - i n g to size. a g e a n d f r e q u e n c y . - Rapp. P.-v. R 6 u n C o n s . p e r m int. Explor. M e t . 11. 65-133. G r o n t v e d . J. 1962. P r e l i m i n a r y r e p o r t o n t h e p r o d u c t i v i t y of m i c r o b e n t h o s a n d p h y t o p l a n k t o n in t h e
D a n i s h W a d d e n Sea. - M e d d r D a n m . F i s k . - o g H a v u n d e r s . 3. 3 4 7 - 3 7 8 .
H a y e s , M. O. 1975. M o r p h o l o g y of s a n d a c c u m u l a t i o n in e s t u a r i e s : a n i n t r o d u c t i o n to t h e s y m - p o s i u m . In: E s t u a r i n e r e s e a r c h . Ed. b y L. E. Cronin. A c a d Press. N e w Y o r k 2, 3 - 2 2 .
H e y d e m a n n , B. 1967. D a s F r e i l a n d - u n d L a b o r e x p e r i m e n t z u r O k o l o g i e d e r G r e n z e L a n d - M e e r . - V e r h . dt. z o o l Ges. 1967, 2 5 6 - 3 0 9 .
H e y d e m a n n , B.. 1979. R e s p o n s e s of a n i m a l s to s p a t i a l a n d t e m p o r a l e n v i r o n m e n t a l h e t e r o g e n e i t y w i t h i n salt m a r s h e s . In: Ecological p r o c e s s e s in c o a s t a l e n v i r o n m e n t s . Ed. b y R. L. J e f f e r i e s & A. J. Davy. Blackwell. Oxford. 145-163.
J o n g e , V. N. d e E s s i n k K. & B o d e k k e , R.. 1993. T h e D u t c h W a d d e n Sea: a c h a n g e d e c o s y s t e m . H y d r o b i o l o g i a 265, 4 5 - 7 1 .
J o n g e , V, N. d e & P o s t m a . H., 1974. P h o s p h o r u s in t h e D u t c h W a d d e n Sea. - N e t h . J. S e a Res. 8. 139-153.
P r e d i c t i v e r e s e a r c h i n t h e W a d d e n S e a 5 0 5
K u i p e r s , B. R., Wilde, P. A. W. J. d e & C r e u t z b e r g , F., 1981. E n e r g y flow in a tidal flat e c o s y s t e m . - M a r . Ecol. Prog. Set. 5, 2 1 5 - 2 2 1 .
Linke, O., 1939. Die Biota d e s J a d e b u s e n w a t t e s . - H e l g o l / i n d e r wiss. M e e r e s u n t e r s . 1, 2 0 1 - 3 4 8 . M i c h a e l i s , H., 1987. B e s t a n d s a u f n a h m e d e s e u l i t o r a l e n M a k r o b e n t h o s i m J a d e b u s e n in V e r b i n d u n g
m i t e i n e r L u f t b i l d a n a l y s e . - Jber. ForschSt. Kfiste, N o r d e r n e y 38, 1-97.
M i s d o r p , R., S t e y a e r t , F., R o n d e , J. de & Hallie, F., 1989. M o n i t o r i n g t h e w e s t e r n p a r t of t h e D u t c h W a d d e n S e a - s e a l e v e l a n d m o r p h o l o g y . - H e l g o l / i n d e r M e e r e s u n t e r s . 43, 3 3 3 - 3 4 5 .
M S b i u s , K., 1877. Die A u s t e r u n d die A u s t e r n w i r t h s c h a f t . W i e g a n d t , H e m p e l & P a r e y , Berlin, 126 pp. M S b i u s , K., 1893. U b e r die T i e r e d e r s c h l e s w i g - h o l s t e i n i s c h e n A u s t e r n b / i n k e , i h r e p h y s i k a l i s c h e n
u n d b i o l o g i s c h e n L e b e n s v e r h ~ i l t n i s s e . - Jber. p r e u s s . A k a d . Wiss. 7, 3 3 - 5 8 .
P o s t m a , H. (Ed.), 1983. H y d r o g r a p h y of t h e W a d d e n Sea. In: E c o l o g y of t h e W a d d e n Sea. Ed. b y W. J. Wolff. B a l k e m a , R o t t e r d a m , I(2), 1-75.
P o s t m a , H., 1984. I n t r o d u c t i o n to t h e s y m p o s i u m o n o r g a n i c m a t t e r in t h e W a d d e n S e a . - Publ. Set. N e t h . Inst. S e a Res. I0, 15-22.
Reise, K., 1980. H u n d e r t J a h r e BiozSnose. - N a t u r w . Rdsch., Stuttg. 33, 3 2 8 - 3 3 5 . Reise, K., 1985. T i d a l flat ecology. S p r i n g e r , Berlin, 191 pp.
Reise, K , 1990. G r u n d g e d a n k e n z u r 5 k o l o g i s c h e n W a t t f o r s c h u n g . U m w e l t b u n d e s a m t , Berlin, 138-146. (Texte 7/90).
Reise, K., 1990. Karl M S b i u s : D r e d g i n g t h e first c o m m u n i t y c o n c e p t f r o m t h e b o t t o m of t h e sea. - D t .
h y d r o g r . Z. Erg. H. (Reihe B) 22, 149-152.
Reise, K., 1991. M o s a i c cycles in t h e m a r i n e b e n t h o s . In: T h e m o s a i c - c y c l e c o n c e p t of e c o s y s t e m s . Ed. b y H. R e m m e r t , S p r i n g e r , Berlin, 61-82.
Reise, K., H e r r e , E. & Sturm, M., 1989. Historical c h a n g e s in t h e b e n t h o s of t h e W a d d e n S e a a r o u n d t h e i s l a n d of Sylt in t h e N o r t h Sea. - H e l g o l / i n d e r M e e r e s u n t e r s . 43, 4 1 7 - 4 3 3 .
R o s e n b e r g , R., 1985. E u t r o p h i c a t i o n - t h e f u t u r e m a r i n e c o a s t a l n u i s a n c e ? - Mar. Poltut. Bull. I6,
2 2 7 - 2 3 1 .
Smidt, E. L., 1951. A n i m a l p r o d u c t i o n in t h e D a n i s h W a d d e n Sea. - M e d d r D a n m . F i s k . - o g H a v u n d e r s . I I (6), 1-151.
Streif, H., 1989. Barrier i s l a n d s , tidal fiats, a n d c o a s t a l m a r s h e s r e s u l t i n g from a r e l a t i v e rise of s e a l e v e l in E a s t Frisia o n t h e G e r m a n N o r t h S e a coast. K l u w e r , D o r d r e c h t , 2 1 3 - 2 2 3 .
T h a m d r u p , H. M., 1935. Beitr~ige z u r O k o l o g i e d e r W a t t e n f a u n a . - M e d d r D a n m . Fisk.- o g H a v u n d e r s . 10 (2), 1-125.
V a u k , G., Pr'Liter, J. & H a r t w i g , E., i989. L o n g - t e r m p o p u l a t i o n d y n a m i c s of b r e e d i n g b i r d s p e c i e s in t h e G e r m a n W a d d e n S e a area. - H e l g o l / i n d e r M e e r e s u n t e r s . 43, 3 5 7 - 3 6 5 .
W a r m i n g , E., 1904. V a d e r n e s , s a n d e n e s og m a r s k e n s n a t u r h i s t o r i k . - K. d a n s k e V i d e n s k . Selsk. Skr. 7 (2), 1-56.
Wilde, P. A: W. J. d e & B e u k e m a , J. J., 1984. T h e role of t h e z o o b e n t h o s in t h e c o n s u m p t i o n of o r g a n i c m a t t e r in t h e D u t c h W a d d e n Sea. - Publ. Set. N e t h . Inst. S e a Res. 10, 1 4 5 - 1 5 8 . Witte, J. Y. & Zijlstra, J. J., 1984. T h e m e i o f a u n a of a tidal flat in t h e w e s t e r n p a r t of t h e W a d d e n S e a
a n d its role in t h e b e n t h i c e c o s y s t e m . - Mar. Ecol. Prog. Set. 14, 129-138.
W o h l e n b e r g , E., 1931. Die G r t i n e Insel in d e r E i d e r m / . i n d u n g . - Arch. dt. S e e w a r t e 50 (2), 1-34. W o h l e n b e r g , E., 1937. Die W a t t e n m e e r - L e b e n s g e m e i n s c h a f t e n i m K S n i g s h a f e n y o n Sylt. - H e l g o -
1/inder wiss. M e e r e s u n t e r s . I, 1-92.
W o h l e n b e r g , E., 1953. Sinkstoff, S e d i m e n t u n d A n w a c h s a m H i n d e n b u r g d a m m . - Kiiste 2, 3 3 - 9 3 . Wolff, W. J., 1992a. E c o l o g i c a l d e v e l o p m e n t s in t h e W a d d e n S e a u n t i l 1990. - Publ. Ser. N e t h . Inst.
S e a Res. 20, 2 3 - 3 2 .
Wolff, W. J., 1992b. T h e e n d of tradition: 1000 y e a r s of e m b a n k m e n t a n d r e c l a m a t i o n of w e t l a n d s in t h e N e t h e r l a n d s . - A m b i o 21, 2 8 7 - 2 9 1 .
Wolff, W. J. & Zijlstra, J. J., 1980. M a n a g e m e n t of t h e W a d d e n Sea. - H e l g o l ~ n d e r M e e r e s u n t e r s . 33,